羊棲菜多酚通過核轉(zhuǎn)錄因子-κB/絲裂原活化蛋白激酶通路緩解脂多糖誘導(dǎo)的RAW264.7細(xì)胞炎癥反應(yīng)
2022-12-30 06:16:58洪靜霞張攀學(xué)楊文鴿
食品科學(xué) 2022年23期
黃 平,洪靜霞,米 杰,張攀學(xué),李 超,楊文鴿
(寧波大學(xué)食品與藥學(xué)學(xué)院,浙江省動(dòng)物蛋白食品精深加工技術(shù)重點(diǎn)實(shí)驗(yàn)室,浙江 寧波 315211)
炎癥是宿主應(yīng)對(duì)感染、細(xì)胞應(yīng)激或組織損傷的一種防御適應(yīng)性反應(yīng),受多種信號(hào)通路調(diào)控,以保證炎癥過程的啟動(dòng)、維持和消退[1-2]。當(dāng)機(jī)體免疫反應(yīng)異常活躍,可引起過度炎癥,導(dǎo)致細(xì)胞和組織損傷[3]。巨噬細(xì)胞是先天免疫系統(tǒng)的重要組成部分,通過執(zhí)行多種功能(吞噬、產(chǎn)生和分泌多種細(xì)胞因子和生長因子)參與免疫調(diào)節(jié)[4]。脂多糖(lipopolysaccharide,LPS),又稱內(nèi)毒素,為革蘭氏陰性菌細(xì)胞壁外壁的組成成分,可作用于膜受體激活多種細(xì)胞(巨噬細(xì)胞、上皮細(xì)胞、內(nèi)皮細(xì)胞等)信號(hào)轉(zhuǎn)導(dǎo)系統(tǒng),促進(jìn)促炎細(xì)胞因子和其他炎性介質(zhì)的合成及釋放,引發(fā)炎癥反應(yīng)[5]。LPS激活巨噬細(xì)胞的過程主要包括:巨噬細(xì)胞膜上Toll樣受體4(toll-like receptor 4,TLR4)識(shí)別LPS,使TLR4胞內(nèi)的基團(tuán)構(gòu)象發(fā)生變化,將信號(hào)傳入胞內(nèi),激活細(xì)胞中多種蛋白質(zhì)磷酸化級(jí)聯(lián)反應(yīng)鏈,最終激活絲裂原活化蛋白激酶(mitogen activated protein kinases,MAPKs)和核轉(zhuǎn)錄因子(nuclear factor,NF)-κB信號(hào)通路,促進(jìn)多種炎癥介質(zhì)的基因轉(zhuǎn)錄和蛋白表達(dá),如一氧化氮(NO)和前列腺素E2(prostaglandin E2,PGE2)及其上游炎癥酶誘導(dǎo)型一氧化氮合酶(inducible nitric oxide synthase,iNOS)和環(huán)氧合酶-2(cyclooxygenase-2,COX-2)、腫瘤壞死因子(tumor necrosis factor,TNF)-α、白細(xì)胞介素(interleukin,IL)-1β和IL-6等[6-7]。
羊棲菜(Hizikia fusiformis)作為一種可食性褐藻,廣泛分布于我國浙江沿海一帶,富含多種活性物質(zhì),營養(yǎng)價(jià)值高[8]。羊棲菜多酚(Hizikia fusiformispolyphenols,HFPs)是羊棲菜中重要活性物質(zhì)之一,約占藻體干質(zhì)量的2%,HFPs主要為間苯三酚衍生物及連苯三酚衍生物,具有抗氧化[9]、抗菌[10]、抗炎[11]及抑制代謝酶[12]等作用。倪立穎等[11]采用LPS刺激胚胎期斑馬魚建立體內(nèi)炎癥模型,評(píng)估HFPs提取物的抗炎活性,結(jié)果表明HFPs提取物顯著降低胚胎斑馬魚體內(nèi)NO水平、細(xì)胞死亡率和活性氧生成率。Yang等[13]在研究HFPs體外抗炎功效時(shí)發(fā)現(xiàn),其減少了巨噬細(xì)胞促炎因子(NO、IL-6和TNF-α)的分泌。目前,有關(guān)HFPs抗炎作用的報(bào)道主要集中于其對(duì)部分炎性介質(zhì)的影響,而缺乏系統(tǒng)而深入的機(jī)制研究。本研究采用LPS誘導(dǎo)RAW264.7巨噬細(xì)胞建立體外炎癥模型,基于MAPKs和NF-κB信號(hào)通路,從基因、蛋白相對(duì)表達(dá)水平和細(xì)胞吞噬功能等方面評(píng)估HFPs的抗炎活性,為其在新型抑炎制劑的開發(fā)利用提供理論依據(jù)。
1 材料與方法
1.1 材料與試劑
羊棲菜干制品(產(chǎn)自浙江洞頭海域) 溫州星貝海藻食品有限公司。
小鼠巨噬細(xì)胞RAW264.7 中國科學(xué)院細(xì)胞庫。
達(dá)爾伯克(氏)改良伊格爾(氏)培養(yǎng)基(Dulbecco’s modified Eagle medium,DMEM)、DMEM/F12無酚紅培養(yǎng)基、胎牛血清 美國HyClone公司;噻唑藍(lán)(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide,MTT)、LPS、二喹啉甲酸(bicinchoninic acid,BCA)蛋白濃度測(cè)定試劑盒、5×蛋白上樣緩沖液、高效RIPA細(xì)胞快速裂解液 北京索萊寶科技有限公司;N-(1-萘基)乙二胺鹽酸鹽、對(duì)氨基苯磺酰胺 上海源葉生物科技有限公司;TRIzol試劑 美國Omega公司;綠色熒光微球 北京百靈威科技有限公司;Taq-based聚合酶鏈?zhǔn)椒磻?yīng)(polymerase chain reaction,PCR)試劑盒 日本ToYoBo公司;FastStart Essential DNA Green Master 美國Roche公司;c-Jun N末端激酶(c-Jun N-terminal kinase,JNK)兔單克隆抗體、p-JNK兔單克隆抗體、p38 MAPK兔多克隆抗體、p-p38 MAPK兔多克隆抗體、NF-κB p65兔單克隆抗體、p-NF-κB p65兔單克隆抗體、iNOS兔多克隆抗體 武漢愛博泰克生物科技公司;Tris緩沖鹽溶液+吐溫(Tris buffered saline+tween,TBST)洗膜液 山東思科捷生物技術(shù)有限公司;增強(qiáng)型化學(xué)發(fā)光(enhanced chemiluminescence,ECL)免疫印跡檢測(cè)試劑 江蘇新賽美生物科技有限公司;其他試劑均為分析純。
1.2 儀器與設(shè)備
1300系列A2型生物安全柜、梯度PCR儀、恒溫恒濕培養(yǎng)箱 美國Thermo Fisher Scientific公司;SpectraMax i3多功能酶標(biāo)儀 美國Molecular Devices公司;CFX 96 Touch實(shí)時(shí)熒光定量PCR儀 美國Bio-Rad公司;Gallios流式細(xì)胞儀 美國Beckman公司;DYCZ-24DN型電泳儀、DYCZ-40D轉(zhuǎn)印槽 北京六一儀器廠;ChemiScope 4300 pro化學(xué)發(fā)光成像系統(tǒng) 上海勤翔科學(xué)儀器有限公司。
1.3 方法
1.3.1 羊棲菜多酚的制備
參照Wang Chen[14]、李亞嫻[15]等的方法并略作修改。取干燥羊棲菜粉末,加入體積分?jǐn)?shù)60%乙醇溶液超聲輔助浸提(料液比1∶20(m/V)、50 ℃、60 min),抽濾后真空減壓蒸餾去除乙醇,獲羊棲菜粗多酚溶液,依次采用等體積石油醚、乙酸乙酯分級(jí)萃取,真空減壓蒸餾后凍干,獲得HFPs粉末。測(cè)得HFPs粉末總酚含量為(23.67±0.24)mg/g。利用無菌水配制成多酚母液,再以細(xì)胞培養(yǎng)液稀釋至不同濃度,用于后續(xù)實(shí)驗(yàn)。
1.3.2 細(xì)胞培養(yǎng)
完全培養(yǎng)基(含10%(體積分?jǐn)?shù),下同)胎牛血清和1%青鏈霉素雙抗DMEM培養(yǎng)基)培養(yǎng)RAW264.7細(xì)胞,37 ℃、5% CO2培養(yǎng),細(xì)胞融合度約80%時(shí)進(jìn)行傳代。
1.3.3 RAW264.7細(xì)胞活力測(cè)定
取對(duì)數(shù)生長期細(xì)胞,以3×105個(gè)/mL接種于96 孔板,每孔100 μL。待細(xì)胞貼壁后,更換為含不同質(zhì)量濃度(0(即對(duì)照組)、10、20、40、60、80、100、120、140、160、180、200、250、300、350 μg/mL)HFPs的新鮮完全培養(yǎng)基,培養(yǎng)24 h。棄去上清液,加入100 μL含0.5 mg/mL MTT的DMEM/F12無酚紅培養(yǎng)基,37 ℃避光孵育4 h。再次棄去上清液,每孔加入200 μL二甲基亞砜,37 ℃振蕩30 min,測(cè)定540 nm處OD540nm值。按公式(1)計(jì)算細(xì)胞活力。

1.3.4 RAW264.7細(xì)胞NO相對(duì)含量測(cè)定
參照1.3.3節(jié)方法進(jìn)行細(xì)胞接種,培養(yǎng)24 h細(xì)胞貼壁后,將細(xì)胞分為3 組:陰性對(duì)照組、LPS陽性對(duì)照組、HFPs+LPS組。陰性對(duì)照組加入含10%胎牛血清的DMEM培養(yǎng)基;LPS陽性對(duì)照組加入含LPS(1 μg/mL)和10%胎牛血清的DMEM培養(yǎng)基刺激24 h;HFPs+LPS組經(jīng)含不同質(zhì)量濃度(10、20、40、60、80、100、120、140、160、180、200、250、300、350 μg/mL)HFPs和10%胎牛血清的DMEM培養(yǎng)基培養(yǎng)24 h后,以含LPS(1 μg/mL)和10%胎牛血清的DMEM培養(yǎng)基刺激24 h。采用Griess法測(cè)定各組在540 nm處OD540nm值,按公式(2)計(jì)算NO相對(duì)含量。

1.3.5 RAW264.7細(xì)胞炎癥相關(guān)基因相對(duì)表達(dá)量的測(cè)定
取對(duì)數(shù)生長期細(xì)胞,以4×105個(gè)/mL接種于6 孔板,每孔2.5 mL。細(xì)胞分為3 組:陰性對(duì)照組,LPS陽性對(duì)照組、HFPs+LPS組。LPS陽性對(duì)照組在含10%胎牛血清的DMEM培養(yǎng)基中培養(yǎng)24 h后,以含LPS(1 μg/mL)和10%胎牛血清的DMEM培養(yǎng)基刺激3、6、12、24 h;HFPs+LPS組經(jīng)含不同質(zhì)量濃度(30、60、90、120 μg/mL)HFPs和10%胎牛血清的DMEM培養(yǎng)基培養(yǎng)24 h后,以含LPS(1 μg/mL)和10%胎牛血清的DMEM培養(yǎng)基刺激3、6、12、24 h;陰性對(duì)照組用含10%胎牛血清的DMEM培養(yǎng)基孵育24 h后吸棄培養(yǎng)液,用含不同質(zhì)量濃度(0、30、60、90、120 μg/mL)HFPs和10%胎牛血清的DMEM培養(yǎng)基培養(yǎng)24 h。培養(yǎng)結(jié)束后棄去培養(yǎng)液,用0.01 mol/L、pH 7.2(后同)的磷酸鹽緩沖液(phosphate buffered saline,PBS)清洗細(xì)胞2 次,每孔加入500 μL TRIzol試劑提取總RNA,并將RNA逆轉(zhuǎn)錄為cDNA。以cDNA為模板,采用SYBR Green嵌合熒光法與引物(表1)進(jìn)行實(shí)時(shí)熒光定量PCR。測(cè)定目的基因(IL-1β、IL-6、TNF-α、COX-2、iNOS)和內(nèi)參基因次黃嘌呤磷酸核糖基轉(zhuǎn)移酶1(hypoxanthine-guanine phosphoribosyl transferase,hprt1)Ct值,采用2-ΔΔCt法計(jì)算目的基因的相對(duì)表達(dá)量。

表1 實(shí)時(shí)熒光定量PCR引物序列Table 1 Primer sequences used for qPCR
1.3.6 RAW264.7細(xì)胞吞噬能力的測(cè)定
參照李煜等[16]的方法并略作修改。按照1.3.5節(jié)方法分組進(jìn)行藥物處理,陰性對(duì)照組為無LPS和HFPs處理,LPS組和HFPs+LPS組LPS刺激3 h后,去除培養(yǎng)基,加入1 mL熒光微球工作液(1×107個(gè)/mL)避光孵育2 h。收集細(xì)胞,1 000 r/min離心5 min,PBS洗2 次,加入1 mL PBS重懸細(xì)胞,經(jīng)75 μm尼龍膜過濾后立即采用流式細(xì)胞儀測(cè)定細(xì)胞熒光強(qiáng)度。以實(shí)驗(yàn)組與陰性對(duì)照組熒光強(qiáng)度的比值百分?jǐn)?shù)表征細(xì)胞吞噬能力。
1.3.7 MAPKs、NF-κB信號(hào)通路關(guān)鍵蛋白相對(duì)表達(dá)量測(cè)定
按照1.3.5節(jié)方法進(jìn)行分組和處理,陰性對(duì)照組為無LPS和HFPs處理,LPS組和HFPs+LPS組LPS刺激誘導(dǎo)24 h后,收集細(xì)胞。加入200 μL RIPA細(xì)胞裂解液于冰上裂解30 min,離心(12 000 r/min、4 ℃)10 min取上清液。采用BCA蛋白質(zhì)量濃度測(cè)定試劑盒測(cè)定各組蛋白濃度,用蛋白上樣緩沖液調(diào)整蛋白質(zhì)量濃度為2 mg/mL,95 ℃加熱10 min使蛋白變性然后進(jìn)行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳分離;濕轉(zhuǎn)到聚偏二氟乙烯膜;5%(質(zhì)量分?jǐn)?shù))脫脂奶粉(TBST溶液配制)封閉1 h;一抗4 ℃孵育12 h;二抗室溫孵育2 h;加入ECL試劑進(jìn)行顯色反應(yīng),測(cè)定蛋白條帶灰度值。以β-actin為內(nèi)參,目的蛋白包括iNOS、p65、p-p65、p38、p-p38、JNK和p-JNK,分別以p-p38/p38、p-JNK/JNK、p-p65/p65表示相應(yīng)蛋白磷酸化水平。
1.4 數(shù)據(jù)統(tǒng)計(jì)與分析
每個(gè)實(shí)驗(yàn)重復(fù)3 次,結(jié)果均以平均值±標(biāo)準(zhǔn)差表示,采用Prism 8.3軟件進(jìn)行單因素方差分析。采用t檢驗(yàn)進(jìn)行顯著性分析,P<0.05表示差異顯著。
2 結(jié)果與分析
2.1 HFPs對(duì)RAW264.7細(xì)胞活力的影響
如圖1所示,與對(duì)照組相比,當(dāng)HFPs質(zhì)量濃度不高于160 μg/mL時(shí),其對(duì)細(xì)胞活力無明顯作用;進(jìn)一步提高多酚質(zhì)量濃度后,HFPs顯著抑制細(xì)胞增殖(P<0.05),并呈劑量依賴性,當(dāng)HFPs質(zhì)量濃度達(dá)到350 μg/mL,細(xì)胞活力顯著降為對(duì)照組的50%(P<0.05)。

圖1 HFPs對(duì)RAW264.7細(xì)胞活力的影響Fig. 1 Effects of HFPs on the viability of RAW264.7 cells
2.2 HFPs對(duì)RAWA264.7細(xì)胞NO相對(duì)含量的影響
巨噬細(xì)胞經(jīng)LPS刺激后,可產(chǎn)生多種炎性介質(zhì)包括促炎細(xì)胞因子、前列腺素和NO等。如圖2所示,與陰性對(duì)照組相比,細(xì)胞經(jīng)LPS刺激24 h后,其NO相對(duì)含量顯著升高(P<0.05)。與陽性對(duì)照相比,采用HFPs預(yù)處理可劑量依賴性緩解LPS刺激導(dǎo)致的NO產(chǎn)生,且當(dāng)HFPs質(zhì)量濃度為100 μg/mL以上時(shí),抑制效果顯著(P<0.05)。
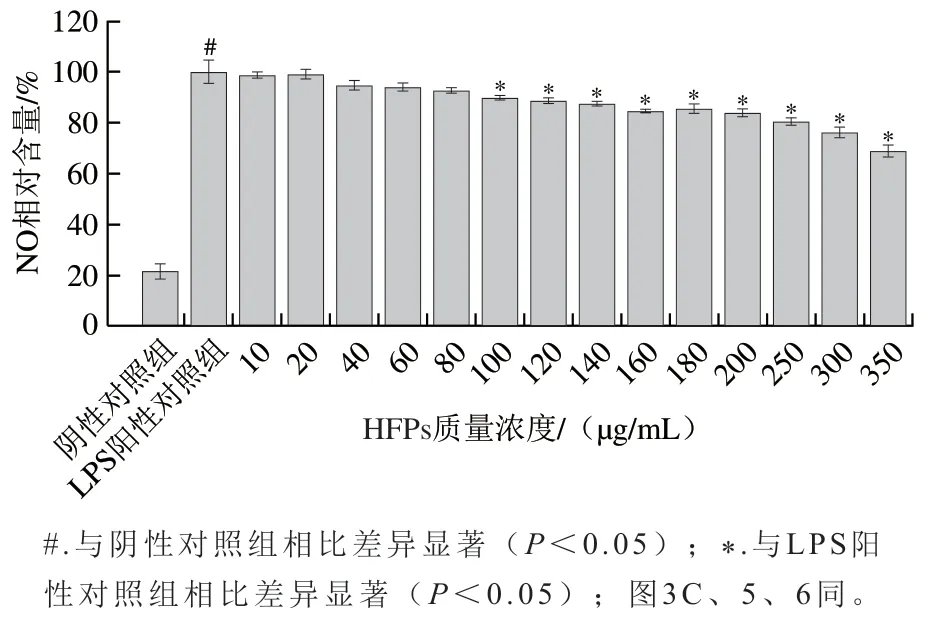
圖2 HFPs對(duì)RAW264.7細(xì)胞NO生成量的影響Fig. 2 Effect of HFPs on NO production in RAW264.7 cells
2.3 HFPs對(duì)RAW264.7細(xì)胞iNOS mRNA和蛋白表達(dá)的影響
如圖3A所示,LPS顯著刺激細(xì)胞iNOS基因轉(zhuǎn)錄(P<0.05),且iNOSmRNA表達(dá)水平隨LPS誘導(dǎo)時(shí)間延長而增加。HFPs預(yù)處理24 h后再經(jīng)LPS誘導(dǎo)細(xì)胞,HFPs對(duì)iNOSmRNA表達(dá)水平的影響與LPS刺激時(shí)間有關(guān),短期LPS刺激(0~12 h)后HFPs對(duì)iNOSmRNA表達(dá)水平無明顯作用,而當(dāng)LPS誘導(dǎo)時(shí)間延長至24 h時(shí),HFPs(60~120 μg/mL)處理顯著降低了iNOSmRNA相對(duì)表達(dá)量(P<0.05)。蛋白免疫印跡實(shí)驗(yàn)進(jìn)一步印證了該結(jié)果,60~120 μg/mL HFPs預(yù)處理顯著降低了LPS誘導(dǎo)24 h巨噬細(xì)胞iNOS蛋白水平(P<0.05)(圖3B、C)。
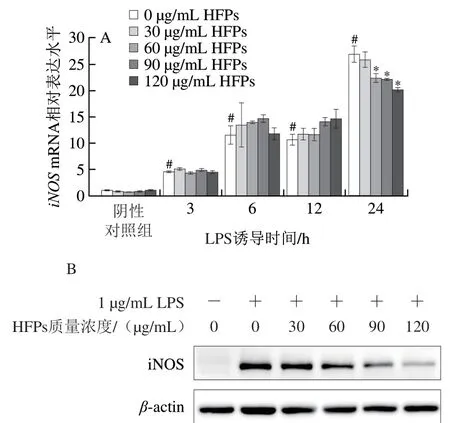

圖3 HFPs對(duì)RAW264.7細(xì)胞iNOS mRNA和蛋白表達(dá)的影響Fig. 3 Effect of HFPs on the expression of iNOS mRNA and protein in RAW264.7 cells
2.4 HFPs對(duì)RAW264.7細(xì)胞炎癥基因相對(duì)表達(dá)量的影響
采用實(shí)時(shí)熒光定量PCR測(cè)定HFPs對(duì)LPS誘導(dǎo)的巨噬細(xì)胞中重要促炎細(xì)胞因子IL-1β、IL-6和TNF-α基因相對(duì)表達(dá)量,以及炎癥相關(guān)酶COX-2基因相對(duì)表達(dá)量的影響。如圖4所示,LPS刺激不同時(shí)間后,所有炎癥介質(zhì)的mRNA表達(dá)水平均顯著提高(P<0.05)。HFPs對(duì)促炎細(xì)胞因子的抑制作用與其劑量、LPS誘導(dǎo)時(shí)間及細(xì)胞因子種類相關(guān)。與LPS組相比,靜息狀態(tài)細(xì)胞經(jīng)HFPs處理再經(jīng)LPS誘導(dǎo)12、24 h后,IL-1β和IL-6的基因相對(duì)表達(dá)量顯著下降;HFPs顯著抑制LPS誘導(dǎo)3 h后TNF-α基因的相對(duì)表達(dá),而在LPS誘導(dǎo)6~24 h的抑制作用總體不明顯。在LPS誘導(dǎo)24 h,較低劑量HFPs(30、60 μg/mL)預(yù)處理對(duì)COX-2基因表達(dá)具有顯著抑制作用,但提高劑量后該抑制效果消失,甚至出現(xiàn)反向促進(jìn)作用,120 μg/mL HFPs預(yù)處理促使COX-2相對(duì)表達(dá)量較LPS組顯著上升(P<0.05)。
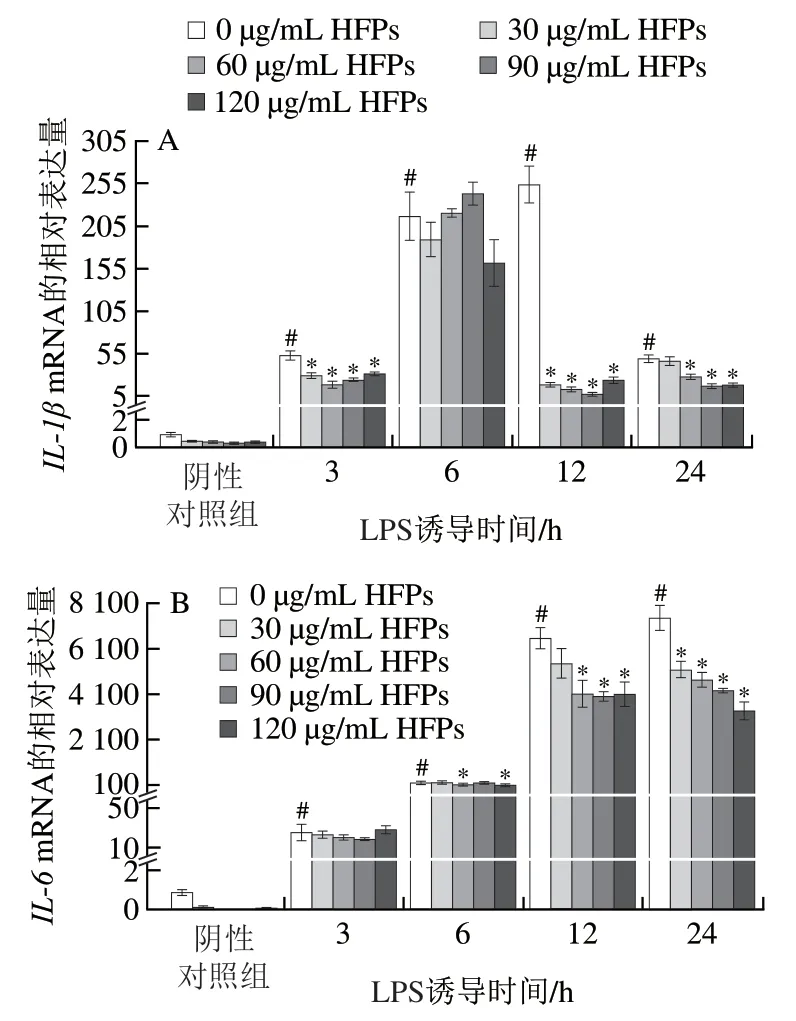
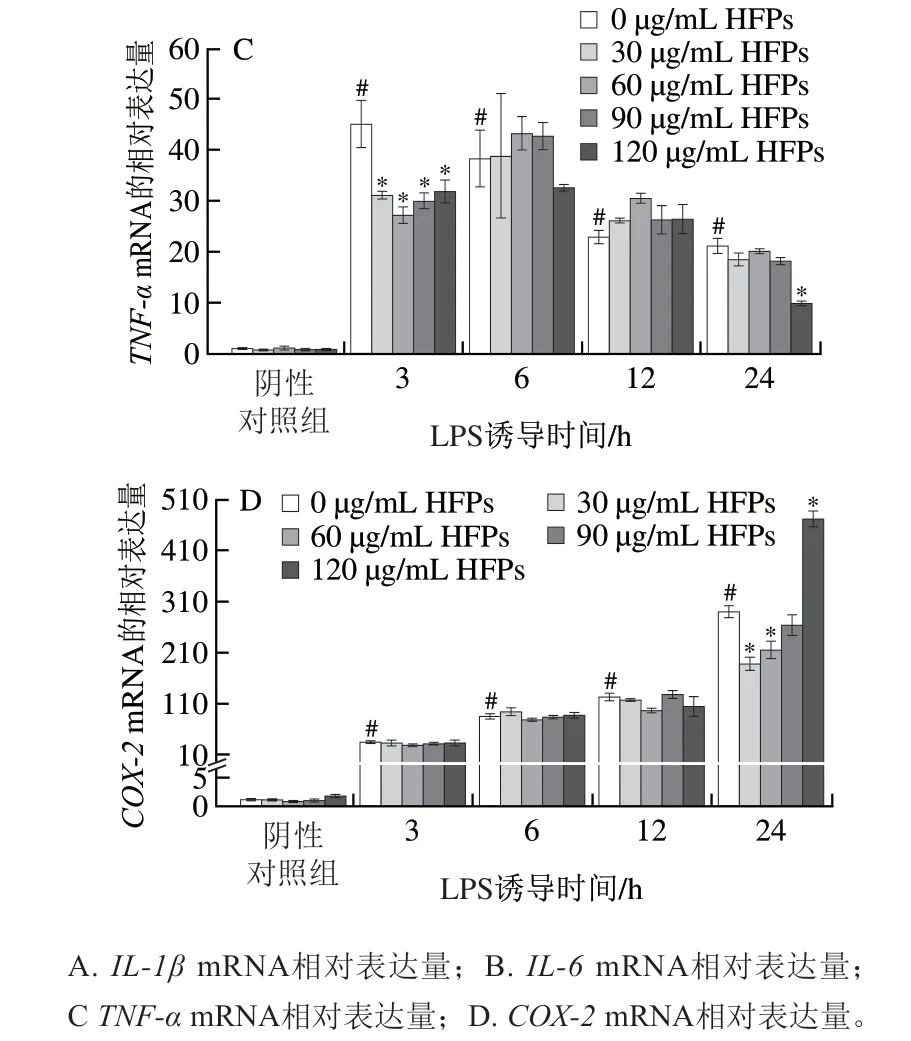
圖4 HFPs對(duì)RAW264.7細(xì)胞炎癥介質(zhì)基因相對(duì)表達(dá)量的影響Fig. 4 Effect of HFPs on gene expression of inflammatory mediators in RAW264.7 cells
2.5 HFPs對(duì)RAW264.7細(xì)胞吞噬能力的影響
如圖5所示,與陰性對(duì)照組相比,LPS刺激3 h顯著促進(jìn)了細(xì)胞吞噬熒光微球的能力(P<0.05)。經(jīng)60、90、120 μg/mL HFPs預(yù)處理后,LPS誘導(dǎo)的小鼠巨噬細(xì)胞吞噬能力受到顯著抑制(P<0.05)。
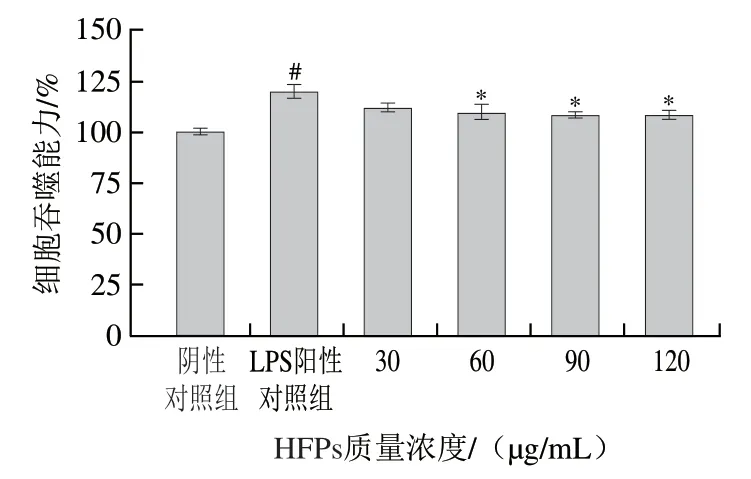
圖5 HFPs對(duì)RAW264.7細(xì)胞吞噬能力的影響Fig. 5 Effect of HFPs on phagocytic capacity of RAW264.7 cells
2.6 HFPs對(duì)RAW264.7細(xì)胞MAPKs、NF-κB信號(hào)通路關(guān)鍵蛋白磷酸化水平的影響
采用蛋白免疫印跡法測(cè)定巨噬細(xì)胞中MAPKs和NF-κB信號(hào)通路關(guān)鍵蛋白的磷酸化水平。如圖6所示,與陰性對(duì)照組相比,LPS刺激3 h后細(xì)胞中p38、JNK和p65蛋白的磷酸化水平顯著升高(P<0.05)。90、120 μg/mL HFPs預(yù)處理對(duì)LPS誘導(dǎo)的巨噬細(xì)胞中p38和p65的磷酸化產(chǎn)生顯著抑制作用(P<0.05),且呈一定的劑量依賴性,但對(duì)JNK的磷酸化沒有顯著影響,表明HFPs的抗炎作用可能與其靶向作用于p38 MAPK和NF-κB p65有關(guān)。

圖6 HFPs對(duì)RAW264.7細(xì)胞MAPKs和NF-κB信號(hào)通路關(guān)鍵蛋白磷酸化水平的影響Fig. 6 Effect of HFPs on the expression of key proteins involved in MAPKs and NF-κB signaling pathways in RAW264.7 cells
3 討 論
巨噬細(xì)胞是機(jī)體免疫應(yīng)答的第一道防線,具有識(shí)別、吞噬、清除細(xì)菌及外來異物等功能[17]。然而,當(dāng)巨噬細(xì)胞被異常激活,可分泌大量促炎細(xì)胞因子進(jìn)而引發(fā)炎癥風(fēng)暴,導(dǎo)致機(jī)體組織損傷,故其靜息-激活的動(dòng)態(tài)平衡在維持免疫系統(tǒng)穩(wěn)態(tài)中起重要作用[18]。針對(duì)過度免疫反應(yīng),一般可通過阻斷或抑制炎癥相關(guān)因子的表達(dá)達(dá)到緩解炎癥及其引起的機(jī)體損傷。
細(xì)胞增殖能力是機(jī)體的重要生命特征和體現(xiàn)細(xì)胞活力的重要指標(biāo)。MTT法是一種簡單、快捷、無放射性污染測(cè)定細(xì)胞活力的方法[19],在本研究中用于評(píng)估HFPs對(duì)小鼠巨噬細(xì)胞的潛在細(xì)胞毒性。結(jié)果顯示,HFPs的最大安全質(zhì)量濃度為160 μg/mL,即不高于該劑量時(shí)HFPs對(duì)RAW264.7細(xì)胞活力無影響。
LPS是革蘭氏陰性菌細(xì)胞壁外壁的組成成分,又名內(nèi)毒素,是引起機(jī)體炎癥損傷的重要誘因,可激活巨噬細(xì)胞產(chǎn)生NO、iNOS、COX-2等多種炎癥介質(zhì)和IL-6、IL-1β、TNF-α等促炎細(xì)胞因子[20-22]。IL-1β大量產(chǎn)生于炎癥初期,其局部激活是介導(dǎo)促炎反應(yīng)的中心環(huán)節(jié),可導(dǎo)致繼發(fā)性炎癥介質(zhì)的產(chǎn)生[20];TNF-α具有多效性,能增加血管內(nèi)皮細(xì)胞通透性,激活中性粒細(xì)胞和淋巴細(xì)胞,促進(jìn)其他細(xì)胞因子的合成和釋放[23];IL-6作為重要促炎細(xì)胞因子,可誘導(dǎo)B細(xì)胞分化和產(chǎn)生抗體,誘導(dǎo)T細(xì)胞活化、增殖及分化,參與機(jī)體免疫應(yīng)答[24]。研究表明,這些細(xì)胞因子及其他炎性介質(zhì)的大量釋放,與炎癥相關(guān)性疾病的發(fā)生和發(fā)展密切相關(guān)[25-26]。本研究結(jié)果表明,RAW264.7細(xì)胞經(jīng)LPS刺激后,其炎癥介質(zhì)基因(IL-1β、IL-6、TNF-α、COX-2、iNOS)表達(dá)顯著增加,而這一現(xiàn)象在HFPs預(yù)處理后得到緩解,并呈劑量和時(shí)間依賴性。Kim等發(fā)現(xiàn)間苯三酚可降低LPS刺激的RAW264.7細(xì)胞中TNF-α、IL-1β和IL-6基因表達(dá)水平,抑制巨噬細(xì)胞炎癥反應(yīng)[27]。Kang等則研究發(fā)現(xiàn)褐藻Eisenia bicyclis提取物抑制LPS激活的HepG2細(xì)胞中iNOS和COX-2的表達(dá)[28]。結(jié)合本研究結(jié)果,表明HFPs及其他褐藻多酚可通過調(diào)節(jié)多種炎性介質(zhì)的基因轉(zhuǎn)錄水平來抑制炎癥的發(fā)生。
巨噬細(xì)胞的吞噬功能在機(jī)體防御病原體感染中發(fā)揮重要作用。需要注意的是,過多的免疫細(xì)胞(包括巨噬細(xì)胞)被激活,將增加炎癥風(fēng)暴風(fēng)險(xiǎn)[18]。流式細(xì)胞術(shù)結(jié)合熒光標(biāo)記廣泛應(yīng)用于巨噬細(xì)胞吞噬功能評(píng)估[16]。本研究利用該技術(shù)檢測(cè)了HFPs對(duì)RAW264.7細(xì)胞吞噬功能的影響。結(jié)果顯示,RAW264.7細(xì)胞經(jīng)LPS刺激后,其吞噬熒光微球的能力顯著提升,這與Wen Li等[29]的發(fā)現(xiàn)一致。采用HFPs預(yù)處理后,LPS誘導(dǎo)的細(xì)胞吞噬能力受到抑制,表明HFPs通過調(diào)節(jié)細(xì)胞吞噬功能緩解了炎癥反應(yīng)。
NF-κB是具有多種生物活性的核轉(zhuǎn)錄因子,能通過快速調(diào)節(jié)細(xì)胞內(nèi)基因的表達(dá),調(diào)控細(xì)胞增殖、凋亡、突變和炎癥反應(yīng)等生理病理過程[30]。細(xì)胞經(jīng)內(nèi)毒素刺激可誘發(fā)信號(hào)級(jí)聯(lián)反應(yīng),激活NF-κB并核轉(zhuǎn)位,啟動(dòng)下游基因的轉(zhuǎn)錄與蛋白的表達(dá),促進(jìn)炎癥相關(guān)因子的合成與分泌[18]。Yoon等發(fā)現(xiàn)馬尾藻(Sargassum micracanthum)提取物通過NF-κB信號(hào)通路抑制LPS誘導(dǎo)RAW264.7細(xì)胞中iNOS、COX-2、TNF-α、IL-1β、IL-6等mRNA水平的上調(diào)[31]。Yayeh等研究表明,在LPS誘導(dǎo)的RAW264.7細(xì)胞中,褐藻Eisenia bicyclis多酚提取物通過抑制NF-κB p65及其上游激酶的磷酸化降低NF-κB的轉(zhuǎn)錄活性[32]。除NF-κB外,MAPK也是巨噬細(xì)胞炎癥信號(hào)轉(zhuǎn)導(dǎo)中的重要因子,其磷酸化可進(jìn)一步導(dǎo)致NF-κB的激活[33]。Jung等[34-35]研究發(fā)現(xiàn),苷苔(Ecklonia cava)提取物能夠降低LPS誘導(dǎo)的小鼠BV2小膠質(zhì)細(xì)胞中NF-κB的核轉(zhuǎn)位及其DNA結(jié)合能力,而這一現(xiàn)象與抑制MAPKs的激活有關(guān)。目前,NF-κB/MAPKs信號(hào)通路的激活水平,即相關(guān)蛋白的表達(dá)和磷酸化水平,被廣泛應(yīng)用于評(píng)估和篩選抗炎活性物質(zhì)。本研究發(fā)現(xiàn),LPS刺激可顯著促進(jìn)關(guān)鍵蛋白的磷酸化(p-p65、p-p38、p-JNK),而HFPs預(yù)處理可劑量依賴性抑制p-p65和p-p38的表達(dá),表明HFPs的抗炎活性與調(diào)控p38 MAPK和NF-κB p65信號(hào)通路有關(guān)。不同抗炎藥物的抗炎機(jī)制不同,但主要通過阻斷或遏制炎癥相關(guān)通路,或直接中和機(jī)體中促炎介質(zhì)以達(dá)到抗炎效果[36]。本實(shí)驗(yàn)結(jié)果表明,HFPs可選擇性抑制LPS激活的p38/NF-κB信號(hào)通路,減少了IL-1β、IL-6、TNF-α、iNOS和COX-2等的表達(dá),從而緩解炎癥。
綜上所述,HFPs可通過抑制NF-κB/MAPKs信號(hào)通路蛋白p65和p38的磷酸化水平,減少促炎細(xì)胞因子和其他炎性介質(zhì)的合成與分泌,降低巨噬細(xì)胞吞噬作用,從而發(fā)揮抗炎活性。值得注意的是,許多報(bào)道證實(shí)了體內(nèi)外試驗(yàn)結(jié)果具有良好的相關(guān)性[37],但生物體內(nèi)情況復(fù)雜,化合物在體內(nèi)外的作用效果可能出現(xiàn)差異,如多酚進(jìn)入體內(nèi)后可能會(huì)被胃液分解成小分子酚類物質(zhì),導(dǎo)致效果減弱[38]。因此,在體外實(shí)驗(yàn)的基礎(chǔ)上還需結(jié)合體內(nèi)實(shí)驗(yàn),以確定HFPs在完整生物體中產(chǎn)生的抗炎作用,為其開發(fā)新的抗炎藥物提供重要依據(jù)。
