‘晉農1號’達烏里胡枝子根瘤內生解磷菌的鑒定及特性研究
2023-01-03 10:17:56楊凱元梁銀萍韓玲娟
草地學報 2022年12期
關鍵詞:能力
楊凱元, 黃 臣, 高 鵬, 梁銀萍, 韓玲娟, 趙 祥
(山西農業大學草業學院, 山西 太谷 030801)
土壤中廣泛分布的根瘤菌可以通過誘導植物根、莖皮層細胞增生形成具有固氮功能的共生體,將空氣中游離態的氮還原成銨態氮,加速植物對氮素的吸收和利用[1]。其中,豆科植物和根瘤菌共生體的固氮量約占生物固氮總量的65%[2],在提高產量、改善品質方面發揮重要作用[3]。根瘤菌固氮是一個高耗能過程,每還原一個N2分子需要消耗16分子三磷酸腺苷(ATP)[4],因此固氮過程對磷元素的需求量大。磷缺乏不僅直接導致豆科植物生長受限,而且抑制了根瘤活性及其固氮能力,造成植物產量和品質的大幅降低[5-6]。研究表明,在低磷條件下,根瘤菌可以通過根瘤表面吸收周圍土壤中的可溶性磷[7],而土壤中95%以上的磷為難溶性磷[8],不能被根瘤菌直接吸收利用。因此,一些兼具解磷能力的根瘤菌或者根瘤內生非結瘤解磷菌,通過構建復合菌群的方式在根瘤中共存,分解周圍的難溶性磷供固氮菌使用[9-10],從而促進結瘤,提高根瘤固氮能力。
目前,已從豆科植物根瘤中分離出許多非結瘤內生菌(Non-rhizobial endophytes,NRE),如葉桿菌屬Phyllobacterium、泛菌屬Pantoea、芽孢桿菌屬Bacillus、假單胞菌屬Pseudomonas和農桿菌屬Agrobacterium等[11],證實不僅具有解磷、聯合固氮的功能[12],而且具有促生、防病等廣泛的生物學作用[13-14]。與根際解磷菌不同,根瘤內生解磷菌能夠為根瘤菌結瘤固氮提供磷[9],目前已有研究通過根瘤菌和根瘤內生解磷菌雙接種試驗,進一步證實了大豆Glycinemax、鷹嘴豆Cicerarietinum等豆科植物的結瘤固氮能力顯著增強,并促進根系生長,顯著提高了作物的產量[15-17]。不同豆科植物與根瘤內生菌建立的共生體中,解磷菌的類型和解磷能力存在差異,趙龍飛等[14]從大豆根瘤內分離出7株解磷能力較強的內生菌,其中惡臭假單胞菌Pseudomonasputida的解磷量最高可達452 μg·mL-1;Rajput等[18]從綠豆Vignaradiata根瘤內分離出1株解木聚糖類芽胞桿菌Paenibacillusxylanilyticus,但解磷量較低,為134.48 μg·mL-1。因此,在豆科植物栽培過程中,挖掘和應用具有較強解磷能力的根瘤內生菌是促進植物結瘤固氮,增加產量,改善品質,保證農業綠色可持續發展的重要途徑。
達烏里胡枝子是豆科(Leguminomase)胡枝子屬(Lespedeza)的多年生植物,能與根瘤菌共生,是山西、陜西等黃土高原地區優良的鄉土牧草和水土保持先鋒植物[19]。目前,國內外有關胡枝子屬植物根瘤內生菌的研究較多,如Palaniappan等[20]從韓國的胡枝子屬植物根瘤中共分離出13個種共39株內生菌,其中24株具有解磷能力;Subramanian等[21]進一步將其中具有解磷能力的巨大芽孢桿菌Bacillusmegaterium與水稻Oryzasativa葉片內生菌微枝桿菌Methylobacteriumoryzae和大豆慢生根瘤菌Bradyrhizobiumjaponicum共接種,發現大豆的結瘤固氮能力顯著提升;Busby等[22]利用高通量測序對比了入侵物種截葉鐵掃帚(Lespedezacuneata)與本地L.virginica、L.repens、L.procumbens、L.capitata、L.stuevei和L.violacea等6個胡枝子種的根瘤內生菌多樣性,發現后者的種類更為豐富,其中伯克氏菌目Burkholderiales和腸桿菌科Enterobacteriaceae細菌為常見的解磷菌。可見,胡枝子屬植物根瘤中存在具有解磷和共生固氮能力的內生菌,且種類與寄主生長的地理位置密切相關。然而,目前尚未見關于山西省種植的達烏里胡枝子根瘤內生菌,尤其是解磷菌的研究報道。本團隊于2014年獲得1個國家審定草新品種‘晉農1號’達烏里胡枝子,該品種適應性強、耐旱、耐貧瘠且品質優良,營養價值豐富,對于黃土高原地區生態修復以及飼草生產發揮著重要作用[23]。本文通過分離和鑒定‘晉農1號’達烏里胡枝子根瘤內生菌,篩選解磷菌并分析其抗逆和促生特性,以期為研發適合于黃土高原地區甚至退化鹽堿地胡枝子建植的根瘤微生物復合菌劑提供菌種資源。
1 材料與方法
1.1 材料
1.1.1根瘤樣品采集 2021年8月中旬,在山西省晉中市太谷區(112°35′25″E,37°26′1″N)的‘晉農1號’達烏里胡枝子種植田采集盛花期[24]的植株樣品,種植田面積為0.3 ha,采用“五點取樣法”在種植田設置5個5 m×5 m的重復小區,每個小區在對角線上每間隔5 m隨機選取3株達烏里胡枝子,共采集15株。用75%乙醇表面消毒的鐵鏟挖取完整的根系土胚,抖落根系表面的土壤,挑選具有根瘤的植株帶回室內。用滅菌剪刀剪取根瘤部位根系,置于75%酒精中消毒30 s,無菌蒸餾水漂洗3次,放入底部盛有干燥劑、中間用滅菌脫脂棉隔離的5 mL離心管中,置于4℃低溫冰箱中保存,3 d內進行根瘤內生菌的分離。
1.1.2培養基 參考不同的文獻方法配制:牛肉膏蛋白胨培養基、YMA(Yeast mannitol agar)培養基、PKO(Pikovaskaia's)無機磷培養基、蒙金娜有機磷培養基[14],TY液體培養基[25],淀粉瓊脂培養基、明膠培養基、葡萄糖蛋白胨液體培養基、1%胰胨水溶液[26],CAS(Chrome azurol S)鐵載體檢測培養基[27],葡萄糖蛋白胨培養基[28]。
培養基均于121℃高溫高壓滅菌20 min,其中明膠培養基、葡萄糖蛋白胨液體培養基、1%胰胨水溶液分裝于試管,TY液體培養基分裝于錐形瓶,其余培養基均制備成平板備用。
1.2 實驗方法
1.2.1達烏里胡枝子根瘤內生菌的分離 使用牛肉膏蛋白胨培養基分離根瘤內生菌。將干燥的根瘤樣品用無菌水浸泡2 h使其完全吸脹,置于95%乙醇中處理30 s,5% NaClO溶液消毒5 min,無菌水沖洗5~7次。用無菌刀片沿根瘤中間部位切開,將根瘤切面在培養基表面劃線,置于28℃恒溫培養箱(MJ-I型,上海一恒公司)中培養。待菌落出現后,挑取單菌落繼代劃線純化培養,直至獲得純菌株,在斜面培養基上4℃低溫保存。
1.2.2根瘤內生解磷菌的初篩及其解磷能力和堿性磷酸酶活性的測定 解磷菌初篩:將保存的根瘤內生菌菌株活化后,制備成菌懸液(OD600=2.0)。吸取5 μL菌懸液,接種在定量20 mL的PKO無機磷培養基和蒙金娜有機磷培養基上,每個菌株4次重復,28℃恒溫培養,第7 d采用“十字交叉法”測定透明圈直徑(D)和菌落直徑(d),將可溶性指數(D/d)≥1.1的解磷菌用于開展后續試驗。
解有機磷和無機磷能力測定:將上述篩選的解磷菌活化后,制備成菌懸液(OD600=0.8)。吸取1 mL菌懸液接種在60 mL蒙金娜液體培養基中,每個菌株3次重復,接種無菌水為對照,28℃、180 r·min-1振蕩培養(ZQLY-180E型,上海知楚公司)5 d,混勻取10 mL培養液,10 000 r·min-1離心(H2050R型,湖南湘儀公司)15 min,采用“鉬銻抗比色法”測定上清液有效磷含量[29],確定菌株解有機磷能力,使用pH計測定上清液pH值[14]。以磷酸鈣為磷源,定量測定菌株解無機磷能力,方法同解有機磷能力測定。將解有機磷和無機磷試驗中上清液有效磷含量顯著高于對照的菌株確定為有效解磷菌。
堿性磷酸酶活性測定:使用堿性磷酸酶(AKP/ALP)活性檢測試劑盒對上述上清液的堿性磷酸酶活性進行測定。主要步驟有,在37℃,遮光條件下與上清液反應15 min,每mL上清液每min催化產生1 μmol酚定義為1個酶活力單位。
1.2.3根瘤內生解磷菌的鑒定 解磷菌形態特征觀察:將篩選的有效解磷菌接種在YMA培養基上,28℃恒溫培養2 d,觀察菌落形態特征,測量菌落直徑;參考伯杰氏細菌系統學手冊(Bergey’s Manual of Systematic Bacteriology,Second Edition,2004)進行革蘭氏染色、芽孢染色,使用顯微鏡(BX50F-3型,日本Olympus公司)觀察菌體形態、測量大小,每個菌株測量50個。
解磷菌的生理生化反應:參考沈萍等[30]的方法,對篩選的有效解磷菌進行接觸酶反應、淀粉水解反應、明膠液化反應和甲基紅反應的測定。
解磷菌的16S rDNA序列相似性與系統發育分析:挑取篩選的有效解磷菌單菌落置于盛有20 μL無菌水的PCR八連排管中,在PCR儀上95℃加熱7 min,離心取上清液作為模板DNA備用。利用通用引物P1(5’-AGA GTT TGA TCC TGG CTC AGA ACG AAC GCT-3’)和反向引物P6(5’-TAC GGC TAC CTT GTT ACG ACT TCA CCC C-3’)對模板DNA進行擴增。PCR反應體系為2×Taq Master Mix 25 μL,上、下游引物(10 μmol/L)各2 μL,DNA模板2 μL,ddH2O 19 μL。PCR反應程序為94℃/3 min;94℃/30 s,55℃/30 s,72℃/1 min,30個循環;72℃/5 min。PCR產物經1%瓊脂糖凝膠電泳檢測合格后,送上海生工生物工程股份有限公司測序。獲得的16S rDNA序列編輯后提交NCBI數據庫獲得登錄號,并利用EZBioCloud(https://www.ezbiocloud.net/identify/)進行序列比對。使用MEGA 7.0軟件將序列與EZBioCloud數據庫中的模式菌株序列進行系統發育分析,采用鄰接法(Neighbor-Joining method)構建系統發育樹,自展值(bootstrap)為1 000次。
1.2.4根瘤內生解磷菌抗逆性測定 有效解磷菌耐鹽、耐酸堿性測定:分別制備NaCl含量為0.01%,0.1%,1%,2%,3%,4%,5%,6%,7%,8%,9%和pH值為4,5,6,7,8,9,10,11,12的YMA培養基,吸取5 μL有效解磷菌菌懸液(OD600=2.0)接種在不同處理的培養基上,每個菌株3次重復,28℃恒溫培養,第3 d測定菌落直徑。
有效解磷菌高低溫度耐受性測定:吸取5 μL有效解磷菌菌懸液(OD600=2.0)接種在YMA培養基上,每個菌株3次重復,分別置于溫度為4℃,20℃,28℃,37℃和60℃的恒溫培養箱中培養,第3 d測定菌落直徑。
1.2.5根瘤內生解磷菌其它促生特性測定 有效解磷菌其它促生特性測定:對有效解磷菌進行產鐵載體能力[25]、產有機酸能力[26]和吲哚反應[28]測定;其中,產鐵載體能力和產有機酸能力測定方法如下,將有效解磷菌分別接種在CAS培養基和葡萄糖蛋白胨培養基上,每個菌株4次重復,28℃恒溫培養,第5 d采用“十字交叉法”測定透明圈直徑(D)和菌落直徑(d),以可溶性指數(D/d)表示菌株產鐵載體和產有機酸能力。
1.3 數據分析
利用Microsoft Excel 2019整理數據,繪制根瘤內生菌屬組成的餅圖;使用SPSS 26.0進行數據分析,對菌株的可溶性指數、液體培養基中有效磷含量、堿性磷酸酶活性以及其它促生特性等指標進行單因素方差分析和LSD法多重比較(P<0.05)。采用Origin 2021 統計軟件繪制根瘤內生解磷菌的解磷能力柱狀圖。
2 結果與分析
2.1 達烏里胡枝子根瘤內生菌屬的組成
從達烏里胡枝子根瘤中分離出88株內生菌,共13屬,其中農桿菌屬Agrobacterium分離頻率最高,為19.3%,短桿菌屬Brevibacterium和近芽孢桿菌屬Peribacillus次之,分離頻率分別為18.2%和17.1%;其它菌株如普里斯特氏菌屬Priestia、根瘤菌屬Rhizobium、慢生根瘤菌屬Bradyrhizobium、芽孢桿菌屬Bacillus、新根瘤菌屬Neorhizobium、黃單胞菌屬Xanthomonas、假單胞菌屬Pseudomonas、鏈霉菌屬Streptomyces、葉桿菌屬Phyllobacterium和泛菌屬Pantoea的分離率均小于10.0%(圖1)。

圖1 達烏里胡枝子根瘤內生菌屬的組成Fig.1 Genus composition of nodule endophytic bacteria of Lespedeza daurica
2.2 達烏里胡枝子根瘤內生解磷菌的初篩及其解磷能力和堿性磷酸酶活性
2.2.1根瘤內生解磷菌的初篩 初步確定88株內生菌中有11株具有解磷能力,其中1株可單解無機磷(圖2B),8株可單解有機磷(圖2A),2株可兼解無機磷和有機磷(圖2)。其中,TG7,TG41,TG42,TG43,TG44,TG47和TG68在蒙金娜有機磷培養基以及TG47和TG68在PKO無機磷培養基上的可溶性指數大于1.1。

圖2 達烏里胡枝子根瘤內生解磷菌的可溶性指數Fig.2 Solubility index of endophytic phosphate-solubilizing bacteria isolated from root nodules of Lespedeza daurica 注:A,根瘤內生解磷菌在蒙金娜有機磷培養基上的可溶性指數;B,根瘤內生解磷菌在PKO無機磷培養基上的可溶性指數;不同小寫字母表示差異顯著(P<0.05)Note:A,Solubility index of nodule endophytic phosphate-solubilizing bacteria on organic phosphorus medium;B,Solubility index of nodule endophytic phosphate-solubilizing bacteria on inorganic phosphorus medium;different lowercase letters indicates significant difference at the 0.05 level. The same as below
2.2.2根瘤內生解磷菌解有機磷和無機磷能力及其堿性磷酸酶活性 TG7,TG41,TG42,TG43,TG44,TG47和TG68在蒙金娜液體培養基培養5 d后,TG41,TG43,TG47和TG68的培養液有效磷含量顯著高于對照(P<0.05),分別比對照提升了128.1%,159.4%,303.1%和168.8%(圖3A)。培養液pH僅TG47比對照高0.03,TG7,TG41,TG42和TG43的培養液pH顯著低于對照(P<0.05),分別比對照下降了0.41,0.33,0.42和0.72(圖3B)。TG47和TG68在PKO無機磷液體培養基培養5 d后,TG47的培養液有效磷含量顯著高于對照(P<0.05),比對照提高了234.39倍(圖4A)。TG47的培養液pH顯著低于對照(P<0.05),比對照下降了1.04(圖4B)。因此,TG41,TG43,TG47和TG68為有效解磷菌。

圖3 根瘤內生解磷菌解有機磷能力(A)、培養液pH(B)以及堿性磷酸酶活性(C)Fig.3 Phosphate-solubilizing ability to organic phosphorus (A),pH of culture solution (B) and alkaline phosphatase activity (C) of endophytic phosphate-solubilizing bacteria isolated from root nodules of Lespedeza daurica

圖4 根瘤內生解磷菌解無機磷能力(A)及培養液pH(B)Fig.4 Phosphate-solubilizing ability to inorganic phosphorus (A) and pH of culture solution (B) of endophytic phosphate-solubilizing bacteria isolated from root nodules of Lespedeza daurica
TG7,TG41,TG42,TG43,TG44,TG47和TG68在蒙金娜液體培養基培養5 d后,TG41,TG42,TG43,TG44和TG47的堿性磷酸酶活性顯著高于對照(P<0.05),分別比對照提升了337.9%,303.5%,466.8%,388.7%和1209.8%(圖3C),其中,TG47的堿性磷酸酶活性顯著高于對照和其它菌株(P<0.05)。
2.3 根瘤內生解磷菌的鑒定
2.3.1達烏里胡枝子根瘤內生解磷菌的形態特征 有效解磷菌的菌落均為圓形、邊緣完整且中心隆起。其中,TG41和TG43菌落白色,生長2 d的菌落直徑分別為6.94和6.99 mm(圖5A-a,b),革蘭氏染色陽性(圖5B-a,b),有芽孢(圖5C-a,b),菌體桿狀,大小分別為(1.06~1.78)×(1.95~4.11)μm和(1.17~1.55)×(2.60~4.00) μm(表1);TG47和TG68的菌落分別為乳白色和黃色,生長2 d的菌落直徑分別為6.77和5.66 mm(圖5A-c,d),革蘭氏染色陰性(圖5B-c,d),無芽孢(圖5C-c,d),菌體直桿狀,大小分別為(0.66~0.75)×(1.40~1.79)μm和(0.55~0.91)×(1.18~2.64)μm(表1)。

圖5 根瘤內生解磷菌的形態特征Fig.5 Morphological characteristics of endophytic phosphate-solubilizing bacteria isolated from root nodules of Lespedeza daurica注:A,有效解磷菌的菌落形態特征,a ~ d分別為TG41,TG43,TG47和TG68的菌落;B,有效解磷菌的革蘭氏染色結果,a ~ d分別為TG41,TG43,TG47和TG68;C,有效解磷菌的芽孢染色結果,a ~ d分別為TG41,TG43,TG47和TG68;標尺=5 μmNote:A,Colony morphological characteristics of effective phosphate-solubilizing bacteria,a ~ d show the colonies of TG41,TG43,TG47 and TG68,respectively;B,Gram staining results of effective phosphate-solubilizing bacteria,a ~ d show the results of TG41,TG43,TG47 and TG68,respectively;C,Spore staining results of effective phosphate-solubilizing bacteria,a ~ d show the results of TG41,TG43,TG47 and TG68,respectively;scale = 5 μm

表1 根瘤內生解磷菌的形態特征Table 1 Morphological characteristics of endophytic phosphate-solubilizing bacteria isolated from root nodules of Lespedeza daurica
2.3.2達烏里胡枝子根瘤內生解磷菌的生理生化特性 TG41和TG43的過氧化氫酶反應、淀粉水解反應和明膠液化反應均呈陽性,甲基紅反應均呈陰性;TG47和TG68過氧化氫酶反應、淀粉水解反應和明膠液化反應均呈陰性;TG47甲基紅反應呈陰性,TG68甲基紅反應呈陽性(表2)。
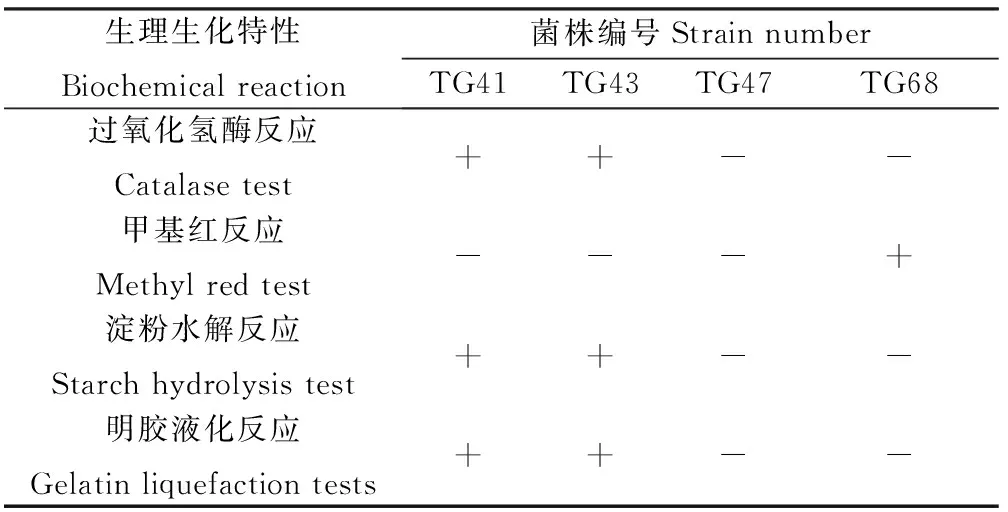
表2 根瘤內生解磷菌的生理生化特性Table 2 Biochemical reaction of endophytic phosphate-solubilizing bacteria isolated from root nodules of Lespedeza daurica
2.3.3達烏里胡枝子根瘤內生解磷菌16S rDNA序列相似性與系統發育分析 TG41,TG43,TG47和TG68的GenBank登錄號分別為ON529568,ON529570,ON529572和ON529573,16S rDNA序列長度分別為1 458 bp,1 460 bp,1 469 bp和1 506 bp。通過EZBioCloud數據庫比對和系統發育分析(圖6),TG41和TG43與模式菌株辣椒芽孢桿菌Bacilluszanthoxyli1433T(登錄號:KX865140)的同源性均為99.58%;TG47與模式菌株尊杜克葉桿菌PhyllobacteriumzundukenseTri 48T(登錄號:KX426256)的同源性為96.28%;TG68與模式菌株成團泛菌PantoeaagglomeransDSM 3493T(登錄號:AJ233423)的同源性為99.10%。結合菌株生理生化特性及培養特征,確定TG41和TG43為辣椒芽孢桿菌Bacilluszanthoxyli,TG47為尊杜克葉桿菌Phyllobacteriumzundukense,TG68為成團泛菌Pantoeaagglomerans。

圖6 根瘤內生解磷菌的最大相似性種系統發育樹Fig.6 Phylogenetic tree of max similar species of endophytic phosphate-solubilizing bacteria isolated from root nodules of Lespedeza daurica
2.4 達烏里胡枝子根瘤內生解磷菌的抗逆性
TG41,TG43和TG68具有較好的耐鹽、耐酸堿能力,分別在鹽濃度0.01%~9.00%和pH4~ pH12的培養基上均可生長;TG47次之,可在鹽濃度2.00%和pH5~pH10的培養基上生長(表3)。

表3 根瘤內生解磷菌在不同脅迫下的菌落直徑Table 3 Colony diameter of endophytic phosphate-solubilizing bacteria isolated from root nodules of Lespedeza daurica under different stresses 單位:mm
所有有效解磷菌在60℃均未生長,不耐高溫;TG47和TG68在4℃仍可生長,具有一定的耐低溫能力。其中,TG41和TG43在20℃~37℃生長良好,TG47和TG68在20℃~28℃生長良好。
2.5 達烏里胡枝子根瘤內生解磷菌的其它促生特性
4株有效解磷菌吲哚反應呈陰性,均具有分泌鐵載體能力,TG47的分泌能力較強;除TG47的3株有效解磷菌均具分泌有機酸的能力,TG68的分泌能力較強(表4)。

表4 根瘤內生解磷菌的其它促生特性Table 4 Other plant growth-promoting characteristics of endophytic phosphate-solubilizing bacteria isolated from root nodules of Lespedeza daurica
3 討論
3.1 達烏里胡枝子根瘤內生菌屬的組成
豆科植物根瘤是植物-微生物共生的特殊微環境,蘊含著豐富的內生菌資源[31]。達烏里胡枝子具有較高的飼用價值和水土保持能力,是山西省的優良鄉土植物,其根瘤內生菌具有較高的研究價值。Palaniappan等[20]從韓國的胡枝子屬植物根瘤中分離出39株內生菌,其中伯克氏菌屬Burkholderia、慢生根瘤菌屬Bradyrhizobium和根瘤菌屬Rhizobium為優勢屬;Busby等[22]對北美6種胡枝子屬植物的根瘤進行高通量測序發現,其內生菌種類豐富,共63屬,其中慢生根瘤菌屬Bradyrhizobium、類固醇桿菌屬Steroidobacter和紅微菌屬Rhodomicrobium為優勢屬;冀玉良等[32]對商洛地區多花胡枝子分離的根瘤內生菌進行系統發育分析,發現根瘤菌屬Rhizobium和中華根瘤菌屬Sinorhizobium為優勢屬。本研究從山西省種植的達烏里胡枝子根瘤中分離出88株內生菌,共13屬,其中根瘤菌屬Rhizobium和(或)慢生根瘤菌屬Bradyrhizobium等在已報道的胡枝子屬植物根瘤中均有發現。包括胡枝子在內的豆科植物上,這些根瘤菌也是形成根瘤,產生固氮功能的必要物種[33]。與上述研究有所不同,本研究中農桿菌屬Agrobacterium、短桿菌屬Brevibacterium和近芽孢桿菌屬Peribacillus為優勢屬,這表明不同地區的胡枝子屬植物根瘤內生菌多樣性豐富,但優勢屬存在差異,這可能是胡枝子物種、生長的地理位置、氣候和土壤條件[34]以及人類活動[35]共同作用所致。此外,本研究還分離出胡枝子屬植物根瘤中尚未報道的內生菌,包括新根瘤菌屬Neorhizobium、短桿菌屬Brevibacterium、近芽孢桿菌屬Peribacillus、普里斯特氏菌屬Priestia、假單胞菌屬Pseudomonas、鏈霉菌屬Streptomyces、葉桿菌屬Phyllobacterium和泛菌屬Pantoea等,極大地豐富了胡枝子屬植物的根瘤內生菌資源庫。
3.2 達烏里胡枝子根瘤內生解磷菌的類型、解磷能力及解磷方式
不同功能的內生菌以構建復合菌群的方式在根瘤內共存[10],其中包括兼具固氮和解磷能力的內生菌[36]和具有解磷能力的非固氮內生菌[40]等,能夠分解土壤中的難溶性磷供其自身構建細胞和植物生長利用[37]或者分解根瘤內部的難溶性磷供根瘤菌利用[40]。陳曉琳等[38]從花櫚木Ormosiahenryi根瘤中分離篩選出1株解磷能力較強的內生菌,解有機磷和無機磷量分別為71.2和31.1 μg·mL-1,固氮酶活性為2.54 μmol·g-1·h-1,經回接試驗確定為根瘤菌;韓梅等[39]從大豆根瘤中篩選出1株兼具固氮和解磷能力的快生型根瘤菌,第7 d解磷量為35.5 μg·mL-1;本研究中TG47和TG68兼具固氮(未發表數據)和解磷能力,經鑒定分別為尊杜克葉桿菌Phyllobacteriumzundukense和成團泛菌Pantoeaagglomerans,表明達烏里胡枝子根瘤中還存在除根瘤菌以外的其它兼具固氮能力的解磷菌,這可能是在協同進化過程中發生了共生基因的橫向轉移[41]。此外,根瘤中還存在具有解磷能力的非固氮內生菌,Mohsin等[42]從豌豆Pisumsativum根瘤中篩選出4株具有解磷能力的非固氮內生菌,解無機磷量為5.57~11.73 μg·mL-1,分別為蒼白桿菌屬Ochrobactrum和腸桿菌屬Enterobacter;鐘宇舟等[43]從大豆根瘤中分離了5株為非固氮內生菌,解磷量為7.47~10.62 μg·mL-1,分別為芽孢桿菌屬Bacillus、腸桿菌屬Enterobacter和假單胞菌屬Pseudomonas;本研究中TG41和TG43解有機磷量為0.73和0.83 μg·mL-1,無固氮能力,經鑒定均為辣椒芽孢桿菌Bacilluszanthoxyli,這表明不同豆科植物與根瘤內生菌建立的共生體中,解磷菌的類型和解磷能力存在差異。本研究已從胡枝子根瘤中分離出具有解磷能力的內生菌,其能否為根瘤菌固氮作用提供磷元素,這還需要通過同位素標記等方法,對內生菌磷轉化和吸收的途徑進行進一步研究。

3.3 達烏里胡枝子根瘤內生解磷菌的抗逆和促生潛力
達烏里胡枝子是黃土高原地區的代表性植物之一,能與根瘤菌共生,具有良好的環境適應性[19]。內生菌在與宿主植物長期協同進化過程中,能夠通過誘導系統抗性以及增強對環境脅迫的耐受性來促進植物生長[49]。本研究從達烏里胡枝子根瘤內分離的芽孢桿菌屬Bacillus和泛菌屬Pantoea具有解磷能力和良好的耐鹽、耐酸堿能力,并且與Palaniappan等[21]的研究相比,本研究中達烏里胡枝子根瘤內生解磷菌的屬組成也更加豐富,這可能是達烏里胡枝子具有良好適應性的原因之一。此外,在豆科植物-根瘤菌共生體中,鐵參與固氮酶、豆血紅蛋白和鐵氧化還原蛋白等關鍵物質的合成[50],對于維持根瘤的固氮活性具有重要意義[51],本研究篩選的4株有效解磷菌兼有分泌鐵載體能力,可螯合周圍環境中的鐵改善自身營養狀況,還能供給植物或與病原微生物競爭鐵營養,達到促生防病的目的[52],但其能否為根瘤菌固氮酶等關鍵物質的合成提供鐵元素仍有待進一步研究。因此,4株有效解磷菌具有較好的抗逆能力和促生潛力,為黃土高原甚至退化鹽堿地建植達烏里胡枝子使用的根瘤微生物復合菌劑提供了菌種資源。因此,后續將進一步開展根瘤菌屬Rhizobium、慢生根瘤菌屬Bradyrhizobium和新根瘤菌屬Neorhizobium的結瘤固氮能力及田間促生效果研究,并進行根瘤內生解磷菌與根瘤菌的復合接種試驗,探究其對根瘤菌結瘤固氮所發揮的作用。
4 結論
山西省種植達烏里胡枝子的根瘤內生菌資源豐富,分屬13屬,除豆科植物形成根瘤的常見根瘤菌屬Rhizobium和慢生根瘤菌屬Bradyrhizobium外,農桿菌屬Agrobacterium、短桿菌屬Brevibacterium和近芽孢桿菌屬Peribacillus也是其優勢根瘤內生菌。其中,4株為有效解磷菌,分別為辣椒芽孢桿菌Bacilluszanthoxyli(TG41和TG43)、尊杜克葉桿菌Phyllobacteriumzundukense(TG47)和成團泛菌Pantoeaagglomerans(TG68),解有機磷量為0.73~1.29 μg·mL-1,TG47和TG68解無機磷量為72.97和1.29 μg·mL-1,菌株分泌磷酸酶為解磷的主要方式。4株解磷菌均能分泌鐵載體,TG41、TG43和TG68還能分泌有機酸,具有一定的促生潛力。此外,這些菌株具有較強的耐鹽和耐酸堿能力,可用于后續研發適合黃土高原甚至退化鹽堿地建植達烏里胡枝子使用的根瘤微生物復合菌劑。
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
中學生數理化·七年級數學人教版(2022年6期)2022-06-05 06:50:58
意林(兒童繪本)(2020年2期)2021-01-07 02:12:04
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
作文成功之路·小學版(2020年5期)2020-06-11 12:48:46
意林(兒童繪本)(2019年9期)2019-10-15 08:51:46
中國生殖健康(2019年10期)2019-01-07 01:21:14
人大建設(2018年6期)2018-08-16 07:23:10
新高考(英語進階)(2018年1期)2018-04-18 14:00:11
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
