羊肚菌白霉病病原鑒定及生物學(xué)特性研究
2023-01-17 07:01:56蘇文英劉曉梅紀(jì)偉梁長(zhǎng)東趙書(shū)光任立凱
浙江農(nóng)業(yè)科學(xué) 2023年1期
關(guān)鍵詞:生長(zhǎng)
蘇文英,劉曉梅,紀(jì)偉,梁長(zhǎng)東,趙書(shū)光,任立凱*
(1.連云港市農(nóng)業(yè)科學(xué)院,江蘇 連云港 222006;2.灌南縣農(nóng)業(yè)技術(shù)推廣中心,江蘇 連云港 222006)
羊肚菌(Morchella)是一種珍稀食藥用菌,在菌物分類(lèi)學(xué)上隸屬于子囊菌亞門(mén)(Ascomycota)、盤(pán)菌綱(Discomycetes)、盤(pán)菌目(Pezizales)、羊肚菌科(Morchellaceae)、羊肚菌屬(Morchella)[1]。羊肚菌風(fēng)味獨(dú)特,營(yíng)養(yǎng)價(jià)值極高,富含多種蛋白質(zhì)及人體所需氨基酸,具有抑制腫瘤、調(diào)節(jié)機(jī)體免疫功能,能夠抗疲勞、保肝、保腎、調(diào)血脂及保護(hù)胃腸功能等作用,具有重要的經(jīng)濟(jì)價(jià)值[2-3]。由于羊肚菌產(chǎn)業(yè)成本低、見(jiàn)效快、效益高的優(yōu)勢(shì),其栽培區(qū)域由最初的川渝地區(qū)迅速擴(kuò)張,現(xiàn)如今全國(guó)各地除海南以外的各省份均有種植[4-5]。羊肚菌屬于土腐生類(lèi)型,菌絲體需要在土壤中的有機(jī)物、無(wú)機(jī)物、水分及各種微生物的作用下才能出菇。因此,羊肚菌大田栽培占全行業(yè)栽培的90%以上,整個(gè)栽培過(guò)程羊肚菌菌絲體及子實(shí)體都暴露在外界環(huán)境中,與環(huán)境中的各種生物體接觸,容易招致各種病蟲(chóng)害侵襲,給生產(chǎn)帶來(lái)?yè)p失。此外,在栽培管理后期,高溫高濕的環(huán)境也會(huì)加劇病害的發(fā)生和蔓延[6]。
近幾年,羊肚菌病害在全國(guó)范圍內(nèi)頻發(fā),病害種類(lèi)包括細(xì)菌性病害和真菌性病害,細(xì)菌性病害主要為軟腐病及紅體病;真菌性病害主要為白霉病。羊肚菌白霉病是其子實(shí)體時(shí)期最主要的病害,發(fā)病時(shí)子實(shí)體受侵染部位被白色絨毛狀菌絲覆蓋,嚴(yán)重時(shí)子實(shí)體受侵染部位會(huì)枯萎,病害發(fā)生率達(dá)60%~80%,使羊肚菌的商品性狀和經(jīng)濟(jì)價(jià)值大打折扣,給種植戶(hù)帶來(lái)巨大的經(jīng)濟(jì)損失。此外,羊肚菌白霉病菌適應(yīng)性極強(qiáng),且能快速傳播,一旦發(fā)病極難控制。因此,掌握羊肚菌白霉病病原菌的物種歸屬及生物學(xué)特性,對(duì)于該病害的預(yù)防和控制及進(jìn)一步提升羊肚菌種植的經(jīng)濟(jì)效益具有一定的實(shí)踐指導(dǎo)意義。
本研究通過(guò)對(duì)羊肚菌白霉病病原菌的分離鑒定及生物學(xué)特性進(jìn)行研究,以期為該病害的早期診斷和科學(xué)防治提供理論依據(jù)。
1 材料與方法
1.1 供試菌株
2020年3月在江蘇省連云港市農(nóng)業(yè)科學(xué)院試驗(yàn)基地,采集發(fā)病子實(shí)體并帶回實(shí)驗(yàn)室,觀察記錄病害癥狀。
1.2 供試培養(yǎng)基
MYG培養(yǎng)基(稱(chēng)取10 g麥芽糖、5 g葡萄糖、5 g酵母浸粉、15 g瓊脂,加入蒸餾水定容至1 L);基礎(chǔ)培養(yǎng)基(稱(chēng)取20 g葡萄糖、10 g蛋白胨、1 g K2HPO4、0.5 g MgSO4、15 g瓊脂,加入蒸餾水定容至1 L);馬鈴薯葡萄糖瓊脂(PDA)培養(yǎng)基(稱(chēng)取200 g馬鈴薯、20 g葡萄糖、15 g瓊脂,加入蒸餾水定容至1 L);薩氏(SDAY)培養(yǎng)基(稱(chēng)取10 g酵母浸粉、40 g葡萄糖、10 g蛋白胨、15 g瓊脂,加入蒸餾水定容至1 L);察氏(Czapek)培養(yǎng)基(稱(chēng)取2 g NaNO3、1 g K2HPO4、0.5 g KCl、0.5 g MgSO4、0.01 g FeSO4、30 g蔗糖、15 g瓊脂,加入蒸餾水定容至1 L),所有培養(yǎng)基均在121 ℃下高壓滅菌20 min。
1.3 病原菌的分離與鑒定
采用組織分離法進(jìn)行病原菌的分離和純化,在超凈工作臺(tái)中,用75%乙醇棉擦拭病害子實(shí)體表面,用接種刀切取病健交界處1~2 mm的組織塊置于MYG平板內(nèi)于25 ℃下恒溫培養(yǎng)。待組織塊周?chē)L(zhǎng)出菌絲,切取尖端菌絲轉(zhuǎn)至新的MYG培養(yǎng)基純化培養(yǎng)。共獲得1個(gè)菌株標(biāo)記為JS-1,菌株在MYG培養(yǎng)基上培養(yǎng)10 d后置于4 ℃保藏備用。
利用基因組DNA提取試劑盒(北京康為世紀(jì)生物科技有限公司,北京)提取病原真菌DNA,選擇通用引物ITS1、ITS4對(duì)提取的DNA進(jìn)行擴(kuò)增,擴(kuò)增產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳檢測(cè),檢測(cè)正確后送至生工生物工程(上海)股份有限公司進(jìn)行測(cè)序,測(cè)序結(jié)果進(jìn)行BLAST同源性比對(duì)。通過(guò)比對(duì)測(cè)序結(jié)果,選取相似性較高的菌株相關(guān)種屬序列,利用MEGA 7.0構(gòu)建系統(tǒng)發(fā)育樹(shù)。
將菌株JS-1接種于MYG平板上,25 ℃恒溫暗培養(yǎng)10 d,觀察記錄菌落特征及顯微形態(tài)。
根據(jù)柯赫氏法則進(jìn)行回接致病性檢測(cè)。
1.4 病原菌生物學(xué)特性測(cè)定
1.4.1 不同培養(yǎng)基對(duì)病原菌菌絲生長(zhǎng)的影響
用8 mm打孔器在致病菌菌落邊緣打孔,將菌塊分別接種于PDA培養(yǎng)基、SDAY培養(yǎng)基、MYG培養(yǎng)基、Czapek培養(yǎng)基上,每個(gè)處理5個(gè)重復(fù),于25 ℃恒溫暗培養(yǎng)。培養(yǎng)期間觀察記錄菌絲均勻度及密度等長(zhǎng)勢(shì)情況,并計(jì)算菌絲生長(zhǎng)速率。
1.4.2 碳源試驗(yàn)
分別以等量麥芽糖、淀粉、蔗糖、乳糖替換基礎(chǔ)培養(yǎng)基中的葡萄糖,121 ℃滅菌20 min后倒入培養(yǎng)皿中,待培養(yǎng)基凝固后,用直徑8 mm的打孔器取菌塊接種于平板中央,每個(gè)處理5個(gè)重復(fù),25 ℃恒溫條件下黑暗培養(yǎng),觀察并記錄菌絲長(zhǎng)勢(shì)。
1.4.3 氮源試驗(yàn)
分別以等量硫酸銨、酵母浸粉、氯化銨、尿素替換基礎(chǔ)培養(yǎng)基中的蛋白胨,121 ℃滅菌20 min后倒入培養(yǎng)皿中,待培養(yǎng)基凝固后,用直徑8 mm的打孔器取菌塊接種于平板中央,每個(gè)處理5個(gè)重復(fù),25 ℃恒溫條件下黑暗培養(yǎng),觀察并記錄菌絲長(zhǎng)勢(shì)。
1.4.4 溫度試驗(yàn)
將接種后的平板培養(yǎng)皿分別置于15、20、25、30、35、40 ℃條件下培養(yǎng),每個(gè)處理5個(gè)重復(fù)(培養(yǎng)基為基礎(chǔ)培養(yǎng)基),觀察并記錄菌絲長(zhǎng)勢(shì)。
1.4.5 pH試驗(yàn)
用鹽酸和氫氧化鈉溶液調(diào)各培養(yǎng)基初始pH為5、6、7、8、9,觀察并記錄菌絲長(zhǎng)勢(shì)。
1.5 數(shù)據(jù)分析
用Excel和SPSS 17.0對(duì)數(shù)據(jù)進(jìn)行整理和分析。
2 結(jié)果與分析
2.1 病害癥狀
2020年3月對(duì)連云港市農(nóng)業(yè)科學(xué)院試驗(yàn)基地羊肚菌大棚調(diào)查發(fā)現(xiàn),發(fā)病初期在子實(shí)體上出現(xiàn)白色小斑點(diǎn),斑點(diǎn)逐漸擴(kuò)大并穿孔,幼小的子實(shí)體被病原菌的白色菌絲覆蓋,最終停止生長(zhǎng)(圖1中A和B)。
2.2 病原菌的分離純化及形態(tài)學(xué)鑒定
從發(fā)病羊肚菌樣品中,分離獲得致病菌JS-1。將致病菌菌餅覆蓋在羊肚菌菌帽部分,再置于培養(yǎng)皿中,3次重復(fù),保濕培養(yǎng)若干天,直至有病斑形成,將病斑部位再進(jìn)行分離培養(yǎng),通過(guò)培養(yǎng)觀察菌落形態(tài)、顯微形態(tài)及ITS序列確定分離到的病原菌與發(fā)病菌株的致病菌為同一株病原菌。
在MYG培養(yǎng)基和25 ℃黑暗條件下,菌株JS-1的菌落初期為白色,輪狀生長(zhǎng),邊緣規(guī)則,菌絲絨毛狀,較發(fā)達(dá),后期菌絲生長(zhǎng)部位培養(yǎng)基萎縮(圖1中C)。分生孢子梗側(cè)生,分生孢子呈鏈?zhǔn)剑环稚咦訔l形或梭形,有間隔(圖1中D)。

A—自然感病子實(shí)體;B—感病部位;C—病原菌在MYG平板25 ℃培養(yǎng);D—病原菌顯微形態(tài)。圖1 病原菌形態(tài)特征
2.3 病原菌的分子生物學(xué)鑒定
經(jīng)PCR擴(kuò)增和序列測(cè)定,獲得供試菌株ITS序列片段,BLAST在線比對(duì)結(jié)果表明,JS-1與長(zhǎng)毛擬青霉Paecilomycespenicillatus(登錄號(hào):EU146306.1)相似度為99.81%;與長(zhǎng)孢卵單隔孢霉Diplo?sporalongispora(登錄號(hào):KX427537.1)相似度為99.44%。將致病菌的ITS序列與相近種菌株構(gòu)建系統(tǒng)發(fā)育樹(shù)(圖2),結(jié)果顯示,所得病原菌JS-1與長(zhǎng)毛擬青霉菌株及長(zhǎng)孢卵單隔孢霉聚于同一分支,說(shuō)明其親緣關(guān)系較近。
2.4 病原菌生物學(xué)特性
致病菌菌絲在不同培養(yǎng)基下的生長(zhǎng)情況見(jiàn)表1。在Czapek培養(yǎng)基上,致病菌菌絲生長(zhǎng)速度最快,但中心菌絲濃密,邊緣菌絲稀疏;在MYG培養(yǎng)基上,菌絲生長(zhǎng)速度較快,菌落邊緣整齊,菌絲濃密;其次為PDA培養(yǎng)基;SDAY培養(yǎng)基菌絲生長(zhǎng)最為緩慢。在菌絲生長(zhǎng)后期,可以觀察到培養(yǎng)基萎縮甚至穿孔。
致病菌菌絲在不同pH試驗(yàn)結(jié)果見(jiàn)表2。羊肚菌致病菌菌絲在pH 5~9的條件下均可生長(zhǎng),pH 6~8時(shí)菌絲生長(zhǎng)速度較快,且致病菌菌絲在不同pH下差異并不顯著。
致病菌菌絲在22~34 ℃溫度范圍下的生長(zhǎng)情況見(jiàn)表3,在25 ℃條件下培養(yǎng)的實(shí)驗(yàn)菌株菌絲生長(zhǎng)速度最快,其次為22 ℃,溫度高于31 ℃時(shí),菌絲不生長(zhǎng)。因此,JS-1菌絲較適宜的生長(zhǎng)溫度為25 ℃。
碳源試驗(yàn)結(jié)果表明,致病菌株在不同碳源培養(yǎng)基上均能生長(zhǎng)(表4),但菌落形態(tài)及菌絲生長(zhǎng)速率不同。在以乳糖為碳源的培養(yǎng)基上菌絲生長(zhǎng)速率最快,為0.225 cm·d-1;其次為葡萄糖及淀粉,分別為0.205及0.201 cm·d-1。各培養(yǎng)基的菌絲密度差異較為顯著,其中淀粉為碳源時(shí),菌絲生長(zhǎng)最為濃密;以葡萄糖為碳源時(shí),菌絲較為稀疏。因此,致病菌JS-1菌絲生長(zhǎng)較適宜碳源為乳糖。

圖2 ITS序列系統(tǒng)發(fā)育樹(shù)

表1 培養(yǎng)基對(duì)菌絲生長(zhǎng)的影響
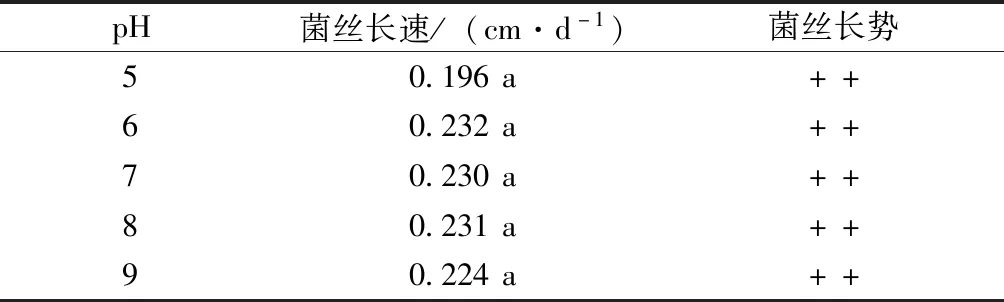
表2 pH對(duì)菌絲生長(zhǎng)的影響

表3 溫度對(duì)菌絲生長(zhǎng)的影響
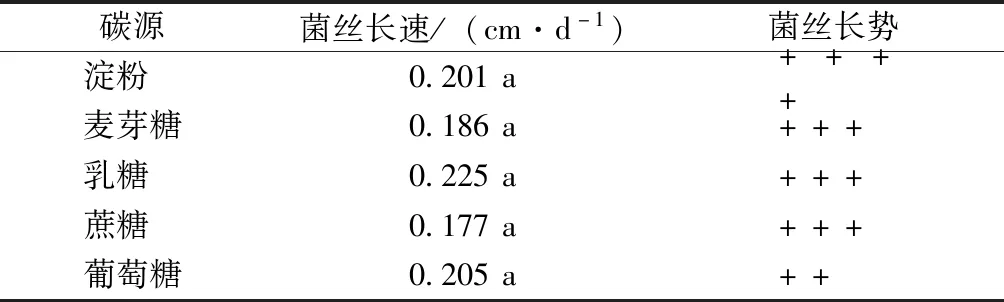
表4 碳源對(duì)菌絲生長(zhǎng)的影響
氮源試驗(yàn)結(jié)果表明,菌株在不同氮源培養(yǎng)基上均能生長(zhǎng),但氮源不同,菌絲體生長(zhǎng)速度及菌落形態(tài)差異較大(表5)。在以酵母浸粉為氮源的培養(yǎng)基上,菌絲體生長(zhǎng)速度最快,為0.312 cm·d-1,菌絲生長(zhǎng)濃密。其次為以氯化銨為氮源的培養(yǎng)基,菌絲生長(zhǎng)速度為0.222 cm·d-1,但菌絲生長(zhǎng)稀疏。而在硫酸銨、磷酸氫二鉀為氮源的培養(yǎng)基上,菌絲體生長(zhǎng)速度很慢,而且菌絲長(zhǎng)勢(shì)差。因此,致病菌JS-1菌絲體較適宜的氮源為酵母浸粉。
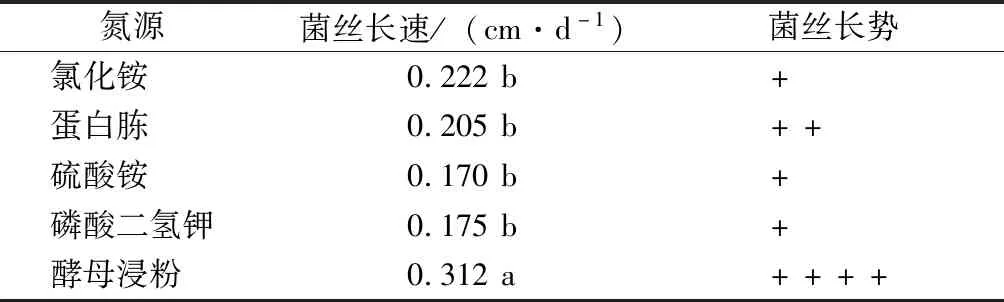
表5 氮源對(duì)菌絲生長(zhǎng)的影響
3 討論
近年來(lái),羊肚菌因其低投入高回報(bào)的特點(diǎn)在全國(guó)范圍內(nèi)種植面積大幅增加,對(duì)其致病菌進(jìn)行研究,對(duì)保證其品質(zhì)和產(chǎn)量有重要意義。目前,已有研究表明,多種病原菌可侵染羊肚菌子實(shí)體導(dǎo)致真菌病害發(fā)生。劉偉等[6]報(bào)道羊肚菌的真菌性病害主要有霉菌性枯萎病、蛛網(wǎng)病、鐮刀菌病。劉天海等[7]對(duì)四川等地暴發(fā)的柄腐病害進(jìn)行調(diào)查鑒定,結(jié)合菌落形態(tài)和顯微特征鑒定柄腐病的病原菌為Fusariumnematophilum,該病原菌菌落正面白色,背面米白色,不透明,隆起,菌絲致密,邊緣整齊,菌落呈同心環(huán)紋,菌絲有隔,有分枝,無(wú)水溶性色素,分生孢子呈鐮刀型,整體較粗壯,中部略寬于兩端,尖端短而小,大多3隔,極少數(shù)1隔、2隔或4隔,呈3隔孢子的隔間距7~12 μm。余苗等[8]對(duì)羊肚菌白腐病病菇進(jìn)行分離鑒定,采用形態(tài)學(xué)研究,結(jié)合ITS序列分析技術(shù)鑒定病原菌為曲霉(Aspergillussp.),該病原真菌菌絲體顏色為白色,菌落邊緣整齊呈圓形,稍凸起,產(chǎn)生分生孢子后呈黃綠色,分生孢子球形、近球形。黃慧等[9]對(duì)貴州六盤(pán)水等地暴發(fā)的羊肚菌菌蓋干腐病進(jìn)行病原菌的分離鑒定,結(jié)合形態(tài)學(xué)和分子生物學(xué)鑒定,發(fā)現(xiàn)病原菌為長(zhǎng)孢卵單隔孢霉(D.longispora),該致病菌菌落邊緣整齊、表面干燥,呈白色絨毛狀,有輪狀花紋,顯微形態(tài)下分生孢子梗透明,呈細(xì)長(zhǎng)棍棒狀,分生孢子鏈生,呈棍棒狀或長(zhǎng)檸檬形狀,有隔。
本研究分離得到的致病菌菌落初期為白色,輪狀生長(zhǎng),邊緣規(guī)則,菌絲絨毛狀,較發(fā)達(dá),后期菌絲生長(zhǎng)部位培養(yǎng)基萎縮,顯微形態(tài)下分生孢子梗側(cè)生,分生孢子鏈?zhǔn)剑环稚咦訔l形或梭形,有間隔。測(cè)序獲得的ITS序列經(jīng)NCBI比對(duì)后發(fā)現(xiàn)與長(zhǎng)毛擬青霉相似度為99.81%;與長(zhǎng)孢卵單隔孢霉相似度為99.44%,進(jìn)一步構(gòu)建系統(tǒng)發(fā)育樹(shù)表明JS-1的ITS序列與長(zhǎng)毛擬青霉和長(zhǎng)孢卵單隔孢霉聚為一支,這兩者均為羊肚菌真菌病害的主要病原菌[10-11]。前人研究[12-13]結(jié)果表明,這2種真菌的分生孢子都呈鏈?zhǔn)剑L(zhǎng)毛擬青霉的分生孢子呈橢圓形或梨形,無(wú)隔;而長(zhǎng)孢卵單隔孢霉的分生孢子為棍棒狀或長(zhǎng)檸檬形,有隔。通過(guò)比對(duì)其形態(tài)學(xué)特征,本研究分離得到的致病菌在形態(tài)上更接近于長(zhǎng)孢卵單隔孢霉。因此,通過(guò)結(jié)合發(fā)病特征、形態(tài)學(xué)觀察及構(gòu)建分子系統(tǒng)發(fā)育樹(shù)表明該病害是由長(zhǎng)孢卵單隔孢霉(D.longispora)侵染引起的。
此外,生物學(xué)特性研究結(jié)果表明,羊肚菌白霉病病菌最適生長(zhǎng)溫度為25 ℃,30 ℃以上不生長(zhǎng),pH在6~9時(shí)生長(zhǎng)較快,說(shuō)明該菌不耐高溫,對(duì)pH適應(yīng)范圍較廣,與黃慧等[9]的研究結(jié)果一致;最適碳、氮源為乳糖及酵母浸粉。本研究通過(guò)對(duì)羊肚菌白霉病病原菌的鑒定,尤其是對(duì)其進(jìn)行系統(tǒng)的生物學(xué)特性研究,為進(jìn)一步研究該病害的發(fā)病規(guī)律及田間防治奠定理論基礎(chǔ)。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
