植物觀賞器官中類胡蘿卜素代謝調控的研究進展
2023-01-05 07:48:20田清尹岳遠征申慧敏潘多楊秀蓮王良桂
生物技術通報 2022年12期
關鍵詞:植物
田清尹 岳遠征 申慧敏 潘多 楊秀蓮 王良桂
(1.南京林業大學風景園林學院 南京 210037;2.南京林業大學風景園林江蘇省重點實驗室,南京 210037)
類胡蘿卜素(carotenoids)是一類重要的脂溶性色素,呈鏈狀或環狀,由碳氫化合物及其含氧衍生物組成,廣泛存在于各種動植物和微生物之中。作為次生代謝物,類胡蘿卜素不僅在植物的生長發育過程中參與或調控許多重要的生理進程,而且也是人體所需的重要營養物質[1]。在植物中,類胡蘿卜素賦予植物的花、葉及果實鮮艷色彩,既提高了觀賞價值,又可以吸引昆蟲傳播花粉促進自身的繁衍。同時,類胡蘿卜素在植物生長發育、光合作用以及植物激素和揮發性物質的產生中也有著重要作用[2]。
植物觀賞器官包括花、果、葉等,其色彩和香味是植物重要的觀賞性狀。積累在植物觀賞器官中不同類型的類胡蘿卜素促進了不同色彩的形成,例如番茄紅素、β-胡蘿卜素和葉黃素的積累分別呈現紅色、橙黃色和黃色[3]。同時類胡蘿卜素裂解后產生多種類型的脫輔基類胡蘿卜素,也可以為植物提供不同的色澤和香味,如在藏紅花(Crocus sativus)中,藏紅花素和藏紅花醛是其柱頭顏色和風味的主要來源[4]。鑒于類胡蘿卜素在調控植物重要觀賞器官著色及香氣物質產生等方面的重要意義,本文對類胡蘿卜素代謝調控植物花、果、葉的色香變化進行了總結,為人工干預調節植物中類胡蘿卜素的種類或含量,進而提高植物的觀賞價值提供參考。
1 類胡蘿卜素代謝途徑及其調控機制
1.1 類胡蘿卜素合成和降解途徑
1.1.1 類胡蘿卜素的生物合成 目前,類胡蘿卜素代謝途徑已被廣泛研究(圖1)[3]。在植物中,類胡蘿卜素的前體物質是經由甲基赤蘚糖醇途徑(methyl erythritol phosphate,MEP)產生的兩種異戊二烯異構體:異戊烯基焦磷酸(isopentenyl diphosphate,IPP)和二甲基丙烯基焦磷酸(dimethylallyl diphosphate,DMAPP)[5]。三分子IPP 和一分子DMAPP 在牻牛兒基牻牛兒基焦磷酸合酶(geranylgeranyl pyrophosphate synthase,GGPPS)的催化下合成牻牛兒基牻牛兒基焦磷酸(geranylgeranyl diphosphate,GGPP)。值得注意的是,IPP 和DMAPP 是類胡蘿卜素與單萜、倍半萜等揮發性物質合成的共同底物,因此,控制IPP 和DMAPP 的流向是植物色彩和香味改良的重要手段[6]。
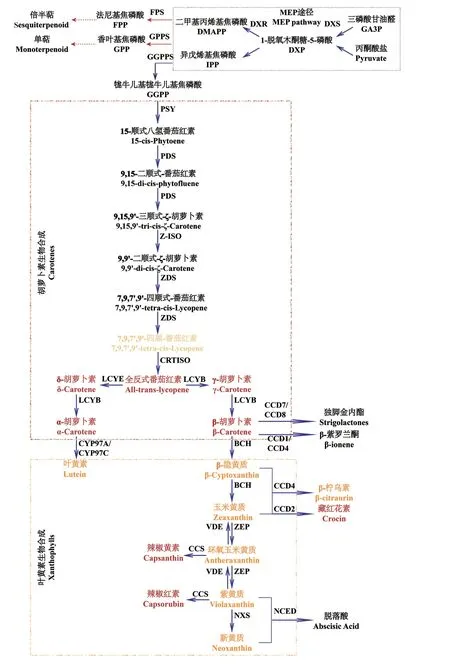
圖1 類胡蘿卜素合成、降解途徑Fig.1 Pathways of carotenoid synthesis and degradation
類胡蘿卜素的生物合成始于八氫番茄紅素合酶(phytoene synthase,PSY)催化兩分子GGPP 縮合,形成無色的15-順式-八氫番茄紅素。然后依次經過八氫番茄紅素脫氫酶(phytoene desaturase,PDS)、ζ-胡蘿卜素脫氫酶(ζ-carotene desaturase,ZDS)催化脫氫,ζ-胡蘿卜素異構酶(ζ-carotene isomerase,Z-ISO)、類胡蘿卜素異構酶(carotene isomerase,CRTISO)催化異構形成紅色的番茄紅素。番茄紅素的環化是類胡蘿卜素生物合成途徑的重要分支點[7],番茄紅素在番茄紅素ε-環化酶(lycopene ε-cyclase,LCYE)和番茄紅素β-環化酶(lycopene β-cyclase,LCYB)共同催化下,生成α-胡蘿卜素,而僅在LCYB 的催化下,則產生β-胡蘿卜素。隨后,含血紅素的細胞色素P450 型羥化酶(cytochrome P450 carotene hydroxylase,CYP)催化α-胡蘿卜素形成葉黃素;非血紅素β-環羥化酶(β-carotene hydrolase,BCH)催化β-胡蘿卜素,依次形成β-隱黃質和玉米黃質。玉米黃質通過玉米黃質環氧酶(zeaxanthin epoxidase,ZEP)環氧化產生環氧玉米黃質和紫黃質,紫黃質則可經紫黃質脫環氧酶(violaxanthin deepoxidase,VDE)重新生成玉米黃質,或是在新黃質合成酶(neoxanthin synthase,NXS)的作用下,形成新黃質。此外,在辣椒(Capsicum annuum)和卷丹(Lilium lancifolium)中,環氧玉米黃質和紫黃質在辣椒紅素合酶(capsanthin-capsorubin synthase,CCS)的催化下轉化為辣椒紅素和辣椒黃素,呈現獨特的紅色和橙色[8-9]。
1.1.2 類胡蘿卜素降解途徑 類胡蘿卜素降解主要包括非特異性機制氧化(如光化學氧化)、非特異性酶分解(脂氧合酶和過氧化物酶)以及類胡蘿卜素裂解雙加氧酶(carotenoid cleavage dioxygenases,CCDs)裂解[10]。在植物中,由CCDs 催化的酶促反應是類胡蘿卜素降解的主要方式[11]。CCDs 家族根據功能不同被分為兩個亞家族:(1)9-順式-環氧類胡蘿卜素雙加氧酶(9-cis-epoxycarotenoid dioxygenase,NCED),特異參與脫落酸前體的產生;(2)類胡蘿卜素裂解雙加氧酶(CCD),能夠催化降解不同的類胡蘿卜素生成多種類型的脫輔基類胡蘿卜素,如β-檸檬酸[12]和β-紫羅蘭酮[13]等,在植物的色素沉積、揮發物質合成等方面有著重要作用。
1.2 類胡蘿卜素代謝分子調控機制
1.2.1 轉錄因子調控 轉錄因子(TFs)是一種能夠識別和結合基因5′端上游特定序列并調控基因表達的蛋白質,在類胡蘿卜素結構基因的轉錄激活或抑制中發揮關鍵作用。在植物的花器官中,類胡蘿卜素代謝途徑的結構基因受到一些轉錄因子間接或直接的調控(圖2-A)。RCP1(reduced carotenoid pigmentation 1)是第一個被報道的花特異性轉錄因子,屬于R2R3-MYB 基因家族,在猴面花(Mimulus lewisii)的花發育過程中正向調控整個類胡蘿卜素合成途徑;之后發現的RCP2也具有類似功能,但是RCP1與RCP2是否直接調控類胡蘿卜素生物合成基因的表達尚不清楚[14-15]。而同基因家族的WHITE PETAL1(WP1)則被證明其作為轉錄激活因子直接調節蒺藜苜蓿(Medicago truncatula)花瓣中MtLYCE和MtLYCB等基因的表達,進而調節花瓣中類胡蘿卜素色素沉著[16];在桂花(Osmanthus fragrans)中,OfWRKY3、OfERF61可以直接與OfCCD4啟動子相互作用,促進類胡蘿卜素裂解形成β-紫羅蘭酮[17-18]。
在植物果實中,已報道參與類胡蘿卜素代謝的轉錄因子有MADS-box、SPL、NAC、AP2/ERF、bHLH等,這些轉錄因子多與果實發育、乙烯生物合成和光形態建成有關(圖2-B)。MADS-box編碼的轉錄因子參與調控果實成熟,在番茄(Solanum lycopersicum)中,SlRIN通過促進番茄紅素的合成并抑制其環化,參與番茄紅素的積累,SlFUL1/2 則調節整個類胡蘿卜素代謝途徑[19-20];SPL轉錄因子與果實生長調控相關,在香蕉(Musa nana)中,MaSPL16直接結合MaLCYB1.1和MaLCYB1.2的啟動子,正向調節類胡蘿卜素合成[21];SlNAC1被報道參與乙烯的生物合成,同時SlNAC1可以與SlPSY1相互作用,抑制番茄果實中總類胡蘿卜素和番茄紅素的積累[22];AP2/ERF是乙烯響應因子,在柑橘(Citrus sinensis)中,由乙烯誘導的CsERF061作為正向調節因子,直接激活CsLCYb2和其他關鍵類胡蘿卜素途徑基因的啟動子[23];BBX-box在光信號傳導中起重要作用,SlBBX20直接與SlPSY1的啟動子結合并激活其表達,促進番茄葉片和果實中類胡蘿卜素的生物合成[24];光敏色素互作因子(PIF)屬于bHLH 家族,與SlPSY啟動子的G-box 光響應元件特異結合,從而抑制SlPSY的表達和番茄中類胡蘿卜素的積累。此外,在木瓜(Carica papaya)中,CpBHLH1/2也參與了光介導的類胡蘿卜素合成調控[25-26]。
在植物的葉片中,僅少數調控類胡蘿卜素代謝的轉錄因子被鑒定出來(圖2-C)。例如PIF1和HY5,在擬南芥(Arabidopsis thaliana)葉片中直接調控AtPSY的表達以響應光/暗周期和溫度的變化[27];AtRAP2.2在體外被證明能結合AtPSY和AtPDS啟動子,然而在擬南芥葉片中過表達AtRAP2.2并沒有導致AtPSY或AtPDS轉錄水平升高,也沒有改變類胡蘿卜素的含量,因此還需要進一步研究相關的調控機制[28]。
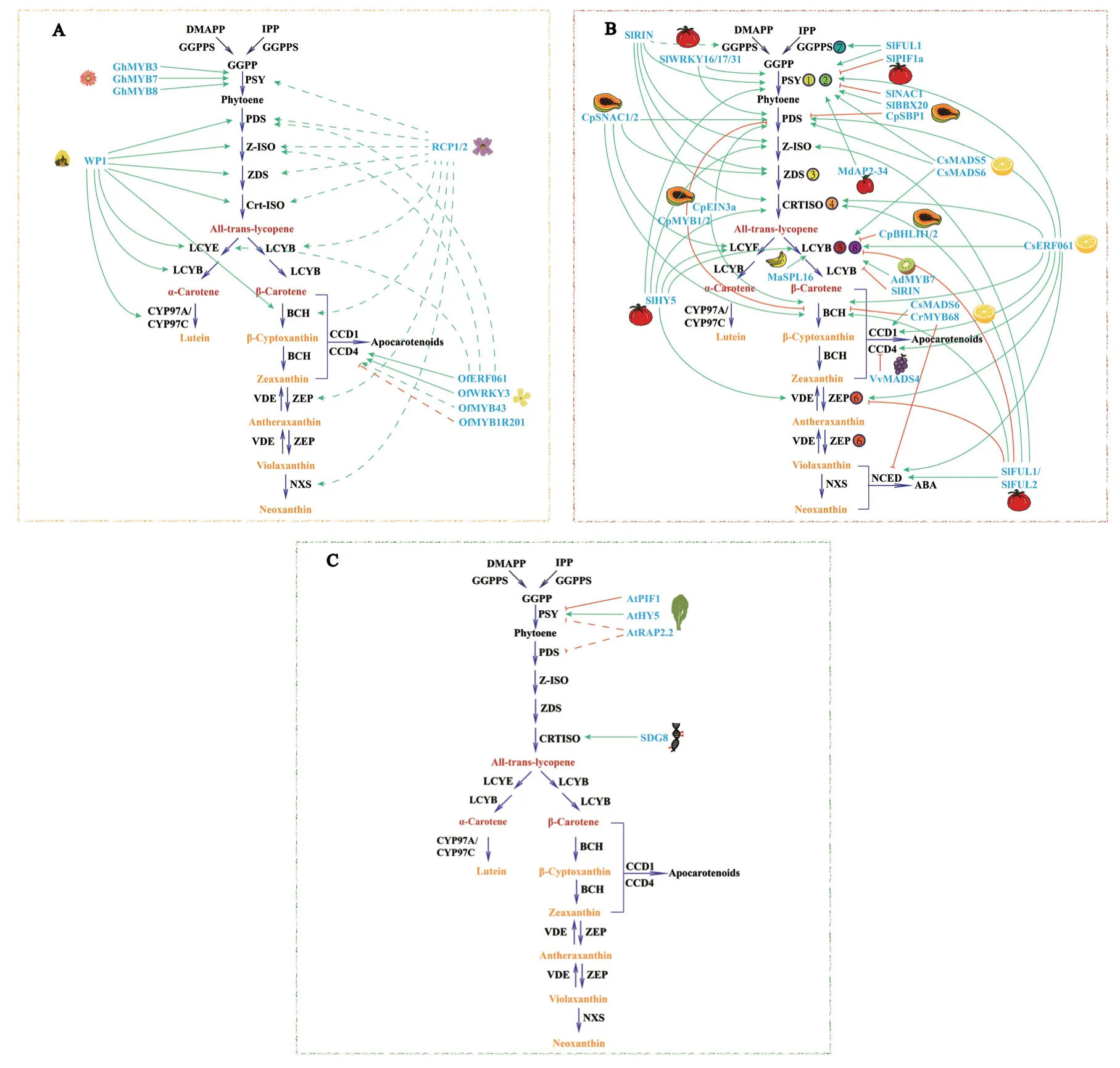
圖2 花、果、葉中類胡蘿卜素代謝調控圖Fig.2 Regulation of carotenoid metabolism in flowers,fruits and leaves
1.2.2 表觀遺傳調控 類胡蘿卜素代謝的表觀遺傳調控主要有組蛋白修飾、DNA 甲基化和miRNA 介導的轉錄后基因沉默等。首先,組蛋白修飾包括泛素化、乙酰化、磷酸化和甲基化等,是一種可逆的表觀遺傳修飾。番茄E3 泛素連接酶(PPSR1)通過泛素化介導PSY 蛋白降解,研究表明PPSR1 功能喪失會導致果實中類胡蘿卜素含量增加[29];沉默組蛋白去乙酰化酶(SlHDT3)可以延緩番茄果實成熟并抑制類胡蘿卜素的積累[30]。其次,DNA 甲基化對相關基因轉錄的抑制作用也會影響類胡蘿卜素的合成和降解,在擬南芥的組蛋白甲基轉移酶(SDG8)缺失突變體中,關鍵基因CRTISO的染色質甲基化狀態改變、表達水平降低,導致葉黃素合成減少[31]。此外,miRNAs 作為植物體內重要的調節因子,也參與調控類胡蘿卜素代謝,例如csi-miR1857 靶向LCYB基因進而調控柑橘紅肉突變體中番茄紅素的積累[32];在番茄中,7 個與類胡蘿卜素生物合成相關的miRNA 被報道,其中miR169i-5p2 和miR1027a、miR482、miR172、miR396、miR395 分別靶向PSY、ZDS、CRTISO、LCYB、ZEP等關鍵的類胡蘿卜素合成酶基因[33]。
2 類胡蘿卜素代謝對植物花色和花香的影響
2.1 類胡蘿卜素相關基因對花色的影響
植物花瓣中類胡蘿卜素含量變化會導致花色改變,而其含量變化受到類胡蘿卜素代謝途徑中關鍵酶基因轉錄水平的影響。PSY 是類胡蘿卜素合成的重要限速酶,抑制文心蘭(Oncidium‘Gower Ramsey’)中OgPSY的轉錄會導致花瓣中類胡蘿卜素積累顯著下降,花色由黃色變為白色[34]。在歐洲油菜(Brassica napus)‘ywf’突變體中,由于BnaA08.PDS3基因突變,阻斷了類胡蘿卜素的生物合成途徑,從而導致類胡蘿卜素含量下降和黃白色花瓣表型[35];CoCRTISO1是引起金盞花(Calendula officinalis)花瓣顏色差異的關鍵基因,黃色花瓣中CoCRTISO1 催化順式番茄紅素轉化為反式番茄紅素,經環化、羥基化后最終積累黃色的葉黃素,而橙色花瓣中CoCRTISO1 酶活性較低而無法催化異構,積累紅色的5-順式番茄紅素[36];CsLCYB2a的表達水平與番紅花柱頭的類胡蘿卜素含量呈正相關,并影響其柱頭的顏色[37];InCHYB的轉錄水平不同被認為是牽牛花(Ipomoea nil)的白色和黃色花瓣之間類胡蘿卜素差異的關鍵[38];BnaC09.ZEP和BnaA09.ZEP的缺失引起歐洲油菜花瓣中類胡蘿卜素含量以及顏色的改變[39]。
類胡蘿卜素的降解多與白花的形成有關,文心蘭‘White Jade’的白花性狀是由OgCCD1的上調引起的,同時在‘Gower Ramsey’的黃色唇部組織中瞬時表達OgCCD1,出現白色斑點[40];落葉黃花杜鵑(Rhododendron japonicum)和常綠白花杜鵑的雜交種在花開放時顏色由黃變白,主要是由親本常綠白花杜鵑中CCD4基因高表達介導的[41]。此外,CCD4功能的破壞往往會導致花瓣顏色的差異,如大豆(Glycine max)‘gmicc’突變體的黃色花朵是由于GmCCD4 蛋白功能喪失導致的[42];菊花(Chrysanthemum morifolium)中CmCCD4a的缺失使得花色從白色變為黃色[43];歐洲油菜黃色花瓣的形成與轉座子插入導致CCD4功能喪失密切相關[44]。
2.2 類胡蘿卜素相關基因對花香的影響
類胡蘿卜素經CCDs 裂解產生的脫輔基類胡蘿卜素,如β-紫羅蘭酮、β-環檸檬醛和β-大馬酮等芳香物質的香氣閾值極低,因此即使這些物質的含量較低,也對花瓣整體香氣的感知有強烈影響。突厥薔薇(Rosa damascena)是一種著名的香花植物,它的香氣物質主要是由RdCCD1 分別切割新黃質和β-胡蘿卜素產生的β-大馬酮和β-紫羅蘭酮[45];在矮牽牛(Petunia hybrida)中,PhCCD1被證明參與β-紫羅蘭酮和香葉基丙酮等香味物質的形成[46]。在藏紅花柱頭發育過程中,CCD4 酶活性增強有助于揮發性β-紫羅蘭酮的產生[4];OfCCD4也是桂花花瓣中產生β-紫羅蘭酮關鍵基因,由于OfCCD4在‘銀桂’和‘丹桂’的表達差異,‘銀桂’中β-紫羅蘭酮含量明顯高于‘丹桂’[17-18]。同時,CCD4的表達差異可以同時影響花瓣的色香,百合(Lilium brownii)在開花后第二天花被片產生強烈香味,且顏色從黃色變成白色,經研究發現LbCCD4可能是控制百合開花過程中色香變化的關鍵基因[47]。
3 類胡蘿卜素代謝對植物果實色澤與風味的影響
3.1 類胡蘿卜素相關基因對果實色澤的影響
類胡蘿卜素相關基因的表達差異往往會導致果實中色素沉著的多樣性。目前利用突變體揭示類胡蘿卜素合成基因的轉錄變化對于果實色素沉著影響的研究被陸續報道,番茄γ 突變體由于基因組中插入轉座子導致SlPSY-1轉錄異常無法積累類胡蘿卜素而形成黃色果肉[48];黃色‘Pinalate’甜橙突變體的產生原因是CsZ-ISO中單核苷酸插入突變,從而使得早期上游類胡蘿卜素的大量積累[49];番茄‘tangerine’突變體中,CRTISO 酶失活導致順式番茄紅素的積累,從而產生粉橙色表型[50],此外在甜瓜(Cucumis melo)‘yofI’突變體和檸檬(Citrus limon)芽變體中也存在類似表型[51-52]。番茄紅素的含量與番茄紅素ε-或β-環化酶的轉錄水平密切相關。木瓜果肉呈現紅色的主要原因是CpLCYb2基因的插入突變導致番茄紅素環化酶的失活,引起番茄紅素積累[53],這與柑橘紅色果實的形成機制相同[7]。此外,在辣椒中,β-胡蘿卜素羥化酶(CHY2)突變導致β-胡蘿卜素及其衍生揮發物的積累,果實顏色由紅色向橙色的轉化[54];CaZEP突變使得玉米黃質環氧酶活性降低,從而使得辣椒中玉米黃質與總類胡蘿卜素含量增加,顏色由黃色變為橙色[55]。
有色的類胡蘿卜素經CCDs 降解可以生成無色的衍生物,從而對果實顏色產生影響。其中最典型的例子是桃(Prunus persica),白果肉中類胡蘿卜素含量只有黃果肉的含量的十分之一,研究發現由于黃果肉中CCD4的突變導致類胡蘿卜素積累[56],隨后利用VIGS 技術敲除了白桃中的CCD4基因,發現黃色色素沉著和類胡蘿卜素含量增加,證明了CCD4是控制桃果肉顏色變化的關鍵基因[57]。此外,類胡蘿卜素還可以降解為有色物質,如β-檸檬酸是一種C30 類胡蘿卜素,由CCD4 酶切割β-隱黃質和玉米黃質形成,使得柑橘類果實呈現紅色[12]。
3.2 類胡蘿卜素相關基因對果實香味的影響
CCD1 和CCD4 通過裂解類胡蘿卜素生成揮發性萜類化合物,進而調節果實的風味和香氣。在葡萄(Vitis vinifera)中,VvCCD1 催化β-胡蘿卜素裂解形成β-紫羅蘭酮;而VvCCD4b 催化ζ-胡蘿卜素裂解形成香葉基丙酮[58]。在番茄中,抑制LeCCD1A和LeCCD1B基因的表達會導致β-紫羅蘭酮和香葉基丙酮含量的下降[59]。同時,在甜瓜、杏(Prunus america)、無花果(Ficus carica)等果實中也存在多種揮發性脫輔基類胡蘿卜素,其種類和含量均與CCD1 酶切割類胡蘿卜素密切相關[60-62]。但是有研究表明,CCD1的表達趨勢并不能準確的反映出揮發性脫輔基類胡蘿卜素的含量變化,其原因可能是CCD1定位于細胞質中,而裂解底物位于質體中[10]。
4 類胡蘿卜素代謝對植物葉片色彩與香味的影響
4.1 類胡蘿卜素相關基因對葉色的影響
葉色是觀賞植物易識別的表型特征,也是重要的觀賞性狀之一。類胡蘿卜素相關基因轉錄水平的改變對葉色的變化起到重要作用,例如在本氏煙草(Nicotiana benthamiana)中,導入番茄PSY的過表達載體,使得葉片積累大量八氫番茄紅素,植株呈亮橙色[63];過表達BoaNXS的轉基因芥藍(Brassica oleracea)植株葉片中新黃質、紫黃質、葉黃素等含量顯著增加,葉色由綠色變為黃綠色[64]。此外,黃葉突變體的產生多與類胡蘿卜素含量變化有關,在銀杏金葉突變體中,上調的Z-ISO、ZDS和LCYE 促進了類胡蘿卜素尤其是葉黃素的積累[65];類胡蘿卜素異構酶(BrCRTISO)的功能喪失導致前番茄紅素積累,使得大白菜(Brassica rapa)內葉呈現橘紅色[66];小麥(Triticum aestivum)黃葉突變體中BCH表達水平較綠葉表型上調,導致總類胡蘿卜素含量顯著降低,而玉米黃質含量升高[67]。
4.2 類胡蘿卜素相關基因對葉片香味的影響
類胡蘿卜素是植物葉片中重要香氣成分的前體物質,目前類胡蘿卜素調控葉片香氣物質產生的研究,僅在茶葉(Camellia sinensis)、煙草(Nicotiana tabacum)以及香露兜(Pandanus amaryllifolius)等植物中被報道。在制茶過程中,CsCCD1a和CsCCD4共同參與裂解β-胡蘿卜素生成β-紫羅酮,最終貢獻了茶葉中的花果香味[68-69]。在煙草葉片中,NtCCD1是催化類胡蘿卜素降解產生醛類與酮類物質的關鍵基因[70],而NtLcy-b基因過表達可以增加β-胡蘿卜素含量,進而提高了β-胡蘿卜素降解產物,如β-二氫大馬酮、β-環檸檬醛等香氣物質的含量[71]。同時,香露兜作為天然香料,在其葉片中鑒定出的α-紫羅蘭酮、β-紫羅蘭酮和β-環檸檬醛等揮發性芳香物質也主要由α-胡蘿卜素和β-胡蘿卜素在類胡蘿卜素裂解酶的催化下生成[72]。
5 基因工程技術在改良植物觀賞器官中類胡蘿卜素的應用
定性或定量改變類胡蘿卜素種類和含量,提高植物觀賞價值是植物育種的重要目標。隨著轉基因技術的不斷發展,人們通過嘗試過表達外源基因,或者抑制內源基因表達和基因敲除等手段對類胡蘿卜素代謝途徑進行人為干預,從而改變類胡蘿卜素的種類或含量以實現植物重要觀賞器官色彩和香味的改良(表1)。
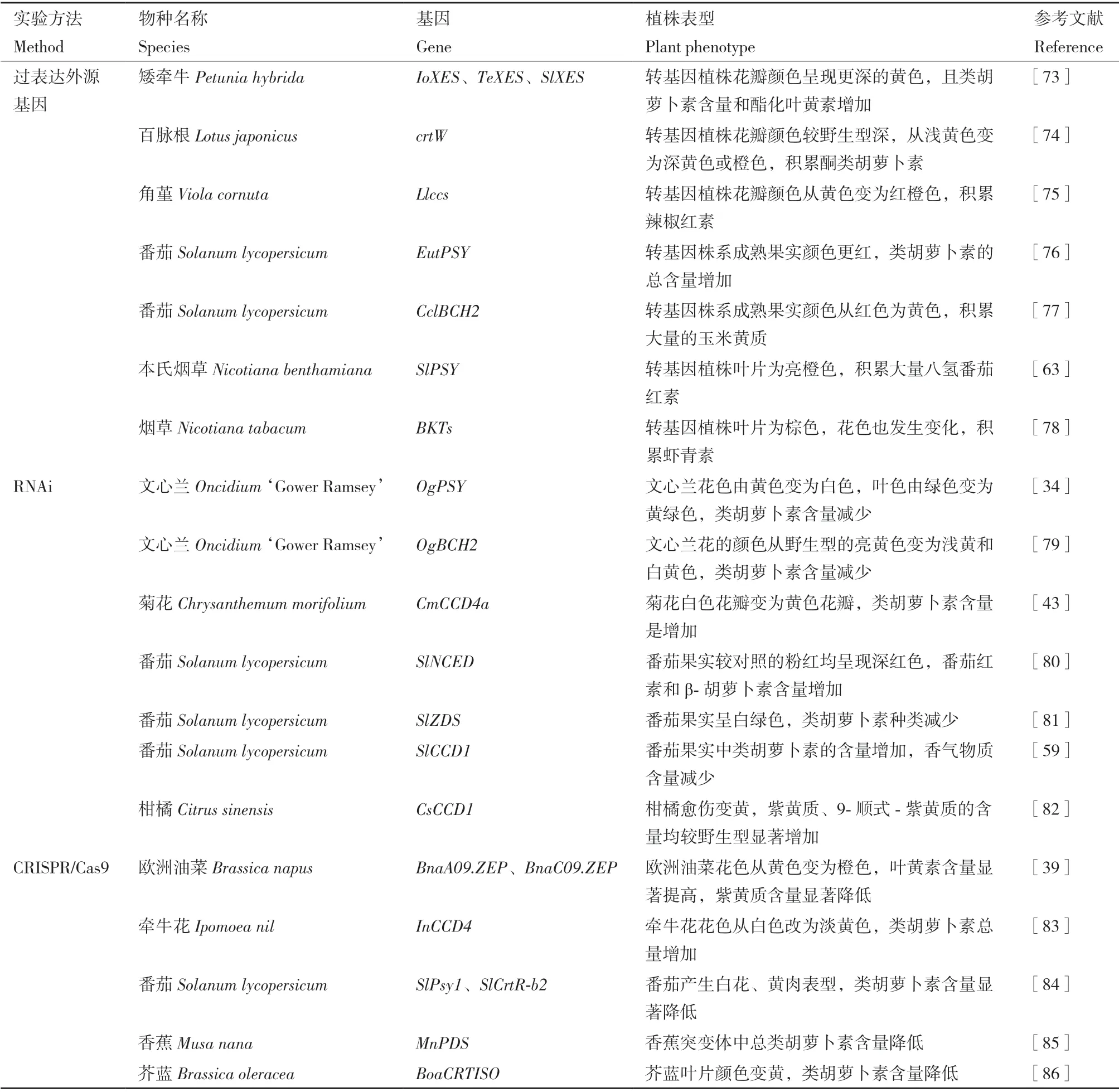
表1 基因工程技術利用類胡蘿卜素代謝相關基因在改植物良觀賞品質的應用Table 1 Application of genetic engineering technology in improving good ornamental characters of plants using carotenoid metabolism related genes
5.1 過表達外源基因
過表達外源基因是植物基因工程的常用技術,通過設計代謝途徑積累特定的類胡蘿卜素,可以使植物產生特定的顏色或香味。蝦青素是一種價值極高的酮類胡蘿卜素,僅在少數生物體中低水平合成,通過葉綠體轉化萊茵衣藻酮酶基因(BKT)的方法,在轉基因煙草葉片中積累蝦青素和其他酮類胡蘿卜素,花色也產生了明顯的改變[78];同時利用農桿菌介導的方法將crtW基因導入百脈根(Lotus japonicus),轉基因植株積累酮類胡蘿卜素,花瓣從淺黃色變為深黃色或橙色[74]。此外,葉黃素酯化可以促進植物花瓣中葉黃素的積累,將小心葉薯(Ipomoea obscura)、番茄和萬壽菊(Tagetes erecta)的葉黃素酯酶基因(XES)引入一個淡黃色花的矮牽牛品種中,類胡蘿卜素含量和酯化葉黃素的比例升高,顏色呈現更深的黃色[73]。綜上所述,導入植物體中缺少或低效表達的外源基因為改變植物觀賞器官中類胡蘿卜素的種類和含量提供了一種獨特而有效的策略。
5.2 RNAi技術
RNA 干擾(RNA interference,RNAi)是一種轉錄后基因沉默現象,利用RNAi 技術,可降低靶基因的表達,而不會徹底敲除目標基因。在文心蘭中,通過RNAi 抑制PSY的表達,花的顏色從亮黃色變為白色或黃白色,葉片呈現黃綠色,且此性狀具有遺傳穩定性[34];下調BCH-A2和BCH-B2的表達,花瓣呈淺黃色和白黃色[79]。在番茄中,利用RNAi干擾SlZDS的表達,β-胡蘿卜素和番茄紅素缺失,果實由紅色變為白綠色,且葉片存在光漂白現象[81];降低SlCCD1的表達能夠提高番茄果實中類胡蘿卜素的含量,同時減少香氣物質的產生[59]。因此,RNAi 技術在定向改良植物性狀中極具應用價值。
5.3 CRISPR/Cas9基因編輯技術
新型基因組編輯技術CRISPR/Cas9 可以通過改變單個基因或一小套基因來操縱重要性狀,與RNAi等基因的抑制技術相比,CRISPR/Cas9 的基因敲除技術加快了分子改良育種的進程。在牽牛花中,通過構建CRISPR/Cas9 系統對InCCD4進行編輯,使得花色由白色到淺黃色的轉變,花瓣中的類胡蘿卜素含量顯著增加[83];在番茄中,敲除Psy1和CrtR-b2,產生白花、黃肉表型[84];在芥藍中,利用CRISPR/Cas9 編輯技術和農桿菌介導的穩定轉化的方法,抑制BoaCRTISO基因的表達,獲得顏色更加鮮艷的黃色芥藍植株[86]。CRISPR/Cas9 技術作為一種應用廣泛的基因編輯技術,為植物觀賞器官色香改良的研究提供了新的方法。
6 展望
作為調控植物觀賞性狀的一個重要因子,類胡蘿卜素不僅是一類天然色素,賦予植物多樣的色彩,而且還是脫輔基類胡蘿卜素合成的前體,對于植物觀賞器官香味的產生有重要的作用。因此研究類胡蘿卜素在調控植物花、果、葉的著色及香氣物質產生等方面具有重要意義,例如研究類胡蘿卜素的調節機制,包括轉錄調控、表觀遺傳調控等調控網絡,能夠更好地揭示植物觀賞器官中類胡蘿卜素代謝的具體機理;挖掘類胡蘿卜素代謝途徑關鍵基因,為利用基因編輯技術定向改良植物色彩和香味提供基因資源。
目前對于類胡蘿卜素的研究大多集中在模式植物以及植物的花和果實中,在葉片中的研究尚少。彩葉的形成機制較為復雜,通常涉及葉綠素、類胡蘿卜素、花青素等多種代謝途徑,因此,如何培育出葉色靚麗的觀賞植物,值得進一步探討。另外,目前利用類胡蘿卜素改良植物觀賞性狀多針對于改變色素沉著,而香味物質產生的研究相對較少,且多著重于類胡蘿卜素的降解,迄今為止,還未解開類胡蘿卜素與單萜和倍半萜等香氣物質合成之間的競爭機制,這仍是一個重要的研究方向。近年來隨著植物分子育種技術的發展,利用基因工程技術改良植物觀賞性狀已經實現,因此深入解析植物觀賞器官中類胡蘿卜素的代謝調控可為定向培育色香俱佳的植物新品種提供了理論支持。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
