水稻粒形基因的遺傳研究進展
2022-12-10 09:27:10于雪然杜懷東李培富
江蘇農業科學 2022年21期
田 穎, 于雪然, 杜懷東, 田 蕾, 李培富
(寧夏優勢特色作物現代分子育種重點實驗室/寧夏大學農學院,寧夏銀川 750021)
水稻(OryzasativaL.)是我國重要的糧食作物之一,大約50%以上的人口以稻米為主食。育種家對矮桿基因和雜種優勢進行了廣泛的應用,在這之后我國的水稻產量開始顯著提高。但在人口增長和耕地減少的嚴峻形勢下,提高并穩定水稻產量仍然是我國水稻育種的主要研究方向[1]。隨著人民生活水平的逐漸改善,消費者對稻米的需求日益多樣化,稻米品質也更加受到重視。因此,如何增加我國水稻產量和有效改良稻米品質等關鍵問題,已經成為現代水稻育種研究的主要目標。研究證明,水稻產量是由多個基因控制的農藝性狀,主要由以下3個因素決定:有效穗數、每穗粒數及千粒質量[2],水稻粒形、籽粒大小和灌漿充實度都顯著影響著千粒質量[3]。水稻粒形主要由粒長、粒寬和長寬比等組成。有研究表明粒形不僅與千粒質量關系密切,而且在一定程度上影響稻米品質[4-5]。育種家認為,改善水稻粒形對促進我國水稻優質育種的發展和稻米品質的提高具有重要的研究意義[6]。近年來,許多已定位和克隆的水稻粒形基因對于闡明粒形的調控機制有著重要的研究意義,為我國水稻的高產優質育種奠定了關鍵的理論基礎和遺傳資源。
1 水稻粒形的遺傳特性
水稻粒形是由多基因控制的性狀,主要為加性效應,是數量性狀遺傳。在早期研究中,芮重慶等選用不同粒形的6個秈稻品種進行研究發現,加性效應在水稻粒長的遺傳中起著決定性的作用,并且細胞核與細胞質之間可能還存在著一定的互作效應[7]。石春海等利用不同的雜交組合研究粒形的遺傳,發現粒長是一個由多基因調控的數量性狀,其中加性效應在遺傳中占比較高[8]。劉金波等使用了主基因+多基因混合遺傳模型,對水稻籽粒長度進行了遺傳分析,認為粒長的遺傳主要是加性效應[9]。
大多數研究表明,多基因的加性效應及顯性效應均控制水稻粒寬,主要受加性效應的控制,遺傳力高,顯性方向隨組合的變化而發生變化[10-11]。呂勇統計了突變體植株F2代群體的粒寬數據,對粒寬作了頻率分布直方圖,發現粒寬在圖中呈現連續正態分布,據此可證實水稻粒寬受到了多基因的控制[12]。石春海等采用多個雜交組合進行研究后發現,水稻粒寬的遺傳主要表現為加性效應[13]。
符福鴻等對三系雜交稻的粒形進行遺傳分析,結果表明加性效應在水稻籽粒長寬比的遺傳中起主導作用[14]。石春海研究發現一些水稻組合具有良好的配合力,加性效應在籽粒長寬比中表現顯著[4]。方平平等采用雙列雜交設計,發現水稻籽粒長寬比主要受到了種子的直接遺傳效應影響[15],在早期的研究中卻發現粒形受母體基因型控制[8,16-21]。
粒形性狀的遺傳存在著一因多效的現象,并且不同粒形性狀之間的相關性存在著差異。石春海等的研究表明,粒長與粒寬之間存在著極顯著的負相關,而與長寬比則表現為顯著的正相關,粒寬與長寬比之間呈現出極顯著的正相關[8]。林荔輝等利用重組自交系群體研究發現粒長與粒寬的相關系數值較小,不存在明顯的相關性[22]。
2 水稻粒形基因的克隆
目前,有很多影響水稻粒形調控的基因已經被克隆(圖1、表1),各個基因調控水稻粒形的方式不同,主要包含轉錄因子調控、G蛋白信號、泛素途徑和植物激素途徑[23]。
GS3是第1個被定位并克隆的調控水稻粒長的主效基因,Fan等用明恢63和川7進行連續雜交和回交,發現該基因編碼的跨膜蛋白可負調控粒長和粒質量[24]。Zhang等選用小粒N643和大粒N411進行雜交,克隆得到一個負調控粒長的主效基因qGL3[25]。 Gao等在研究中發現qGL3可去磷酸化OsGSK3蛋白并使其穩定,進而抑制油菜素內酯信號傳導[26]。GL4是在非洲野生稻W1411中發現并克隆的可調控粒長和粒質量的數量性狀基在座(QTL),非洲栽培稻的籽粒長度由于該基因調控內外穎縱向細胞的伸長而受到影響,同時該基因也可以調節種子的落粒性,繼而影響水稻的千粒質量和單株產量[27]。Ishimaru等利用Nipponbare和Kasalath的回交自交系定位到一個調控粒長和千粒質量的基因TGW6,編碼吲哚乙酸(IAA)-葡萄糖水解酶,該基因功能缺失會影響IAA的供應,進而調控籽粒長度及籽粒質量從而使水稻增產[28]。Yu等從504份栽培稻品種中克隆了一個粒長基因OsLG3,該基因可在不降低稻米品質的同時正向調控水稻籽粒長度,有效提高產量[29]。Xiong等認為過表達OsLG3還可顯著提高水稻的耐旱性[30]。Si等采用全基因組關聯分析(GWAS)的方法,在381個不同水稻品種中克隆了一個調控粒長的主效基因GLW7,編碼OsSPL13,可以通過促進穎殼細胞擴張正向調控水稻的籽粒大小[31]。Heang等發現PGL1是一個正調控籽粒長度的基因,過量表達PGL1時內穎細胞長度會增加,進而引起了水稻籽粒長度和千粒質量增加[32]。
GW2是Song等利用小粒FAZ1和大粒WY3構建F2群體后定位并克隆的主效基因,主要調控粒寬和粒質量,其編碼的環型E3泛素連接酶對細胞的分裂起負調節的作用,GW2的WY3等位基因對水稻籽粒的寬度和千粒質量具有顯著的提高作用[33]。qSW5/GW5/GSE5編碼了一個鈣調素結合蛋白,該基因功能的缺失會增加穎花的外殼細胞數量,從而導致穎花的外殼細胞變寬,水稻籽粒的千粒質量以及產量都會顯著提高[34-36]。Li等用寬粒珍汕97和細長粒H94構建了群體,發現克隆了基因GS5,是控制水稻粒寬、籽粒灌漿和千粒質量的數量性狀基因[37],研究進一步發現該基因還可能受到脫落酸(ABA)的調控[38]。GW8/OsSPL16是由Wang等定位發現的可正調控水稻粒寬和粒質量的基因,研究發現該基因的缺失和低表達導致籽粒變長,相反,該基因高表達會加快細胞分裂和籽粒灌漿,使得水稻籽粒變得充實并且寬度增加,可有效提高水稻產量[39-40]。Shi等發現GW6通過調控細胞生長而非細胞增殖來調控水稻粒寬,GW6通過調節赤霉素(GA)的應答和生物合成,正調控赤霉素信號通路[41]。Sun等研究發現了屬于GRAS基因家族成員的基因GS6,將含有突變體gs6的水稻與野生型水稻的粒形性狀對比后發現,粒寬和粒質量均明顯增加,并且內外稃增厚有更多的凸起和毛狀物,鄰近表皮的厚壁細胞也開始增多[42]。
GL7/GW7是一個可以同時調控粒寬和粒長的主效基因,GL7編碼的LONGIFOLIA蛋白正調控水稻細胞的縱向伸長;GW7表達量上調時水稻籽粒會變得細長,原因是該基因的表達增加了水稻籽粒縱向細胞的分裂,同時橫向細胞的分裂減少[43]。Zhao等構建F3群體定位并克隆得到了基因GS9,可負調控水稻籽粒長寬比,研究發現GS9可能受到油菜素內酯信號的調控[44]。GL2/GS2編碼了生長調節因子OsGRF4,表達量增加時加快了細胞分裂,細胞也變大,進而增加了籽粒的千粒質量及水稻產量[33,45-46]。Ying等利用黃華占(HHZ)和JZ1560構建重組自交系(RIL)群體檢測出調控籽粒長寬比的基因TGW3,編碼一個類似糖原合酶激酶GSK3的激酶,能協同改變穎殼中細胞的大小和數量,負調控水稻籽粒長寬比[47]。

表1 已克隆的水稻粒形相關基因

表1(續)
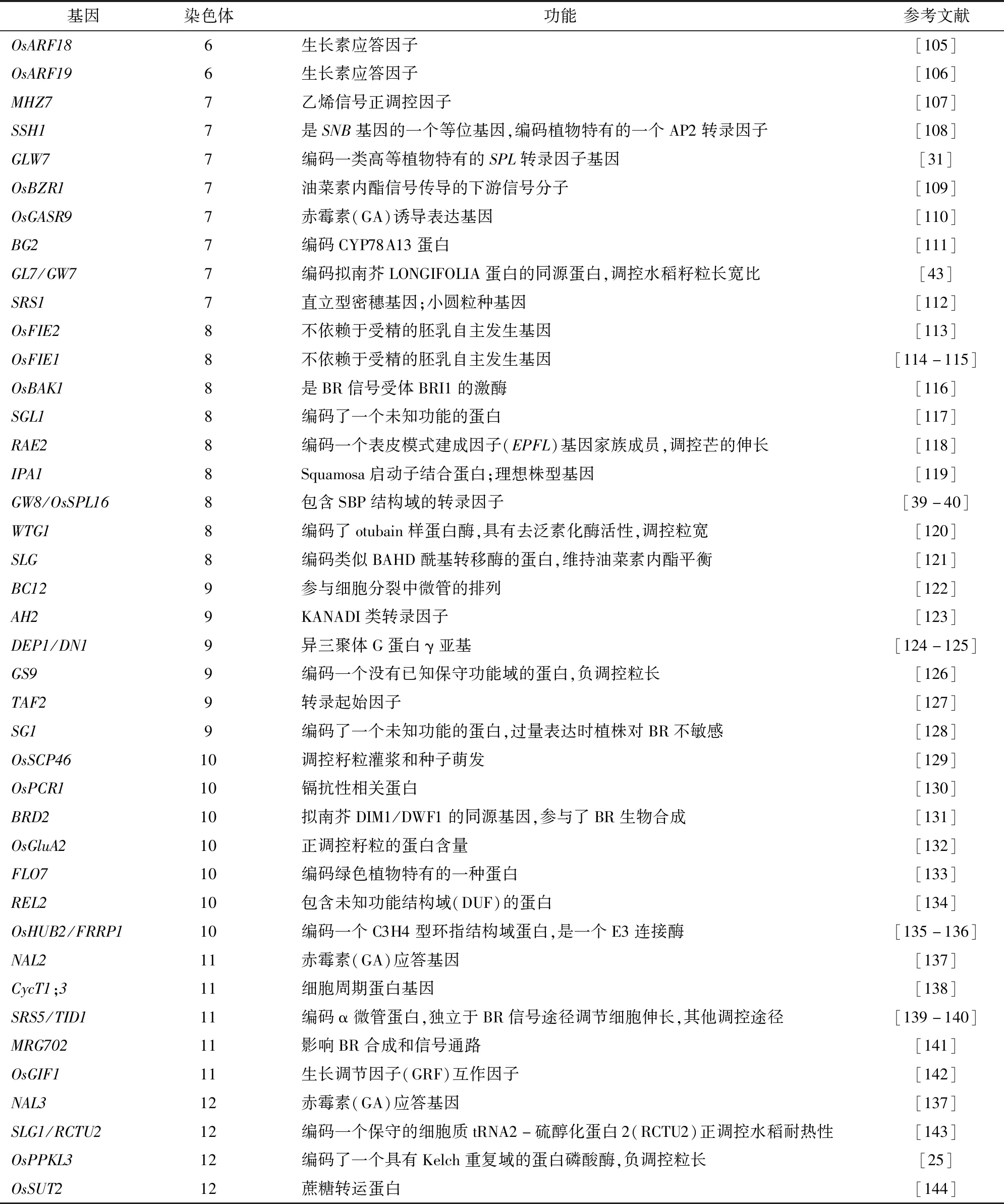
表1(續)
3 水稻粒形基因間的互作
粒形是由多基因調控的數量性狀,不同的粒形基因之間存在著不同程度的相互作用。將這些基因之間的互作原理應用于水稻育種,對提高水稻產量和有效改良粒形有著重要的意義。Yan等通過基因表達分析,來探究GS3、GW2、qSW5、GIF1等4個基因之間的關系,結果表明,對GW2的轉錄進行抑制可以降低qSW5的表達,而GW2和qSW5都會促進GS3的表達,GIF1的表達,受到了GW2和GS3的負向調節,但qSW5可以正調控其表達量;闡明GS3、GW2、qSW5/GW5和GIF1這4個基因的關系,可加速基因聚合育種的進程[141]。Li等發現HGW編碼的上游調控蛋白,可以有效增加抽穗和粒質量基因的表達量,該基因很有可能直接通過GIF1基因的作用來調控水稻粒形和產量。進一步研究了HGW通過GIF1、GW2、GW5和GS3調節粒形和籽粒質量后發現,在轉錄水平下GIF1基因的表達量減少了90%,而其他基因的表達量則大約減少了20%~50%[97]。Gao等使用了雙向方差分析的方法,研究了GS3、qGL3和GS3×qGL3相互作用對水稻粒長的影響,發現GS3和qGL3對粒長的調控表現為累加效應[142]。Wang等研究發現,GW8包含的SBP結構域,可以與GW7啟動子直接結合,使GW7的表達量降低,從而調控了水稻粒寬[39-40]。Hao等研究發現,WG1/OsGRX8調控水稻籽粒大小,是粒寬方向細胞增殖所必需的,GW2可以與WG1互作并將其泛素化,從而調控WG1蛋白的穩定性[143]。Shi等研究發現OsUBP15可能與GW2存在著某種關系,都通過泛素途徑對水稻粒形進行調控[144]。Xia等研究發現,GL3.3與GS3在遺傳上有上位互作效應,這種相互作用可以調控水稻籽粒的大小,目前已經應用于大粒粳稻的育種中[23]。Duan等發現GS2基因和OsGIFs互作后,對細胞增殖和細胞伸長都產生了影響,進而影響了水稻籽粒大小和產量,OsGIF1正調控水稻籽粒的發育[138]。
4 粒形基因在水稻育種上的應用
水稻粒形是影響千粒質量的重要數量性狀,并且直接決定了水稻的品質,將粒形基因合理地應用在水稻育種中,可有效提高水稻產量、改良水稻品質。張劍霞在珍汕97B中聚合了抗白葉枯病基因Xa23和粒長基因GS3,獲得了改良型珍汕97B,是有效抗白葉枯病并且具備良好外觀品質的新品種[145]。楊梯豐等使用分子聚合育種,得到了具有GS3和其他優異基因的純合聚合系,經過測定發現水稻籽粒變長,有效改良了外觀品質[146]。Nan等通過更新孔玉131的GS3位點,發現其粒長、百粒質量和單株總粒質量分別提高了12.05%、16.30%、4.47%[147]。Song等研究發現,含有GW2基因的豐矮占1號細胞數量會增加,并加快了水稻的灌漿速度,籽粒寬度和千粒質量也顯著增加,有效提高了單株產量[33]。Zhao等通過遺傳相互作用分析后發現,GS9的功能獨立于其他先前確定的影響水稻籽粒大小的基因,將等位基因gs9引入優質水稻栽培品種中,可以顯著改善水稻粒形和外觀品質,這表明GS9不論單獨或與其他粒形基因相結合,都可以培育出具有優化粒形的良好水稻品種[44]。Wang等研究發現,相對于親本日本晴,含有GL7基因的近等基因系,水稻籽粒長寬比明顯增加,大大減少了稻米中的堊白粒率和堊白度,使外觀品質得到了極大改善。這表明GL7的有益等位基因和其他與產量和品質相關的基因聚合有益于優良水稻品種的選育[43]。Hu等構建近等基因系研究發現,攜帶GS2/OsGRF4等位基因的水稻品種可以顯著提高千粒質量和產量,可在培育高產水稻品種中得到應用[46]。Li等利用標記輔助回交將GW6基因導入秈稻9311和粳稻中花11中,改良后的水稻籽粒長度、千粒質量和單株產量均有所提高,為培育大粒和高產水稻品種提供了依據[148];Shi等將南洋占的GW6NYZ導入秈稻華粳秈74,可使單株產量提高14.49%,大田產量提高10.44%,將GW6NYZ導入武運粳7號,發現其大田產量提高7.97%,這說明GW6對秈稻和粳稻都有潛在增產作用[41]。Wang等發現GIF1基因在水稻籽粒發育時,控制著蔗糖酶(轉化酶)的活性。在現代栽培稻中含有GIF1基因的籽粒灌漿速率明顯提高,產量也顯著增加[79]。
5 存在問題及展望
經過多年來育種學家的不斷努力,許多與水稻粒形相關的基因都已經被成功克隆。本文歸納總結已定位克隆的粒形相關基因的作用,簡單介紹多基因間的相互作用,并對粒形基因在品質改良和育種中的應用進行詳細描述。
通過總結發現,近年來育種家在水稻粒形基因的定位、克隆及功能方面已經有了很大的進步,但仍有不足的地方,需要進一步加強和改善:(1)目前已經克隆了許多調控水稻粒形的相關基因,但多基因之間的遺傳互作研究比較少見,部分粒形基因間遺傳互作的方式和關系不夠清晰,在育種中的應用也較少;(2)利用粒形基因改良水稻粒形時可能會降低水稻品質,如整精米率隨著粒長的增加而下降,堊白度隨著粒寬的增加而增加,使水稻的加工品質變差[149];(3)將同一粒形基因應用于不同的水稻品種會導致不同的表型性狀,探討為何會產生這樣的結果,可為相關基因位點的育種應用提供精確的理論依據。
為實現高產優質的育種目標,首先要選擇粒形差異大或者親緣關系遠的品種材料,選育出粒形適宜的育種材料。對已定位和克隆的水稻粒形基因,在聚合不同優質基因時,必須要打破水稻產量與品質之間的不良連鎖關系,有效改良稻米品質。隨著我國生物技術的快速發展,育種家可利用高效基因組編輯技術CRISPR/Cas9精準有效地聚合優異等位基因,精確敲除水稻品種中的不利性狀,快速獲得新材料,提高育種效率。在未來的育種工作中,要定位和克隆更多與水稻粒形相關的基因,不斷的更新和完善粒形調控網絡,為水稻的高產優質育種奠定關鍵的理論基礎和遺傳資源。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
軍事文摘(2021年22期)2021-11-26 00:43:51
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
文苑(2020年6期)2020-06-22 08:41:52
科技傳播(2019年22期)2020-01-14 03:06:54
文苑(2019年22期)2019-12-07 05:29:00
中國生殖健康(2019年2期)2019-08-23 08:12:08
