雅魯藏布江下游弧唇裂腹魚的年齡結構與生長特性
2022-12-21 07:24:22張富斌胡華明曹文宣林鵬程
水生生物學報 2022年12期
關鍵詞:生長
王 健 張富斌 胡華明 鞏 政 曹文宣 林鵬程
(1.中國科學院水生生物研究所, 武漢 430072; 2.水利部長江治理與保護重點實驗室, 長江設計集團有限公司, 武漢 430072;3.流域水安全保障湖北省重點實驗室, 長江勘測規(guī)劃設計研究有限責任公司, 武漢 430072; 4.西華師范大學環(huán)境科學與工程學院, 南充 637009; 5.棗莊學院生命科學學院, 棗莊 277160)
裂腹魚亞科魚類(Schizothoracinae fishes, 以下簡稱裂腹魚類)主要分布于青藏高原及其鄰近地區(qū),我國有11屬97種或亞種[1,2]。裂腹魚類具有生長緩慢、壽命較高、性成熟晚、繁殖力低等生物學特征, 使其對高原河流水資源的過度開發(fā)和環(huán)境退化較為敏感[2]。雅魯藏布江流域橫貫西藏, 棲息有多種裂腹魚類。近年來, 受生境退化、過度捕撈、水電梯級開發(fā)和生物入侵等因素影響, 雅魯藏布江流域以尖裸鯉(Oxygymnocypris stewarti)、拉薩裂腹魚(Schizothorax waltoni)、巨須裂腹魚(S.macropogon)和雙須葉須魚(Ptychobarbus dipogon)等為代表的裂腹魚類資源急劇下降, 高原魚類的資源保護問題日益突出[2]。因此, 需要對裂腹魚類的自然種群特征及變動進行系統研究。
雅魯藏布江下游地處西藏東南部, 主要位于墨脫縣境內, 是印度洋濕暖氣流進入高原內部的主要通道。區(qū)域內雨水豐沛, 河谷深切, 河道比降大, 水流湍急。加之兩岸階地堆積松散, 泥石流、塌方、山體滑坡等自然災害時有發(fā)生[3]。隨著經濟社會發(fā)展和國家“十四五”規(guī)劃的實施, 雅魯藏布江下游的水域生態(tài)環(huán)境將受到一定的干擾。作為雅魯藏布江下游的優(yōu)勢物種, 弧唇裂腹魚(Schizothorax curvilabiatus)對于維持下游河流生態(tài)系統的結構和功能具有重要的生態(tài)價值[1]。但弧唇裂腹魚的研究僅涉及形態(tài)、分類、棲息習性和攝食等方面的簡單描述, 缺少詳細的基礎生物學數據[4,5]。
魚類生長是其生活史中重要的生物學過程。年齡與生長特征是研究魚類生物學和種群動態(tài)的基礎和前提, 獲取準確的年齡數據是評估魚類生長率、種群補充量和死亡率的關鍵[6—8]。在此背景下對弧唇裂腹魚的年齡和生長特征進行分析, 以揭示物種的生長特性和種群生長潛力, 為變化環(huán)境下雅魯藏布江下游魚類資源的保護提供科學依據。
1 材料與方法
1.1 樣本采集和年齡材料獲取
2015年12月至2016年11月逐月在雅魯藏布江下游墨脫段開展魚類采集工作(圖1), 獲取弧唇裂腹魚928尾。采樣網具為定置刺網(網目4—8 cm)、撒網(網目6 cm)和地籠(網目1 cm)等(調查采樣均在漁政主管部門備案, 禁用網具取得捕撈許可)。樣本獲取后測定體長、體重、內臟重和性腺重(長度精確至l mm, 重量精確至0.1 g)。根據性腺發(fā)育判斷樣本性別。取出微耳石、鰓蓋骨和脊椎骨, 耳石用清水洗凈后放入離心管中保存, 脊椎骨和鰓蓋骨置于自封袋中冰凍保存。
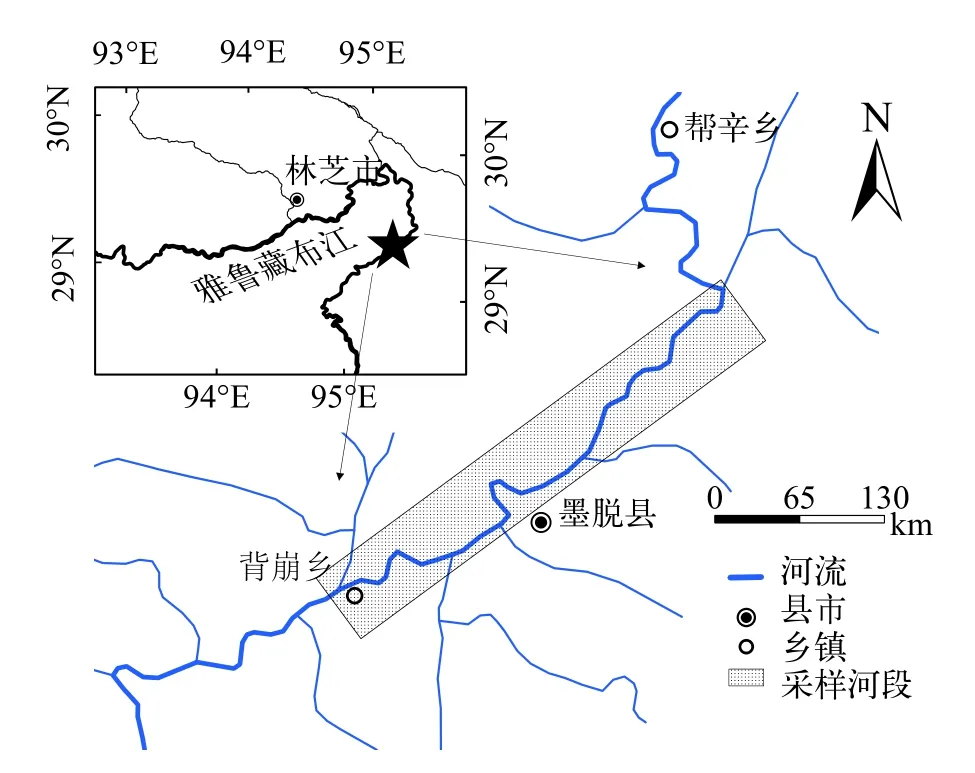
圖1 雅魯藏布江下游弧唇裂腹魚的采樣區(qū)域Fig.1 Sampling area of S.curvilabiatus in the lower reach of Yarlung Zangbo River
1.2 年齡材料處理
耳石用指甲油將微耳石包埋并固定于載波片一邊, 使其凸面朝上, 隨后用800—2500#的水磨砂紙依次打磨, 再用拋光紙拋光。打磨時, 不時地移至顯微鏡下觀察耳石輪紋。當看到耳石中心較為清楚時, 利用丙酮溶解指甲油, 將耳石翻面包埋, 用相同的方法打磨和拋光直至耳石中心和外周輪紋都較清晰為止[9]。
脊椎骨將脊椎骨置于沸水中加熱約10min,去除附著的軟組織后置于1% 的H2O2中浸泡24h。晾干后用剪刀將脊椎骨從中間剖開, 置于解剖鏡下觀察(入射光), 并用二甲苯透明處理。
鰓蓋骨將鰓蓋骨置于沸水中加熱約1min,去除附著的軟組織, 再將其置于1%的 H2O2中浸泡24h后晾干, 最后在解剖鏡下觀察(透射光)。
1.3 年齡材料鑒定和比較
年齡的確認和計數方法參照殷名稱等[10,11]。同一鑒定者對弧唇裂腹魚的耳石磨片, 脊椎骨和鰓蓋骨進行兩次獨立讀數, 兩次讀數間隔3周以上。并對耳石、脊椎骨和鰓蓋骨的年輪清晰度進行5等級評分: 1為非常好, 2為好, 3為一般, 4為很差, 5為難以辨認[12](表1)。

表1 年齡材料輪紋清晰度等級劃分表Tab.1 Readability level of annuli on different calcified structures
1.4 數據分析
體長體重關系通過冪函數(W=aSLb)對弧唇裂腹魚的體長體重關系進行回歸分析, 式中W為體重(g),SL為體長(mm), a, b為常數, a為生長的條件因子, b為冪指數系數。同時將體長體重數據進行對數化處理, 采用協方差分析(ANCOVA)來檢驗不同性別體長體重的關系是否具有顯著性差異[13]。并對異速生長指數(b)和3的差異性進行t檢驗[14],以此推測弧唇裂腹魚是否為勻速生長類型魚類。
生長方程統計雌雄個體不同年齡組實測體長的平均值和范圍。采用von Bertalanffy生長方程Lt=L∞[1-ek(t-t0)]擬合弧唇裂腹魚的生長特征,式中Lt為t齡時的實測體長,L∞為漸進體長,k為生長曲線的平均曲率;t0為理論上體長和體重等于零時的年齡。表觀生長指數(?)被用于同屬魚類生長性能的比較,計算公式如下:?=1gk+2lgL∞[15]。數據用平均數±標準差表示, 統計顯著性為α=0.05。統計分析采用SPSS 16.0和Origin Pro 2016, 圖像分析和處理采用Photoshop 2020。
2 結果
2.1 年齡材料特征
隨機選取了部分樣品的三種年齡材料進行加工處理, 最終對耳石、脊椎骨和鰓蓋骨都成功鑒定年齡的有236尾(體長112—508 mm, 鑒定結果見表2)。三種年齡材料上均存在年輪, 但年輪特征各異。
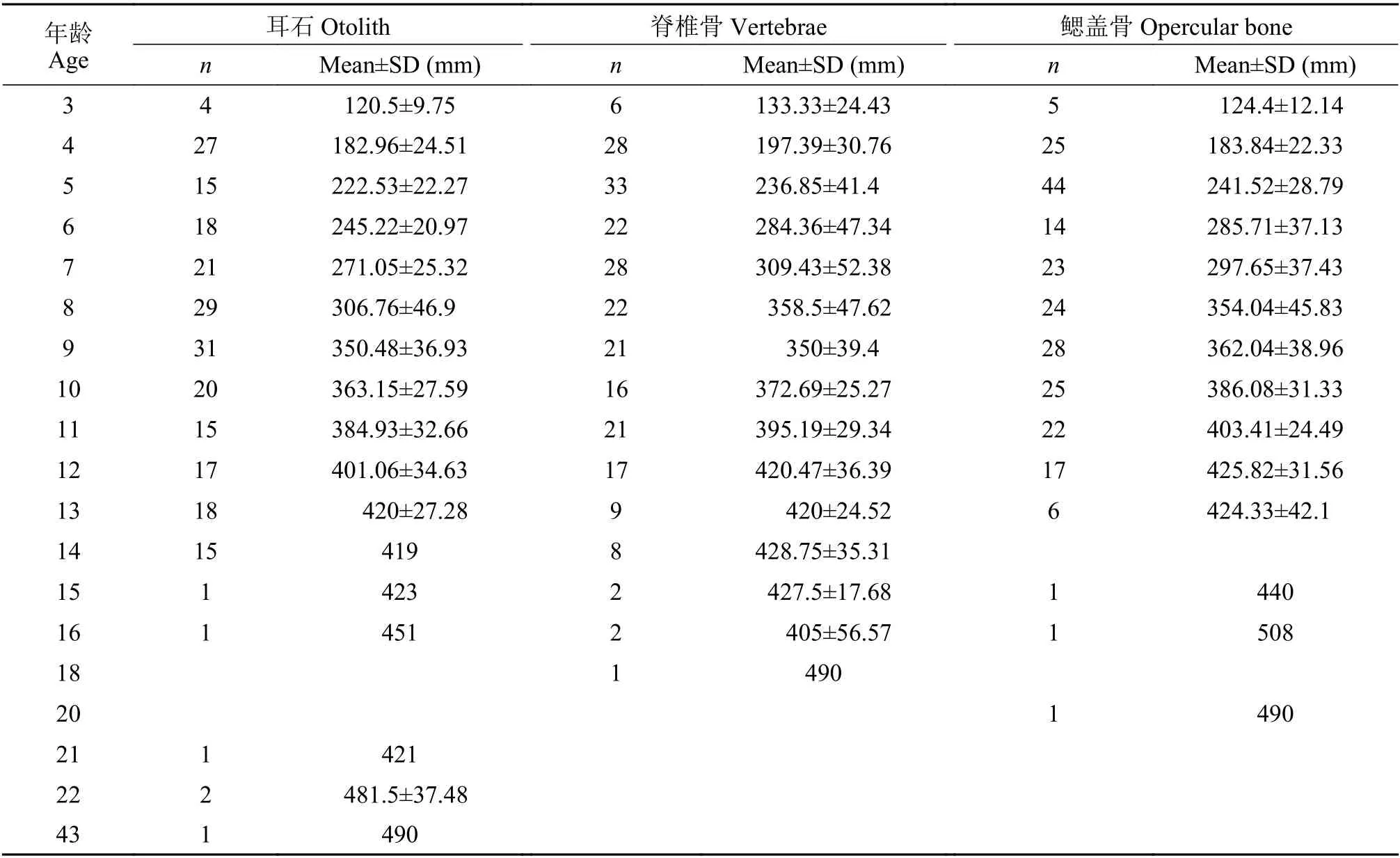
表2 弧唇裂腹魚各個年齡組的樣本數和體長Tab.2 Number and standard length of S.curvilabiatus at different ages
微耳石有清晰輪紋。經過打磨后的微耳石中心有個核區(qū), 在透射光下呈現出明顯的由寬帶和窄帶(或明帶和暗帶)組成的輪紋。年輪間距在耳石上呈規(guī)律性分布, 靠近核區(qū)的輪紋的間距較寬大, 越往邊緣延伸輪紋間距越來越窄(圖2)。
脊椎骨為雙凹型, 在解剖鏡下可觀察到明暗相間的輪紋, 輪紋呈同心橢圓排列。椎骨中央具一小孔, 小孔周圍薄而透明, 輪紋少且細, 較難確認首輪的位置(圖2)。
弧唇裂腹魚的鰓蓋骨呈不規(guī)則扇形, 鰓蓋骨上的年帶由乳白色寬帶和暗黑色窄帶組成。窄帶和寬帶之間有明顯的界限, 這一界限為年輪標志(圖2)。

圖2 弧唇裂腹魚不同年齡材料輪紋特征 (圓點表示年輪)Fig.2 Annuli characteristics of different calcified structures of S.curvilabiatus (dots show annuli)
耳石、脊椎骨和鰓蓋骨的清晰度評分見表3。耳石輪紋最為清晰, 1、2等級的耳石占總數72.03%;脊椎骨次之, 1、2等級的耳石的脊椎骨占總數58.90%; 鰓蓋骨上年輪清晰度最差, 1、2等級的鰓蓋骨只有47.88%。綜上可見, 耳石對弧唇裂腹魚的年齡鑒定效果最佳, 脊椎骨次之, 鰓蓋骨最差。
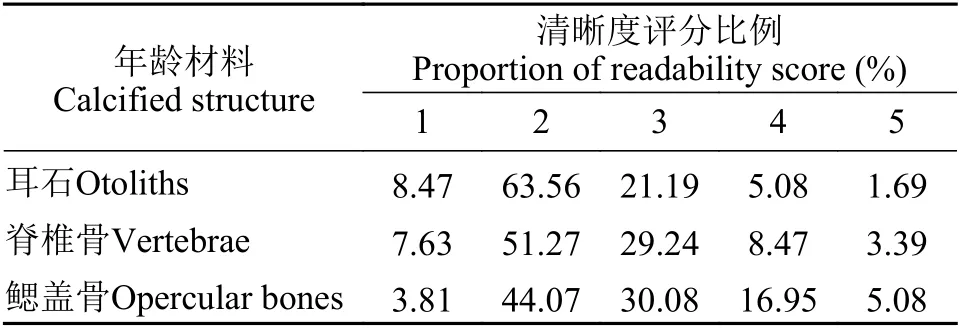
表3 弧唇裂腹魚不同年齡材料的清晰度評分Tab.3 Distribution of readability scores for different calcified structures of S.curvilabiatus (n=236)
從圖3可知, 耳石磨片兩次讀數一直保持較高的吻合率, 鰓蓋骨差異性較大。在 9齡之前, 脊椎骨兩次年齡讀數能保持很好的一致性, 但隨年齡的遞增, 鑒定結果出現差異。整體來看, 弧唇裂腹魚耳石精確性都要優(yōu)于脊椎骨和鰓蓋骨。
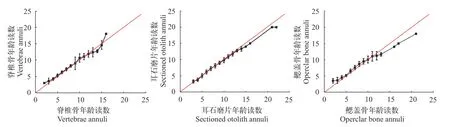
圖3 同一觀察者兩次年齡鑒定結果偏差圖Fig.3 Age bias plots for pairwise age estimates from two independent readings
由表4可以看出, 三種年齡材料所鑒定的平均年齡存在顯著性差異(P<0.05)。用耳石作為年齡材料所鑒定的平均年齡最高(8.87齡), 顯著高于脊椎骨(7.62齡)和鰓蓋骨(7.81齡)所鑒定的平均年齡(P<0.05)。10齡以下和10齡以上樣本分開統計時, 在1—10齡樣本中, 耳石鑒定的平均年齡(7.01齡), 顯著高于鰓蓋骨(6.60齡)和脊椎骨(6.37齡)所鑒定的平均年齡 (P<0.05)。大于10齡時, 脊椎骨(11.63齡)和鰓蓋骨(12.0齡)所鑒定的平均年齡較為接近, 但都顯著低于耳石所鑒定的平均年齡(13.2齡;P<0.05)。
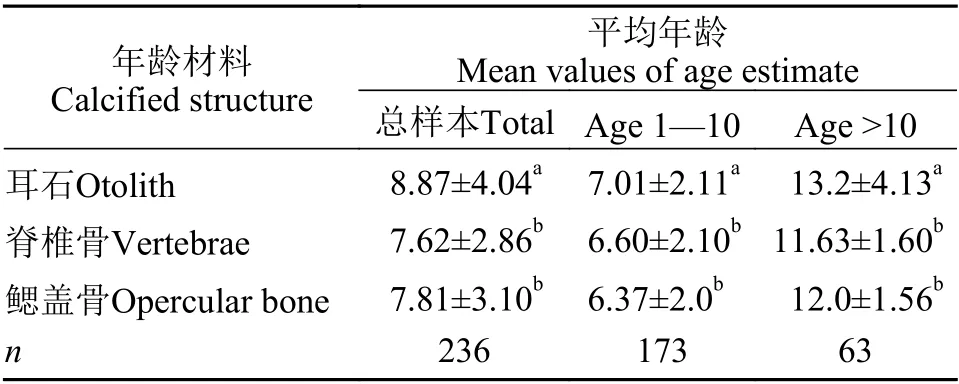
表4 弧唇裂腹魚不同年齡材料所鑒定的平均年齡Tab.4 Mean values of age estimates from different calcified structures of S.curvilabiatus
2.2 體長分布
弧唇裂腹魚的體長為48—508 mm, 均值為(227.4±111.4)mm。65.30%的個體體長集中在100—300 mm (圖4)。在 928尾樣本中, 雌性395尾,體長為85—508 mm; 雄性474尾, 體長為81—455 mm;性別未辨的個體59尾, 體長為48—170 mm。統計分析表明, 雌魚和雄魚的體長分布存在顯著性差異(Kolmogorov-SmirnovZ檢驗,Z=2.397,P<0.05)。
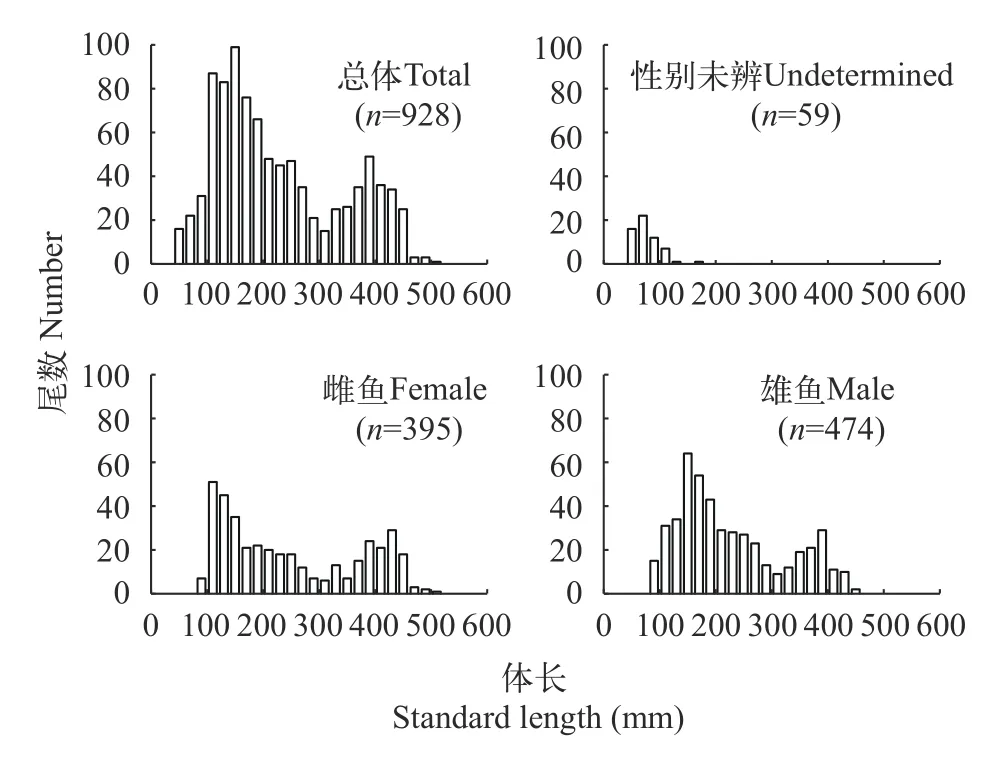
圖4 弧唇裂腹魚體長分布圖Fig.4 Frequency distribution of the standard length groups of S.curvilabiatus
2.3 年齡結構
在928尾樣本中, 因極少數耳石本身就不具備輪紋或者耳石在打磨處理過程中有磨損, 故成功鑒定微耳石年齡的樣本為916尾。其中, 雌魚年齡組樣本量為389尾,年齡為2—43齡; 雄魚年齡組樣本量為470尾,年齡為2—23齡, 性別未辨?zhèn)€體的年齡組樣本量為57尾,年齡為1—4齡。雌雄個體大于15齡的高齡組均少見,年齡分布主要集中在15齡以下。優(yōu)勢年齡組為3—5齡, 占全部樣本的44%(圖5)。不同性別不同年齡組的樣本數和平均體長見表5。
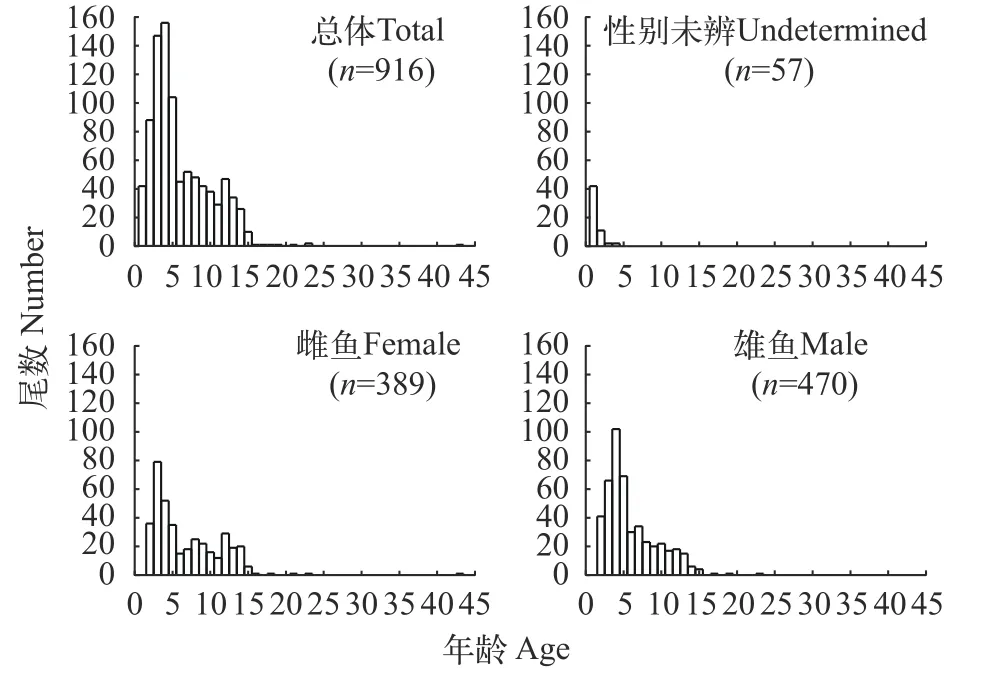
圖5 弧唇裂腹魚年齡組成圖Fig.5 Frequency distribution of estimated age groups of S.curvilabiatus
2.4 生長特征
體長體重關系將性別未辨?zhèn)€體、雌性群體、雄性群體和種群總體的體長和體重數據分別進行擬合(各方程擬合時的樣本量為各群體采集到的樣本數量; 圖6), 結果表明弧唇裂腹魚體長體重關系符合冪函數公式。
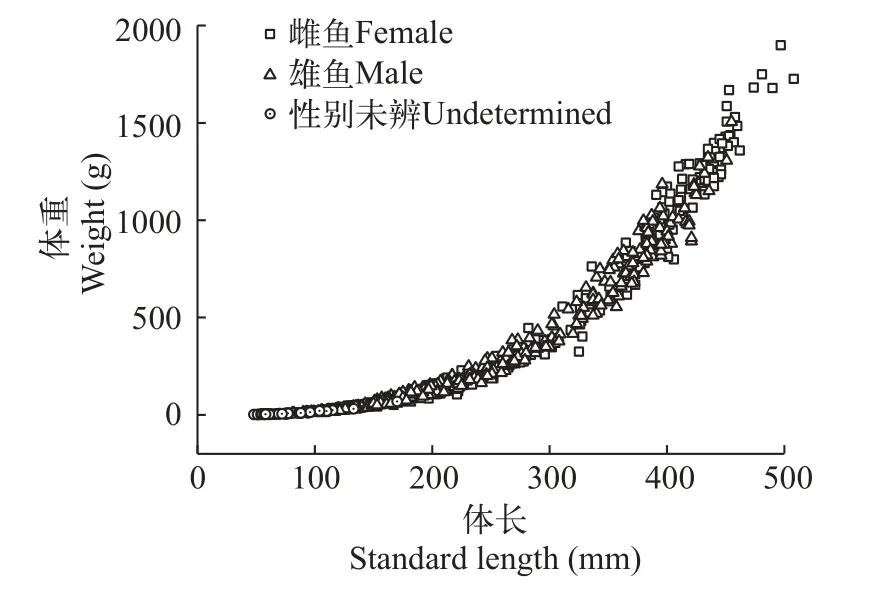
圖6 弧唇裂腹魚體長體重關系圖Fig.6 Length-weight relationship of S.curvilabiatus
性別未辨?zhèn)€體:W=3.122×10–5SL2.855,R2=0.983,n=59;
雌性群體:W=2.500×10–5SL2.914,R2=0.993,n=395;
雄性群體:W= 2.114×10–5SL2.944,R2=0.990,n=474;
種群總體:W=2.279×10–5SL2.931,R2=0.993,n=928。
弧唇裂腹魚幼魚的b值小于3, 為異速生長階段,表明此階段體長生長快于體重生長。經協方差分析, 顯示弧唇裂腹魚雌魚和雄魚間的體長體重關系差異并不顯著(F=0.833,P>0.05), 故將其合并為種群總體的冪函數方程。經t檢驗, 表明種群總體b值與3差異顯著(t=8.788,P<0.05), 表明弧唇裂腹魚為異速生長魚類。
生長方程經檢驗, 2—4齡的雌魚和雄魚,其同年齡組平均體長沒有顯著性差異(two-sample,P>0.05)。von Bertalanffy生長方程擬合時, 將性別未辨?zhèn)€體中2—4齡的樣本數據(n=15)分別納入雌魚(n=389)和雄魚(n=470)的年齡數據中, 以獲取更為準確的t0值[12](表5)。根據實測體長擬合的體長生長方程如下:

表5 弧唇裂腹魚不同年齡組的樣本數和平均體長Tab.5 Number of specimens and range of standard length at age of S.curvilabiatus
雌魚:Lt=590.2[1–e–0.096(t–0.282)](R2=0.932,n=404)
雄魚:Lt=575.3[1–e–0.090(t+0.011)](R2=0.918,n=485)
將體長體重關系式代入體長生長方程, 得到體重生長方程:
雌魚:Wt=3016.8[1–e–0.096(t–0.282)]2.931
雄魚:Wt=2798.9[1–e–0.090(t+0.011)]2.931
雌魚和雄魚群體的表觀生長指數(?)分別為4.5243和4.4740。
弧唇裂腹魚的年齡和體長呈現一定的關系(圖7)。在低于10齡時, 相同年齡雌雄性的體長非常相近,之后隨著年齡的增長, 差異越來越大, 相同年齡的雌性體長大于雄性。
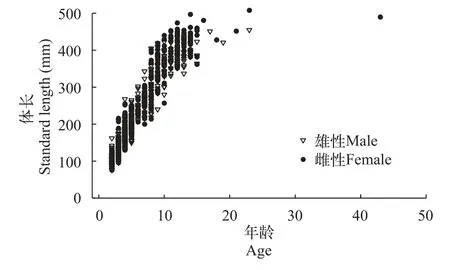
圖7 弧唇裂腹魚的年齡與體長關系圖Fig.7 The relationship between age and standard length of S.curvilabiatus
生長速度和加速度將弧唇裂腹魚的生長方程經過一階求導得到其生長速度方程, 再通過二階求導獲得其生長加速度方程。
雌魚:dL/dt=56.66e–0.096(t–0.282)
dL2/dt2= ?5.44e?0.096(t–0.282)
dW/dt=848.85e–0.096(t–0.282)[1–e–0.096(t–0.282)]1.931
dW2/dt2=81.49e–0.096(t–0.282)[1–e–0.096(t–0.282)]0.931[2.931e–0.096(t–0.282)–1]
雄魚:dL/dt=51.78e–0.090(t+0.011)
dL2/dt2=–4.66 e–0.090(t+0.011)
dW/dt=738.33 e–0.090(t+0.011)[1–e–0.090(t+0.011)]1.931
dW2/dt2=66.45e–0.090(t+0.011)[1–e–0.090(t+0.011)]0.931[2.931e–0.090(t+0.011)–1]
弧唇裂腹魚雌魚體長增長的速度和加速度的變化趨勢與雄魚相似(圖8a和8b)。隨著年齡的增長, 雌魚和雄魚的體長增長速度均呈遞減趨勢, 且遞減速度逐漸減緩, 無限接近于0; 雌魚和雄魚體長增長的加速度隨著年齡的增長呈遞增趨勢, 遞增速度逐漸減緩, 無限接近于0而小于0。
弧唇裂腹魚雌魚的體重加速度變化趨勢與雄魚相似(圖8c和8d), 均呈先升后降的趨勢。當體重生長加速度為0時, 體重生長速度達到最大值, 此為該魚的體重生長拐點。弧唇裂腹魚雌魚拐點年齡(ti)為11.5齡, 雄魚拐點年齡(ti)為11.9齡, 對應的體長分別為388.8和379.0 mm; 對應的體重分別為887.9和823.8 g。

圖8 弧唇裂腹魚的生長速度和生長加速度Fig.8 Growth rate and acceleration of S.curvilabiatus
3 討論
3.1 年齡鑒定
已有的研究顯示, 鱗片、脊椎骨、耳石、鰭條、匙骨和鰓蓋骨等鈣化組織是裂腹魚類年齡鑒定的常用材料[16—24]。不同年齡材料輪紋清晰的程度因研究物種、個體生長差異而不同。劉艷超等[23]用微耳石、星耳石、脊椎骨、鰓蓋骨、臀鱗、胸鱗、側線鱗和背鱗8種年齡材料對西藏拉薩河上游雙須葉須魚的年齡特征進行了比較, 認為微耳石最適宜用于雙須葉須魚的年齡鑒定。Sabah和Khan[24]-利用耳石和脊椎骨等年齡材料對克什米爾河谷三種裂腹魚的年齡和生長進行比較研究, 結果表明耳石用于中唇裂腹魚S.curvifrons和S.niger的年齡鑒定較脊椎骨更準確, 而用脊椎骨鑒定扁嘴裂腹魚S.esocinus的年齡比耳石準確。Ma等[16]和Zhou等[22]的研究結果顯示, 脊椎骨鑒定兩種裂腹魚屬魚類的年齡時, 低齡魚的鑒定結果與耳石差異不大; 而在鑒定高齡魚時, 其結果顯著低于耳石; 相比較耳石和脊椎骨, 鰓蓋骨的輪紋比較密集難以區(qū)分, 且對于高齡魚而言, 干擾紋偏多, 容易造成誤判。
在本研究中, 鰓蓋骨輪紋清晰度不及耳石和脊椎骨, 其年輪讀數也顯著低于耳石和脊椎骨; 而脊椎骨在低齡魚中(低于10齡)與耳石的鑒定結果相當, 但在高齡魚中(10齡以上)年齡判讀結果的準確率和辨識度均較耳石差。綜上所述, 耳石是鑒定裂腹魚類年齡較為合適的材料[16,19—23]。采用體外標記和氯化四環(huán)素熒光標記方法也表明, 用耳石鑒定魚類年齡比鱗片更為準確, 尤其是生長緩慢的長壽命魚類[6]。主要原因在于耳石生長獨立于機體生長, 在慢生長和高齡個體中, 耳石比其他骨質材料生長更快, 且不會存在輪紋的重吸收現象, 故其更能如實反映周期性的季節(jié)生長, 并以規(guī)律性輪紋呈現為年齡[2,25]。
3.2 生長特性
由于高原特殊的生態(tài)環(huán)境, 裂腹魚類往往都具有較高的壽命, 且雌性個體年齡大于雄性個體。已有的報道顯示, 雅魯藏布江異齒裂腹魚雌雄個體的最大年齡分別為50齡和40齡[26], 雙須葉須魚雌雄個體的最大年齡分別為45齡和24齡[17], 拉薩裂腹魚雌雄個體的最大年齡分別為40齡和37齡[22]。在本研究中, 弧唇裂腹魚雌魚和雄魚的最大年齡分別為43齡和23齡, 與上述研究結果接近。
生長系數k和表觀生長指數(?)都是評估魚類種群生長潛力的關鍵參數[2]。Branstetter[28]將生長系數k分為三類: 0.05—0.10屬生長緩慢型, 0.10—0.20屬均速生長型, 0.20—0.50屬快速生長型。生長方程的擬合結果顯示, 弧唇裂腹魚的生長系數k值低于0.1, 其中雌性為0.096, 雄性為0.090; 雌性和雄性群體的表觀生長指數(?)分別為4.5243和4.4740。上述參數與Zhou等[22]、Ma等[26]、朱秀芳和陳毅峰[29]、Li and Chen[30]和Huo等[31]對雅魯藏布江中上游裂腹魚類的研究結果一致(表6)。同時, 弧唇裂腹魚的最大年齡和漸進體長L∞較高(雌性為590.2 mm, 雄性為575.3 mm)。綜上可以判斷, 弧唇裂腹魚為生長緩慢, 壽命較長的魚類, 屬于典型的K-對策選擇者[32]。

表6 文獻中雅魯藏布江裂腹魚屬魚類生長特性對比Tab.6 Comparison of growth characters of Schizothorax fishes in the Yurlung Zangbo River in different studies
魚類個體生長是內源因子和外源因子共同作用的結果。除物種間的遺傳差異外, 食物資源、溫度、光照及其他環(huán)境因子也會影響魚類的個體生長[2]。由于入侵水域營養(yǎng)生態(tài)位的空缺, 加之適口餌料豐度較大, 撫仙湖的黃顙魚種群表現出很好的生長潛力[33]。近年來, 烏倫古湖餌料生物的數量和生物量較1987年均大為減少, 河鱸(Perca fluviatilisLinnaeus)的喜好餌料搖蚊幼蟲生物量僅為1987年的1/10, 從而導致河鱸營養(yǎng)狀況不佳, 影響河鱸生長速度[34]。雅魯藏布江下游河谷深切, 水流湍急,流速可達16 m/s, 加之地質活動頻繁, 滑坡和泥石流時有發(fā)生, 導致干流河道水生生物餌料資源相對匱乏[1,35,36]。故推測弧唇裂腹魚生長緩慢是對食物資源匱乏等惡劣環(huán)境的一種長期適應[2,4,35]。此外, 同為植食性的裂腹魚亞屬魚類, 弧唇裂腹魚的表觀生長指數(?)高于異齒裂腹魚, 可能與雅魯藏布江下游海拔較低, 水溫相對較高有關。水溫較高使弧唇裂腹魚具有較高的新陳代謝速率, 能量轉換效率可能高于異齒裂腹魚。弧唇裂腹魚的表觀生長指數與謝從新等[5]對拉薩裂腹魚和巨須裂腹魚的研究結果較為接近, 但高于朱秀芳和陳毅峰[30]對巨須裂腹魚的研究結果, 可能與所用年齡材料不同有一定關系(表6)。
3.3 弧唇裂腹魚的保護管理
研究指出, K-對策選擇魚類對環(huán)境變化非常敏感, 種群一旦衰退, 其資源恢復速度緩慢[32]。近年來, 人為活動對雅魯藏布江下游河流環(huán)境的干擾逐漸加大。一方面隨著旅游開發(fā)、人口經濟的快速增加、墨脫河段的漁業(yè)捕撈強度逐步增大, 導致弧唇裂腹魚種群以3—5齡為主, 種群低齡化現象突出;另一個方面隨著國家“十四五”規(guī)劃的深入推進, 涉水工程及沿江公路的實施將改變雅魯藏布江下游的河流形態(tài)、水文情勢及水體理化性質, 進而對弧唇裂腹魚等土著魚類的棲息產生不利影響。
針對上述問題, 并結合弧唇裂腹魚的生物學特性及流域魚類的保護現狀, 提出了以下措施與保護建議: (1)控制漁業(yè)捕撈是保護魚類資源的重要手段。弧唇裂腹魚拐點年齡為11齡, 對應的體長體重分別為380 mm和820 g。為此, 建議將其起捕規(guī)格限制在800 g以上, 同時加快完善漁業(yè)資源管理體系建設, 嚴厲打擊非法捕撈。(2)棲息地保護是物種保護最有效的措施之一。雅魯藏布江下游水域生態(tài)學的研究基礎薄弱, 建議加強弧唇裂腹魚產卵場、越冬場和索餌場等重要生境的調查與研究, 在此基礎上選擇合適的江段建設弧唇裂腹魚水產種質資源保護區(qū), 加強其種質資源的保護。(3)目前雅魯藏布江中上游異齒裂腹魚、拉薩裸裂尻魚(Schizopygopsis younghusbandi)等主要魚類的人工繁殖已獲得成功, 形成了較為系統的規(guī)模化人工繁育技術體系[2]。相比之下, 雅魯藏布江下游魚類的人工繁育技術尚不成熟, 建議加強弧唇裂腹魚的人工繁殖及苗種批量培育等關鍵技術的研究, 發(fā)展可持續(xù)的人工養(yǎng)殖, 減少對弧唇裂腹魚野生種群資源的依賴。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
