大粒香水稻葉綠體基因組特征分析
2022-12-30 06:58:52吳朝昕劉雪薇李祖軍龍武華宮彥龍朱速松
廣西植物 2022年11期
關鍵詞:物種
吳朝昕, 劉雪薇, 李祖軍, 龍武華, 宮彥龍, 朱速松
(貴州省農業科學院 水稻研究所, 貴陽 550006 )
葉綠體是植物獲取能量的重要細胞器,具有獨立于核基因組的遺傳體系。因為其基因序列保守,基因結構重排事件遠低于核基因組,結構簡單,一般為母系遺傳,所以被用來揭示物種的進化與親緣關系(李緒英等,2011;Li et al., 2018; Zhang et al., 2018; Jeon & Kim, 2019;朱斌等,2021)。由于發達的測序科技使不少生物實現了葉綠體基因組(cpDNA)測序,因此NCBI中收錄的葉綠體基因逐漸增多,利用葉綠體基因研究親緣關系的報道也不斷增多。在水稻中,Fan等(2020)利用cpDNA分析了33個稻屬物種的親緣關系,結果顯示OryzasativavoucherHSAGSDYD1802與O.sativacultivar TN1、O.sativacultivar RP Bio-226和O.sativacultivar IR8的親緣關系最近。Fang等(2017)通過分析cpDNA探討了根莖野生稻與其他13個稻屬物種的親緣關系,結果顯示與根莖野生稻關系較近的是藥用野生稻且屬于CC基因類型。在其他物種中,張慧等(2018)利用cpDNA分析了益母草及其他16個物種的親緣關系,研究結果很好地解決了野芝麻亞科的進化關系。鄭祎等(2020)用大花君子蘭葉綠體基因序列與10個百合科、5個蘭科、4個鳶尾科及5個石蒜科共24個物種的葉綠體基因組序列進行系統發育分析,研究結果支持大花君子蘭屬于石蒜科,并使用其中23個物種葉綠體基因組中ycf2進行親緣關系分析發現,葉綠體基因組中ycf2可以代替葉綠體基因組全長進行親緣關系分析。
大粒香是貴州省水稻研究所選育,具有稻米粒大且香的特點,并且大粒香在鄉村振興過程中為社會帶來了較高的經濟效益(蔣志謙,2008;羅仁發等,2012)。目前,對大粒香基因組學、品質形成等理論研究的文獻報道不多。基于大粒香在貴州優質稻發展過程中的重要性,為進一步從基因組水平認識和改良大粒香,本研究以大粒香DNA為材料進行測序,構建大粒香cpDNA圖譜,分析密碼子的使用和重復序列,并分析其親緣關系,擬解決大粒香葉綠體基因組以下問題:(1)大粒香cpDNA的基本特征大小;(2)大粒香cpDNA密碼子偏好情況;(3)大粒香cpDNA系統發育所屬分支。
1 材料與方法
1.1 材料
以大粒香為材料,2019年冬季種植于海南三亞師部農場基地,2020年2月選取無病蟲害、長勢良好的水稻葉片,沖洗、擦干液氮速凍暫存,用干冰保存寄回貴州,置于-80 ℃保存,用于提取大粒香DNA。
1.2 方法
1.2.1 大粒香DNA 提取及測序 使用TIANGEN植物DNA試劑盒提取大粒香總DNA,測序公司檢測合格后,構建文庫,進行測序。
1.2.2 cpDNA序列組裝與注釋 將原始數據,除去有污染、低質量的片段,使用SPAdes軟件拼接后組裝。使用CPGAVAS2進行基因注釋,利用OGDRAW繪制大粒香的cpDNA圖譜。
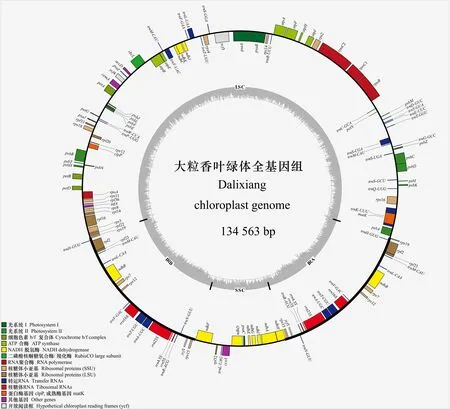
圖 1 大粒香葉綠體基因組圖譜Fig. 1 Gene map of Dalixiang chloroplast genome
1.2.3 密碼子使用分析 使用CodonW進行密碼子使用分析。
1.2.4 重復序列分析 使用Vmatch完成大粒香cpDNA的長重復序列的查找。大粒香cpDNA的SSR篩選則使用MISA軟件,該軟件的檢測參數:單核苷酸大于8時被檢測;二核苷酸和三核苷酸大于4時被檢測。
1.2.5 系統進化分析 為探究大粒香與其他稻屬物種的親緣關系,從NCBI中下載了12個稻屬物種和2種禾本科近源物種的cpDNA(綠竹和高粱),使用RaxML軟件構建系統發育樹。
2 結果與分析
2.1 大粒香cpDNA序列特征
大粒香cpDNA全長為134 563 bp,分為3個區:大單拷貝區(large single copy,LSC)(80 864 bp),GC含量為37.09%;小單拷貝區(small single copy,SSC)(12 347 bp),GC含量為33.37%;反向重復序列區(inverted repeats,IRs)(20 676 bp),GC含量為44.41%。在大粒香cpDNA中注釋129個基因(表1),可分為三類,即蛋白編碼基因、tRNA基因和rRNA基因,其數量分別為85、36和8。其基因功能主要為與自身復制能力有關、與光合作用有關、與其他基因和與未知功能有關4種。在蛋白編碼基因中,rps基因的數量最多有16個,而cemA、infA、ccsA、rbcL、matK、accD、clpP等基因數量僅有1個。其中,有20個基因出現在IR重復區內,分別為ndhB、ycf1、rps12、rps7、rps15、rps9、rpl2、rpl23 8個蛋白編碼基因,rrn23S、rrn4.5S、rrn16S、rrn5S4個核糖體RNA,trnN-GUU、trnH-GUG、trnA-UGC、trnL-CAA、trnT-CGU、trnV-GAC、trnR-ACG、trnM-CAU8個轉運RNA(表1)。統計結果顯示,大粒香cpDNA中有內含子的基因共17個,其中ycf3有2個,剩余16個基因只有1個。trnK-UUU的內含子堿基數最多,而trnL-UAA的最少(表2)。
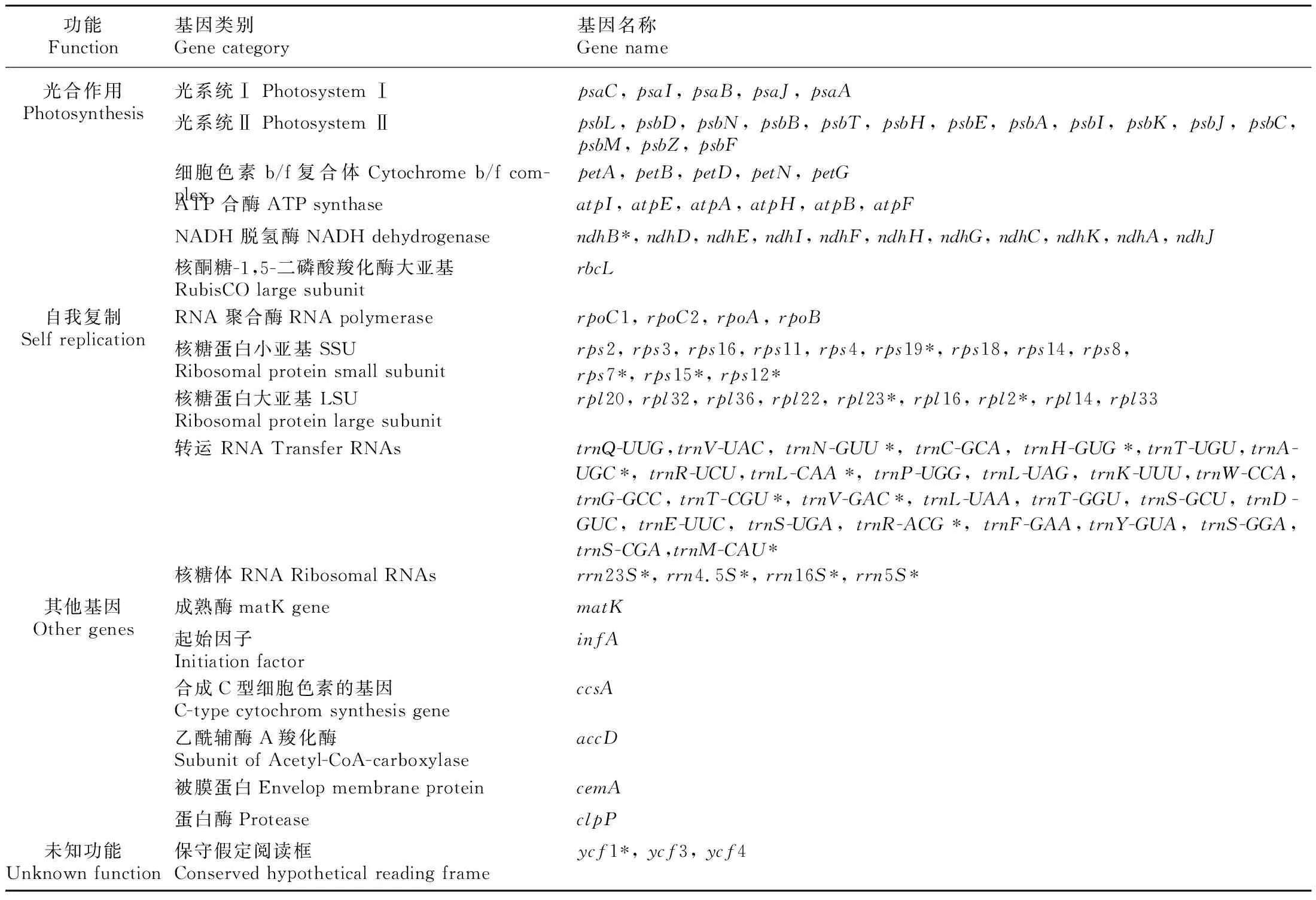
表 1 大粒香cpDNA注釋基因列表Table 1 List of genes found in Dalixiang cpDNA
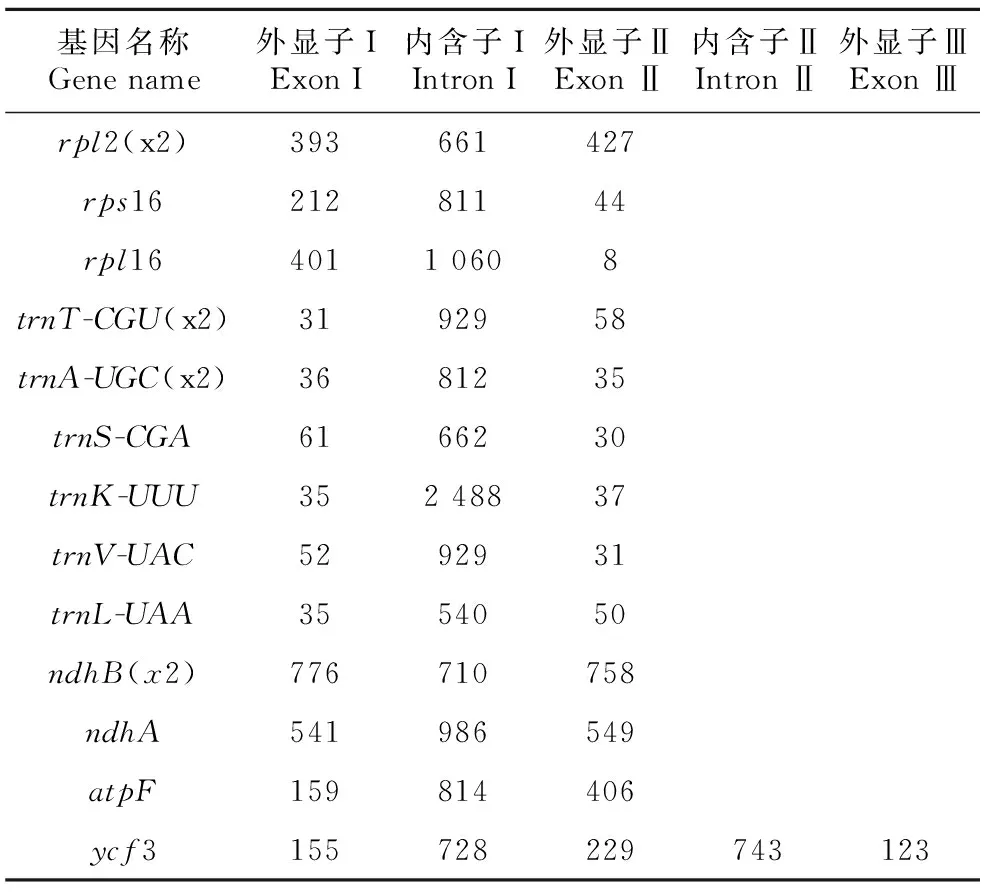
表 2 大粒香cpDNA含有內含子的基因Table 2 Genes with introns in cpDNA of Dalixiang
2.2 大粒香cpDNA密碼子使用分析
在大粒香cpDNA密碼子中,亮氨酸密碼子使用了1 944次,為最多;半胱氨酸的密碼子僅使用198次,為最少。在編碼大粒香cpDNA的密碼子中有30個密碼子偏好性>1,其中以A結尾的有12個,以U(T)結尾的有16個,這表明大粒香cpDNA密碼子偏好A/U(T)堿基,這種情況常出現在杜梨、益母草等多種高等植物中(張慧等,2018;李泳潭等,2020;鄭祎等,2020)(表3)。
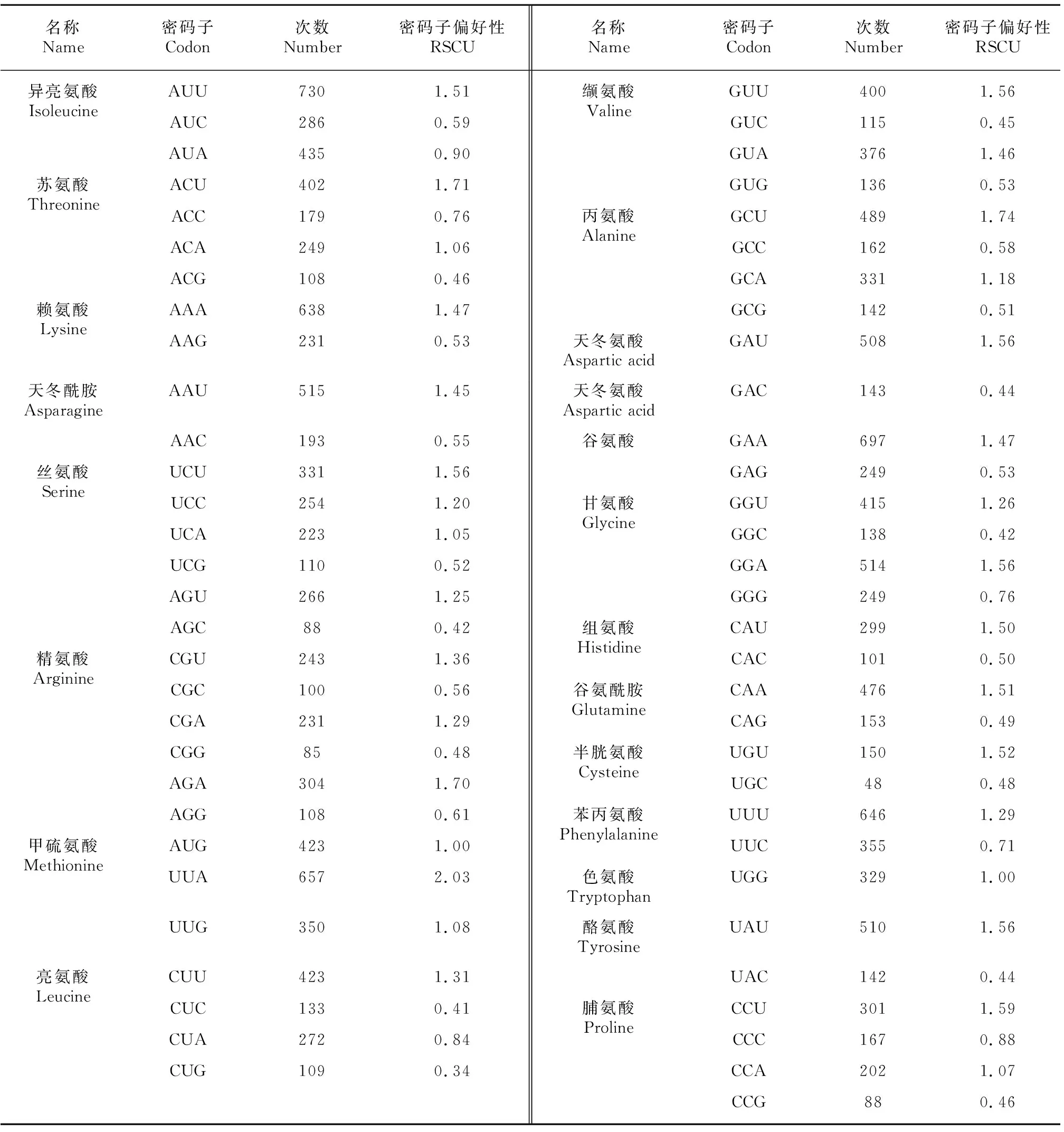
表 3 大粒香cpDNA密碼子使用Table 3 Codon usage in cpDNA of Dalixiang
2.3 大粒香cpDNA長重復序列和SSR分析
在大粒香cpDNA中檢測到19個長重復序列,包含了8個正向重復,長度范圍為30~52 bp,以及11個回文重復,其長度范圍為30~127 bp。最長的127 bp的重復序列位于rps19-psbK的基因間隔區內,而含有最多長重復序列的區間為racL-accD。區域位置分布顯示,絕大多數分布在基因間隔區內(表4)。
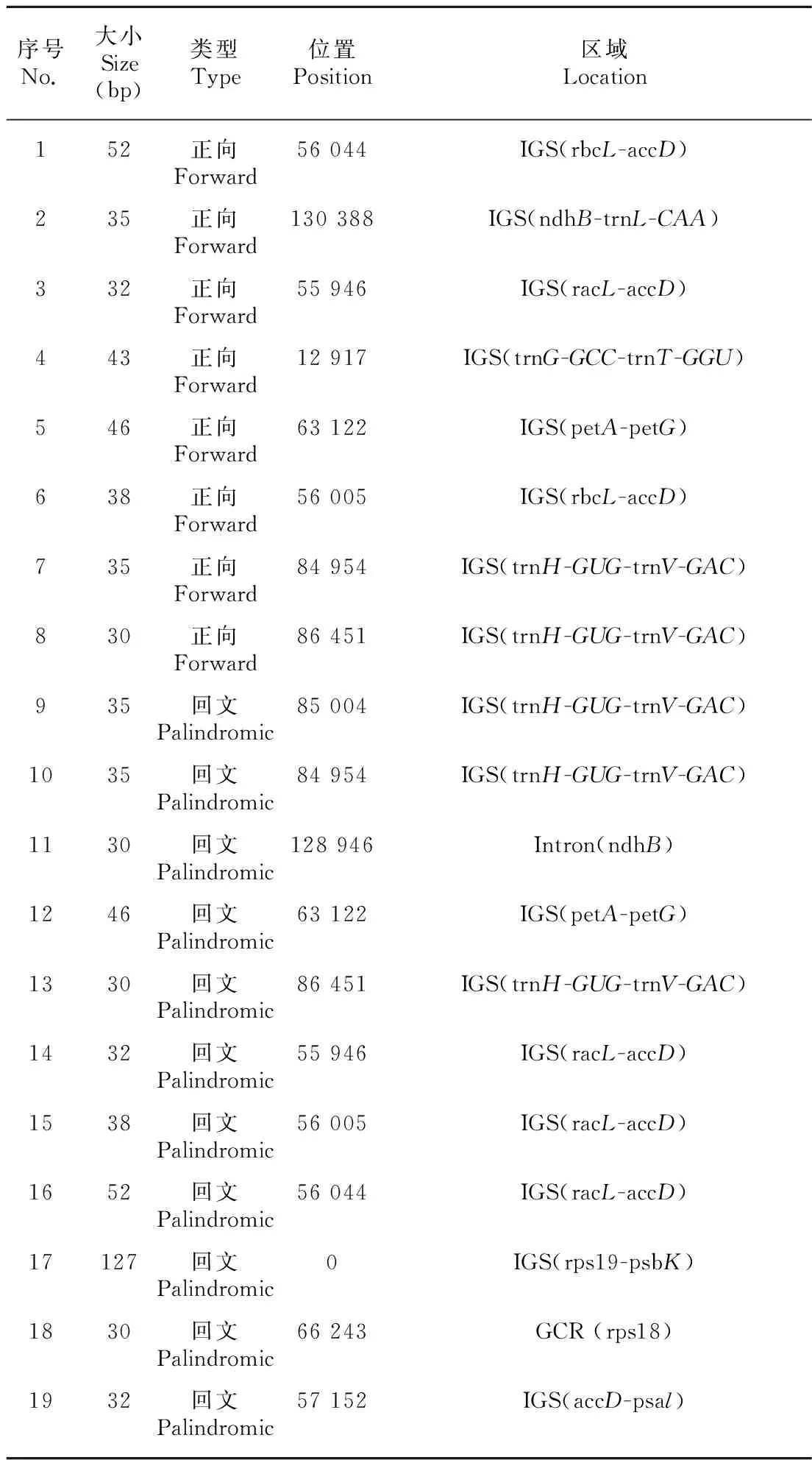
表 4 大粒香cpDNA的重復序列Table 4 cpDNA repeat sequence of Dalixiang
在大粒香cpDNA的129個SSR位點中有95個單核苷酸重復,并且70.07%的SSR由A或T組成,表明SSR位點有使用A/T堿基的偏好。同時,研究表明SSR位點在大粒香cpDNA上分布不均,在LSC區、SSC區以及IRs區分別分布了95個、18個和16個SSR位點(表5)。
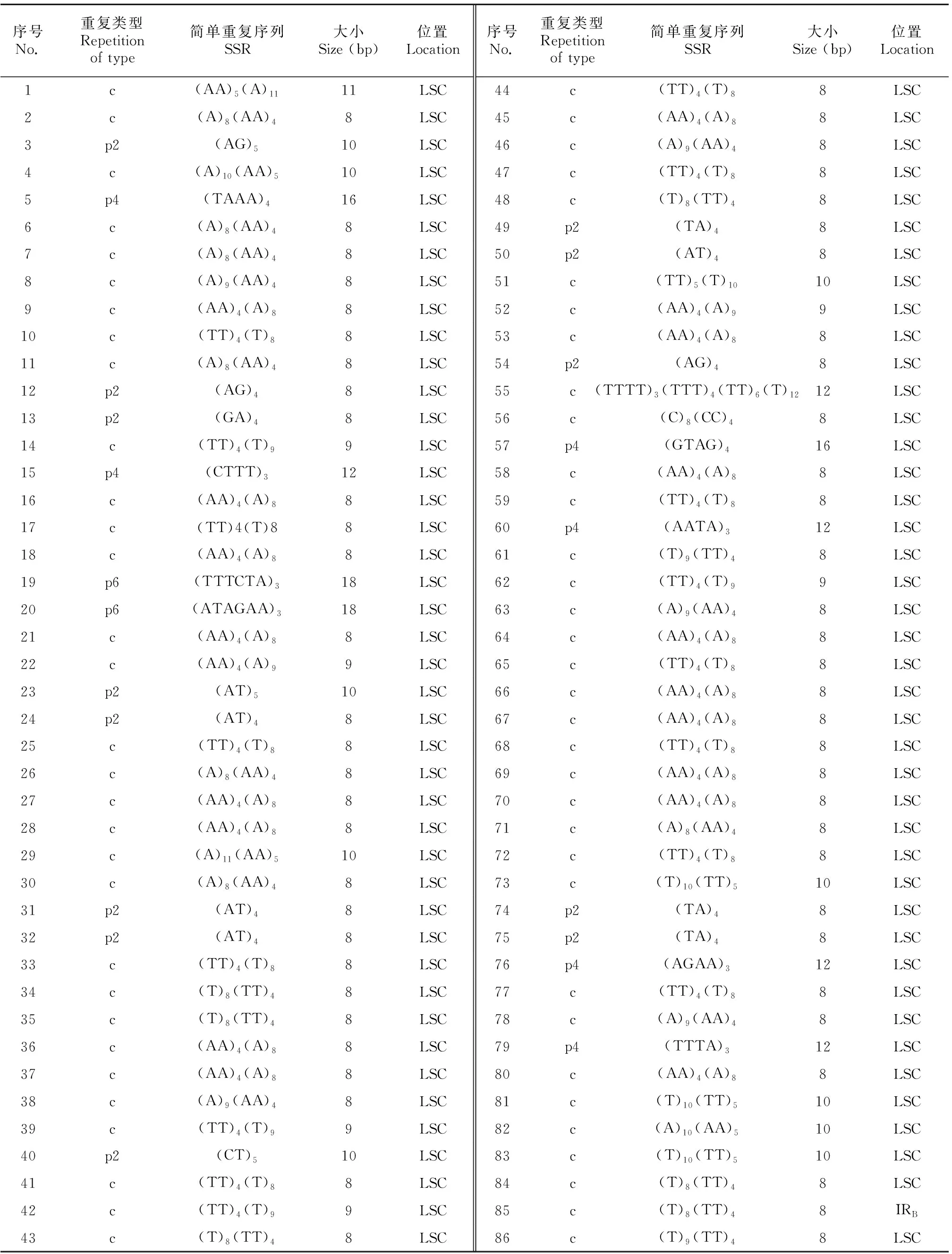
表 5 大粒香cpDNA中的簡單重復序列Table 5 SSR in the cpDNA of Dalixiang
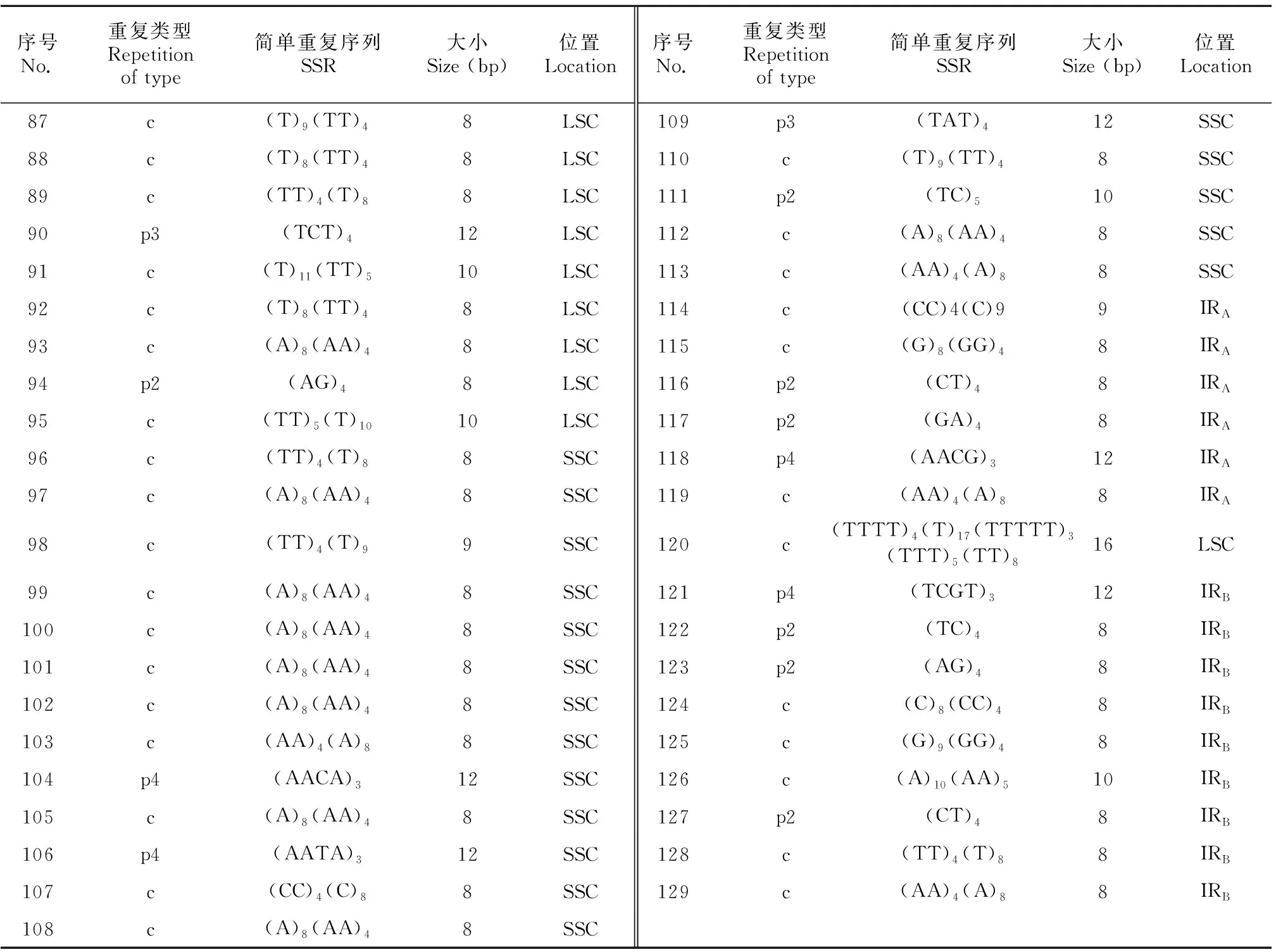
續表5
2.4 大粒香cpDNA系統發育分析
將大粒香與粳稻、秈稻、野生稻及2個外類物種等共15個cpDNA序列構建發育樹。發育樹分析表明,15個物種可分為三類,即第一類為Bambusaoldhamii,第二類為Sorghumbicolor,第三類為13個稻屬物種組成。在第三類群中又可分為4個小類群,其中3種野生稻(O.australiensis300316、O.meridionalis、O.ruifipogon)各為一類,其余10種栽培稻為一類。在栽培稻類群中,粳稻與秈稻分別處于不同進化分支。并且,大粒香水稻與粳稻Tropical Japonica在同一分支,表明兩者的進化關系比其他水稻品種近(圖2)。

圖 2 15種植物的cpDNA序列的系統進化樹Fig. 2 Phylogenetic tree constructed using cpDNA of 15 plants
3 討論與結論
本研究測得大粒香cpDNA全長為134 563 bp,GC含量為39%,LSC為80 864 bp,SSC為12 347 bp,IR為20 676 bp,并注釋到129個基因,與已報道的禾本科數據相符(李裕華等,2020)。前人通過比對不同禾本科植物cpDNA序列表明,雖然葉綠體基因保守程度較高,但一些基因在進化過程中仍然出現退化缺失現象(唐萍等,2011;付濤等,2016)。本研究將大粒香cpDNA與Wang等(2016)報道的熱帶粳稻葉綠體基因組進行比對結果顯示,雖然二者在細胞色素b/f復合體相關基因、光系統Ⅰ、Ⅱ相關基因、核糖體蛋白大和小亞基相關基因、tRNA和未知功能基因等基因差異較少,但在大粒香葉綠體基因中不存在lhbA基因。lhbA基因是和光合作用過程中光系統Ⅱ有關的基因,在熱帶粳稻中存在lhbA基因,并且在喜好溫暖的禾本科植物毛竹中同樣也存在lhbA基因(Yao et al., 2016),這可能是熱帶粳稻為適應熱帶光溫生長環境中逐漸進化而得。
SSR位點可被用于輔助育種和遺傳連鎖作圖等方面的研究,而cpDNA具有序列保守、結構穩定、易測序等優點,有助于解決類群間的遺傳多樣性(Powell et al., 1995; Pugh et al., 2004;Song et al., 2019)。本研究結果表明,大粒香cpDNA的SSR位點對A/U(T)堿基有著明顯偏好,這與前人研究結論相一致(張慧等,2018;李泳潭等,2020;鄭祎等,2020;王一麾等,2021;吳朝昕等,2021)。此外,大粒香cpDNA的密碼子也偏好A/U(T)堿基,這種密碼子使用情況也存在于其他物種中(Shinozaki et al., 1986;Ohyama et al., 1988;鄭祎等,2020;朱斌等,2021;吳朝昕等,2021)。大粒香cpDNA編碼蛋白質的密碼子和SSR位點都偏好A堿基或者U(T)堿基,可能是造成大粒香cpDNA總A/U(T)含量大于總GC含量的原因。根據Niu等(2007)的研究報道,因為A/T核苷酸含有7個氮原子,比G/C核苷酸少一個, 所以富含A/T核苷酸耗能更少,有利于cpDNA的復制。這可能是大粒香cpDNA富含A/U(T)的原因。
系統發育基因組學是利用分子數據來研究生物間發育關系的。由于cpDNA具有序列保守、結構穩定、易測序等優點,因此基于cpDNA進行的系統發育研究得到了很好的發展(Eisen,1998; Eisen & Hanawalt,1999; Delsuc et al., 2005)。本研究對優質稻大粒香cpDNA測序數據進行了系統發育分析,研究結果表明大粒香與熱帶粳稻聚為一類;粳稻與秈稻不為一類,這一結果與林張翔等(2014)的研究結果相同,支持了Huang等(2012)的秈粳稻起源假說。
綜上所述,本研究所獲得的大粒香的cpDNA大小、結構、基因數量、重復序列、密碼子偏好、系統發育樹等特征信息,為進一步研究大粒香的系統進化和育種研究提供了理論依據。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
