龍眼PIFs家族全基因組鑒定及其表達分析
2022-12-05 02:47:28李小芳林曉藝蘇炳茜張城瑜羅彬彬陳裕坤杜迎剛賴鐘雄
福建農(nóng)業(yè)學(xué)報 2022年8期
關(guān)鍵詞:分析
李小芳,林曉藝,蘇炳茜,張城瑜,羅彬彬,陳裕坤,杜迎剛,2 ,賴鐘雄
(1.福建農(nóng)林大學(xué)園藝植物生物工程研究所,福建 福州 350002;2.濰坊科技學(xué)院山東省高校設(shè)施園藝實驗室,山東 濰坊 261000)
0 引言
【研究意義】龍眼(Dimocarpus longanLour.)是無患子科(Sapindaceae)龍眼屬(DimocarpusLour)的熱帶亞熱帶常綠經(jīng)濟果樹,別稱桂圓和三尺農(nóng)味等,具有降血脂、補腦益智、免疫調(diào)節(jié)、抗腫瘤等功效,是一種重要的經(jīng)濟水果。龍眼胚胎的發(fā)育對龍眼坐果率及其果實品質(zhì)有重要影響,因此對龍眼胚胎發(fā)育的深入研究對龍眼產(chǎn)業(yè)的發(fā)展具有重大意義。【前人研究進展】光信號在植物生長發(fā)育過程中發(fā)揮著重要的調(diào)節(jié)作用,植物對光信號的感知主要是通過光感受器來進行,而光敏色素(Phytochromes,phy)作為一種可以感知紅光/遠紅光的植物色素蛋白,以吸收紅光的非活性形式Pr和吸收遠紅光的活性形式Pfr存在[1],在植物的生長發(fā)育中發(fā)揮重要作用[2]。光敏色素互作基因PIFs(Phytochrome interacting factors,PIFs)作為光敏色素極為重要的一種互作基因和重要的信號整合因子參與調(diào)節(jié)植物的眾多生理過程[3]。在光照下,光敏色素通過快速磷酸化、泛素化和蛋白酶體介導(dǎo)的降解促進PIF的周轉(zhuǎn),從而抑制PIF功能誘導(dǎo)轉(zhuǎn)錄重編程,導(dǎo)致光形態(tài)發(fā)育[4]。PIFs屬于basic helix-loop-helix(bHLH)轉(zhuǎn)錄因子家族的第15亞族,植物PIFs轉(zhuǎn)錄因子具有bHLH結(jié)構(gòu)域,大部分還具有保守的APB(Active phytochrome B-binding domain)結(jié)構(gòu)域,APB結(jié)構(gòu)域是PIFs與具有生物活性光敏色素相結(jié)合的不可或缺的元件[5]。PIF3是第一個被鑒定出來的與phyB、phyA互作的bHLH 轉(zhuǎn)錄因子,其他PIFs成員同樣能夠與Pfr形式的phyB發(fā)生互作[6]。PIFs 蛋白序列中保守的 bHLH 結(jié)構(gòu)域能特異性地與啟動子中的G-box元件結(jié)合從而調(diào)控靶基因的表達[7]。通過ChIP-seq(染色質(zhì)免疫共沉淀技術(shù))和RNA-seq(轉(zhuǎn)錄組測序技術(shù)) 分析發(fā)現(xiàn),PIF1、PIF3、PIF4和PIF5 都有各自直接的調(diào)控基因,但同時也發(fā)現(xiàn)有139個基因能同時被這4個PIF蛋白調(diào)控。這些結(jié)果表明 PIF蛋白具有相同和各自不同的DNA結(jié)合特性,從而表現(xiàn)出功能重疊及各自特定功能[8]。研究表明,PIFs可以協(xié)調(diào)光和激素信號傳遞[9-10],參與種子萌發(fā)[11]、避蔭反應(yīng)[12]、開花[13]等生理活動,可以負調(diào)控花青素、葉綠素的生物合成和光合作用相關(guān)基因的表達[14-16],還能響應(yīng)光、蔭、溫度和晝夜條件[17-20]。在非生物脅迫調(diào)控方面,PIFs可以調(diào)控植物對低溫、高溫的耐性[21-24],對干旱和鹽脅迫也有響應(yīng)[25]。總之,PIFs對植物的生長發(fā)育具有極為重要的作用。【本研究切入點】龍眼體胚發(fā)生系統(tǒng)是研究木本植物胚胎發(fā)生的優(yōu)良體系,可以替代合子胚的發(fā)生系統(tǒng),為研究龍眼體胚發(fā)生的分子調(diào)控機制和龍眼分子育種奠定基礎(chǔ)[26]。目前國內(nèi)外缺少對龍眼中PIFs家族的研究,更缺乏PIFs家族在龍眼體胚和不同組織器官發(fā)育過程中所發(fā)揮功能的了解。【擬解決的關(guān)鍵問題】基于龍眼基因組數(shù)據(jù)對DlPIFs家族進行鑒定及相關(guān)生物信息學(xué)分析,通過龍眼不同組織部位轉(zhuǎn)錄組、不同光質(zhì)和不同溫度處理下的轉(zhuǎn)錄組對DlPIFs家族的FPKM(Reads per kilobase per million mapped reads)值進行分析,并且利用qRT-PCR分析DlPIFs在龍眼體胚發(fā)生早期的表達模式及不同生長調(diào)節(jié)劑處理下的表達模式,為DlPIFs在龍眼體胚發(fā)生過程中的功能研究奠定基礎(chǔ)。
1 材料與方法
1.1 材料
龍眼體胚發(fā)生早期3個階段包括EC、ICpEC和GE,參照賴鐘雄等[27]的方法培養(yǎng)獲得。二代龍眼基因組數(shù)據(jù)庫(NCBI登錄號:BioProject PRJNA 3053370)和三代龍眼基因組和不同溫度處理下EC轉(zhuǎn)錄組數(shù)據(jù)庫(另文發(fā)表)、龍眼體胚發(fā)生早期轉(zhuǎn)錄組數(shù)據(jù)庫、龍眼不同組織部位轉(zhuǎn)錄組、不同光質(zhì)處理下龍眼EC轉(zhuǎn)錄組數(shù)據(jù)庫(GenBank登錄號:SRA050205)為本實驗室保存。
1.2 方法
1.2.1DlPIFs基因家族成員的鑒定 擬南芥(Arabidopsis thaliana)PIF基因家族序列從擬南芥TAIR數(shù)據(jù)庫下載(https://www.arabidopsis.org/),采用clustalX2軟件進行序列比對并轉(zhuǎn)換成Stockholm模式的HMM文件,利用HMMER 3.0的HMMbuild建立PIFs隱馬爾科夫模型,利用HMMER 3.0的HMMsearch從龍眼三代基因組數(shù)據(jù)中鑒定出DlPIFs家族成員;使用龍眼蛋白序列構(gòu)建本地Blast數(shù)據(jù)庫,以擬南芥PIF蛋白序列進行Blast搜索,進一步篩選DlPIFs家族成員,再 通 過CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)對DlPIFs家族成員數(shù)量進行最終確認,并利用以上方法從龍眼二代基因組數(shù)據(jù)中篩選并確定家族成員,最后對確定的家族成員進行命名。
1.2.2 DlPIFs蛋白序列的基本理化性質(zhì)、二級結(jié)構(gòu)、亞細胞定位分析 分別利用在線軟件ProtParam(http://web.expasy.org/protparam/)、SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl)和 Plant-mPLoc server(sjtu.edu.cn)對DlPIF蛋白序列的基本理化性質(zhì)、二級結(jié)構(gòu)及亞細胞定位進行分析。
1.2.3DlPIFs家族基因結(jié)構(gòu)和保守基序分析 使用GSDS(http://gsds.cbi.pku.edu.cn/)在線網(wǎng)站對DlPIFs的基因結(jié)構(gòu)進行分析,運用MEME在線軟件(http://meme-suite.org/tools/meme)分析DlPIFs編碼蛋白的保守基序,再使用TBtools進行基因結(jié)構(gòu)和保守基序圖的繪制。
1.2.4DlPIFs染色體定位分析 使用TBtools軟件對龍眼PIFs染色體定位圖進行繪制分析,再通過Artificial Intelligence 2020軟件對繪制的染色體定位圖進行美化。
1.2.5 植物PIFs 系統(tǒng)進化樹分析 使用 MEGA 5對龍眼、擬南芥、水稻(Oryza sativa)、甜橙[Citrus sinensis(L.) Osbeck]的PIFs蛋白序列的多重序列進行比對,使用鄰接法(Neighbor-joining,NJ)構(gòu)建進化樹,執(zhí)行參數(shù)為Poission模式、完全刪除和1 000次重復(fù)[28]。水稻蛋白序列下載于https://ricedata.cn/gene/,甜橙蛋白序列下載于http://citrus.hzau.edu.cn/orange/。
1.2.6DlPIFs家族啟動子順式作用元件分析 利用獲得的PIFs基因家族各成員起始密碼子ATG上游的2 000 bp 的啟動子序列,通過PlantCare(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析啟動子順式作用元件,并通過PlantTFDB(http://planttfdb.gao-lab.org/)對轉(zhuǎn)錄因子結(jié)合位點進行分析。
1.2.7DlPIFs家族成員在龍眼體胚發(fā)生早期、不同組織器官、不同光質(zhì)和不同溫度處理下的FPKM值分析 參照龍眼不同組織器官(供試紅核子龍眼的根、莖、葉、雄花、雌花、幼果、果肉、果皮和果核等不同組織器官的材料均采自福建農(nóng)林大學(xué)創(chuàng)業(yè)園)、不同光質(zhì)[將龍眼EC置于均光培養(yǎng)箱內(nèi),以黑暗處理作為對照,分別對其進行黑暗、藍光、紅光和白光處理,濕度55%~60%,溫度為(25±2) ℃][29]及不同溫度[將龍眼EC材料在25 ℃條件下培養(yǎng)兩天后,再放入不同溫度(15 ℃、25 ℃和35 ℃)培養(yǎng)箱中處理24 h]處理下轉(zhuǎn)錄組數(shù)據(jù)庫中DlPIFs的基因表達水平,對各成員的表達趨勢進行分析,使用TBtools繪制熱圖。
1.2.8DlPIFs家族成員在體胚發(fā)生早期和不同生長調(diào)節(jié)劑處理下的表達模式分析 首先采用TransZol Up試劑盒提取龍眼體胚發(fā)生過程中3個階段的龍眼愈傷和5種生長調(diào)節(jié)劑處理的龍眼EC的RNA,參照 PrimeScript? IV 1st Strand cDNA Synthesis Mix 試劑盒進行cDNA合成。
再使用DNAMAN 7.0軟件對DlPIF家族成員序列進行qRT-PCR引物設(shè)計(表1),以FSD-Q作為內(nèi)參基因[30],對DlPIFs在龍眼體胚發(fā)生早期不同階段和龍眼EC在不同生長調(diào)節(jié)劑處理下的表達水平進行qRT-PCR檢測,采用2-△△Ct法進行相對表達量的計算,每個處理3個重復(fù)。利用Excel軟件對試驗數(shù)據(jù)進行分析,利用Graphpad prism軟件進行折線圖和柱狀圖的繪制,并使用SPSS 26軟件中單因素ANOVA的鄧肯法對數(shù)據(jù)進行差異顯著性分析(P<0.05)。

表1 DlGRF基因家族qRT-PCR引物Table 1 qRT-PCR primers of DlPIFs
2 結(jié)果與分析
2.1 DlPIFs家族成員的鑒定
基于龍眼基因組三代測序數(shù)據(jù),共鑒定出8個DlPIFs基 因 家 族 成 員(Dlo12584、Dlo015892、Dlo034359、Dlo028134 、Dlo014068 、Dlo011892、Dlo31044、Dlo012986),它們都具有bHLH保守結(jié)構(gòu)域(圖1),根據(jù)與擬南芥PIFs序列比對的分析結(jié)果,將DlPIFs各成員分別命名為DlPIF1-1、DlPIF1-2、DlPIF3、DlPIF4、DlPIF5、DlPIF6、DlPIF7、DlPIF8。經(jīng)DNAMAN7.0軟件分析發(fā)現(xiàn),其CDS序列長度依次為 1 605、1 197、2 298、1 617、975、1 695、1 362 和1 431 bp(表2)。基于龍眼基因組二代測序數(shù)據(jù),共鑒定出6個DlPIFs基因家族成員(Dlo_000274.1、Dlo_031800.1、Dlo_000977.1、Dlo_018063.1、Dlo_012166.1、Dlo_028817.1)。利用 DNAMAN 7.0進行序列比對,發(fā)現(xiàn)龍眼基因組二代測序數(shù)據(jù)中所確定成員均與龍眼基因組三代測序數(shù)據(jù)中所確定成員一致。

圖1 龍眼PIFs保守結(jié)構(gòu)域Fig.1 Conserved domain of DlPIFs
2.2 DlPIFs蛋白序列的基本理化性質(zhì)、二級結(jié)構(gòu)及亞細胞定位分析
理化性質(zhì)分析表明DlPIF1~DlPIF8編碼的氨基酸殘基數(shù)分別為534、398、765、538、324、540、453和476個,分子量分別為59 036.32、43 473.33、82 158.84、58 258.97、35 089.75、62 892.14、49 786.89和50 818.37 Da,等電點(pI)分別為6.39、5.44、6.10、6.73、5.48、9.21、8.39 和 8.55,DlPIFs均為不穩(wěn)定、親水性蛋白(表2)。
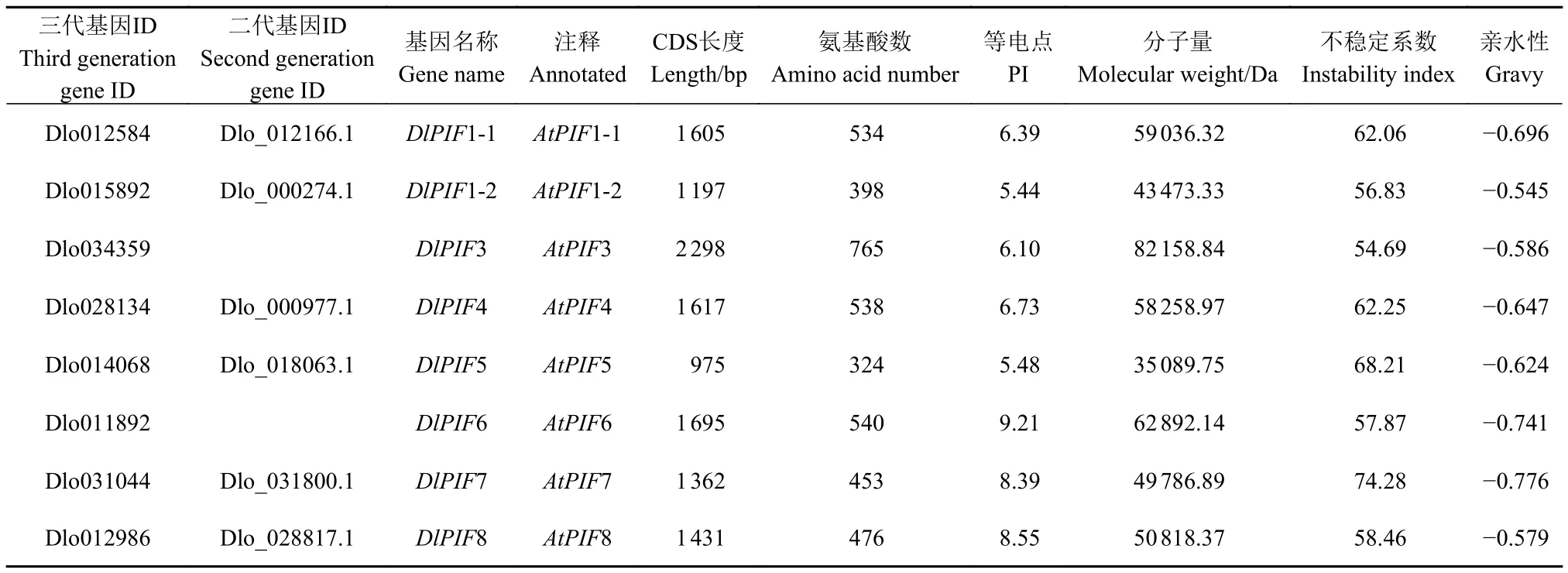
表2 龍眼PIFs成員基本理化性質(zhì)Table 2 Basic physicochemical properties of DlPIFs
亞細胞定位預(yù)測結(jié)果表明DlPIFs定位于細胞核。二級結(jié)構(gòu)分析發(fā)現(xiàn)無規(guī)卷曲所占比例最高,α-螺旋次之,延伸鏈較低,β-轉(zhuǎn)角最低(表3)。
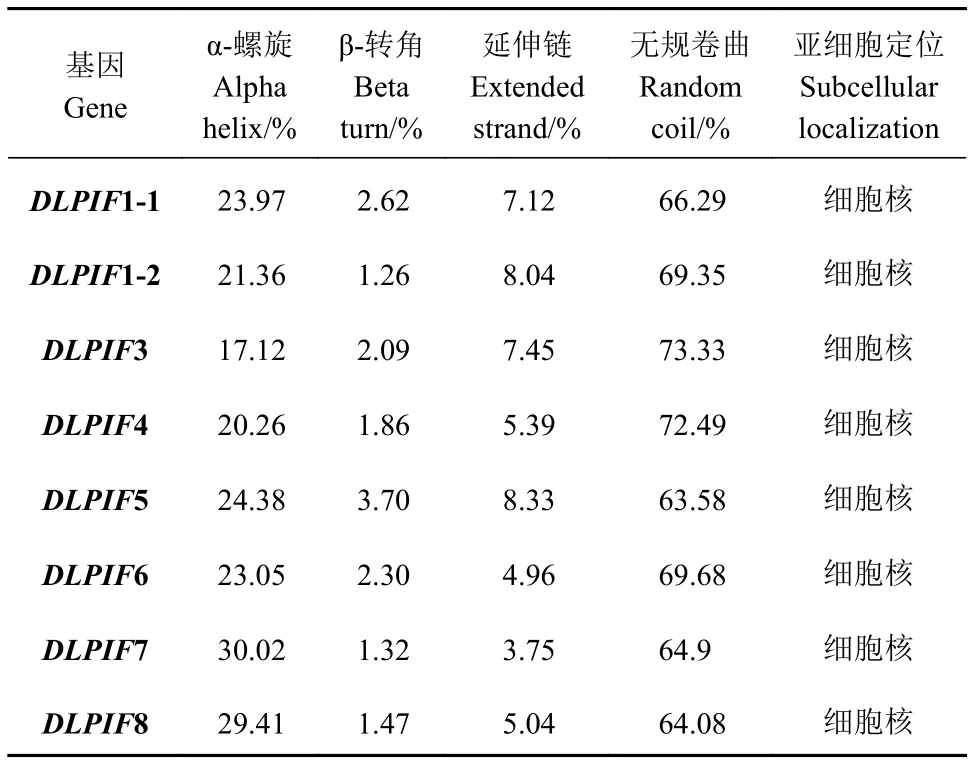
表3 龍眼PIFs家族二級結(jié)構(gòu)及亞細胞定位Table 3 Secondary structure and subcellular localization of DlPIFs
2.3 DlPIFs家族基因結(jié)構(gòu)、保守基序及染色體定位分析
通過對DlPIFs家族成員的基因結(jié)構(gòu)進行分析發(fā)現(xiàn),DlPIF1-1和DlPIF5有4個內(nèi)含子和5個外顯子;DlPIF1-2和DlPIF4有6個內(nèi)含子和7個外顯子;DlPIF3、DlPIF6和DlPIF7有5個內(nèi)含子和6個外顯子;DlPIF8有7個內(nèi)含子和8個外顯子(圖2)。
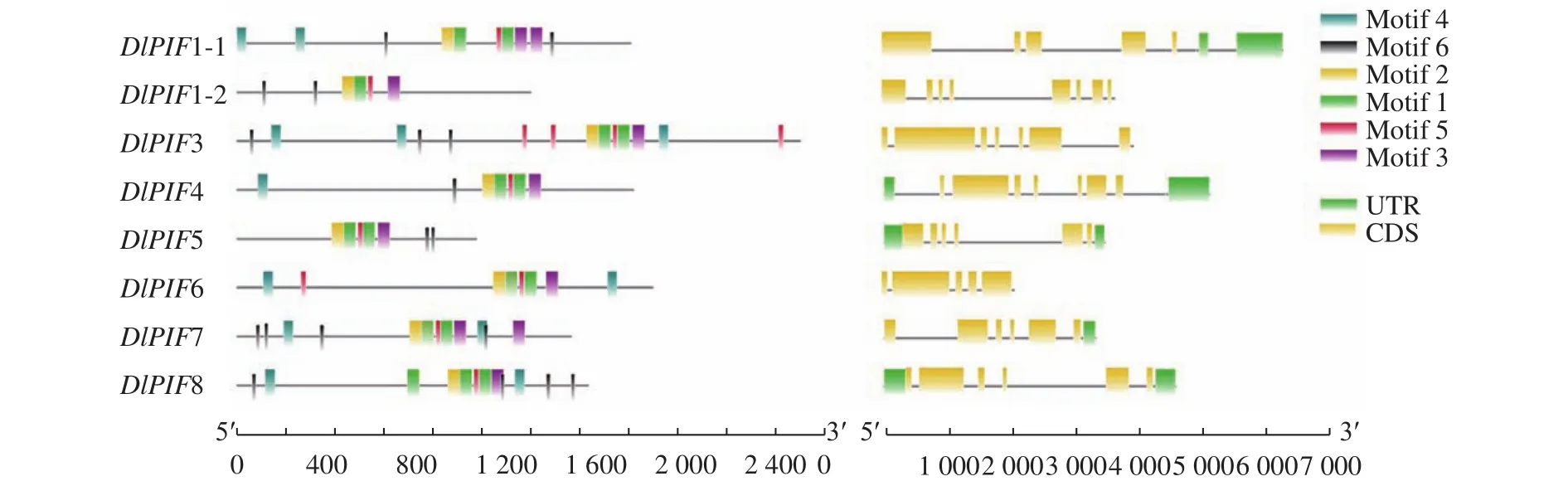
圖2 龍眼PIFs家族基因結(jié)構(gòu)和蛋白質(zhì)保守基序分布Fig.2 Gene structure and protein conserved motifs of DlPIFs
使用MEME對DlPIFs的蛋白序列進行保守基序分析,共獲得6個保守基序,8個DlPIF家族成員均含有這6個保守基序。
DlPIFs染色體定位結(jié)果見圖3,結(jié)果表明7個家族成員定位在染色體上。其中DlPIF1-1、DlPIF6和DlPIF8均定位在chr5染色體上,DlPIF5、DlPIF1-2、DlPIF4和DlPIF7分別定位在chr6、chr7、chr13和chr15上。DlPIF3并未定位在染色體上,推測其為葉綠體或線粒體基因。
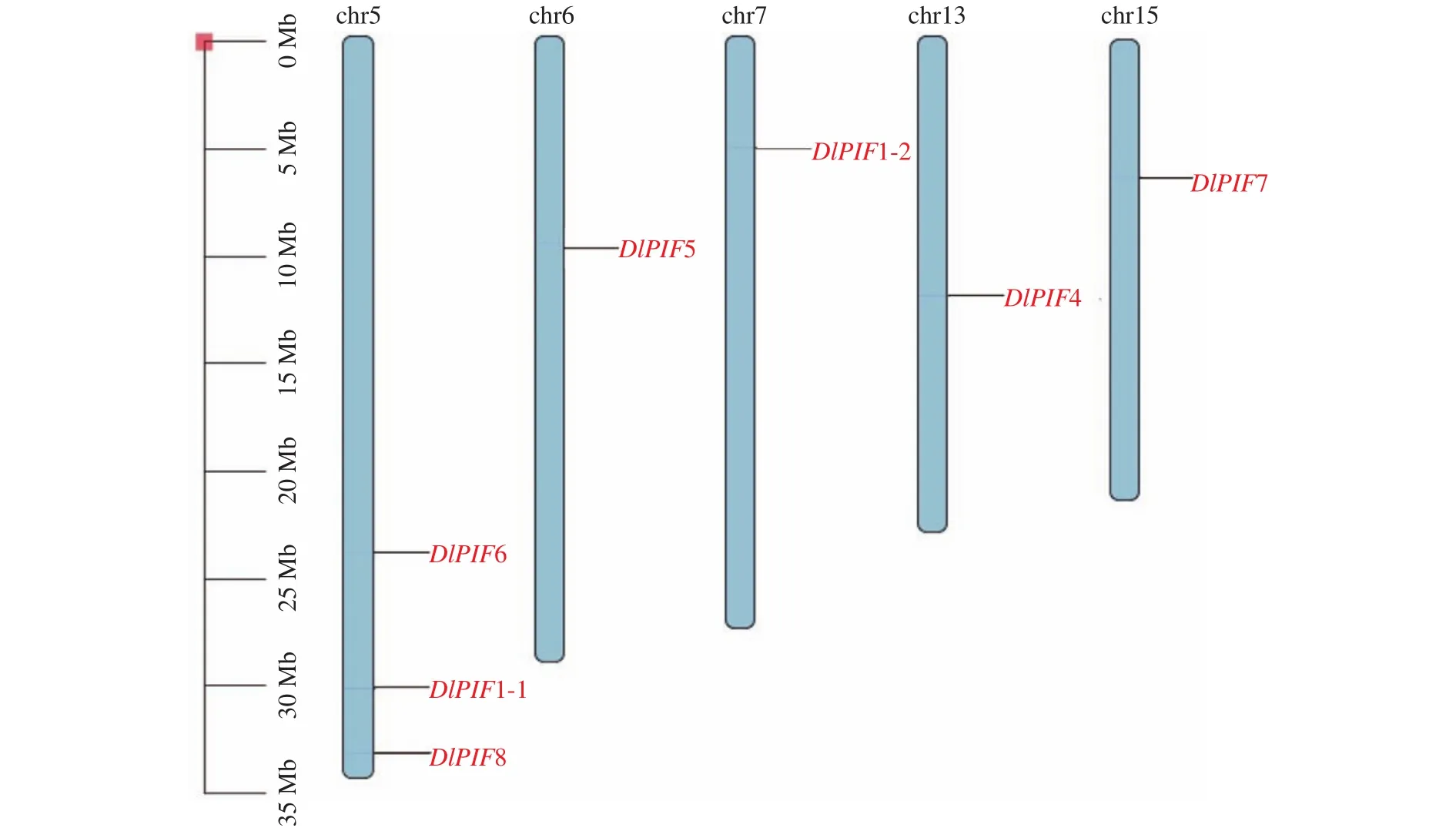
圖3 DlPIFs染色體定位Fig.3 Chromosome distribution of DlPIFs
2.4 DlPIFs家族啟動子順式作用元件分析
利用PlantCARE在線預(yù)測分析DlPIF基因家族的啟動子順式作用元件發(fā)現(xiàn),DlPIFs家族成員啟動子區(qū)均包含TATA-box和CAAT-box兩個核心元件(圖4)。DlPIFs家族的啟動子區(qū)還包含多種作用元件,且家族成員間的作用元件的數(shù)量和種類存在一定差異。
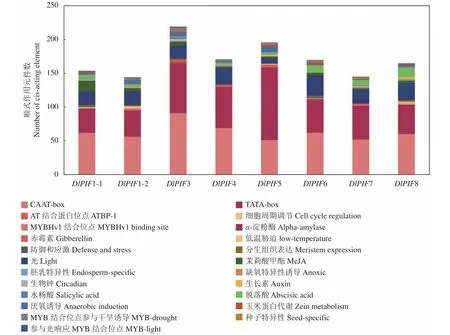
圖4 DlPIFs家族啟動子順式作用元件分析Fig.4 Cis-acting elements of DlPIFs
DlPIFs基因家族成員均包含光響應(yīng)相關(guān)的作用元件,數(shù)量相似,DlPIF1-2、DlPIF3和DlPIF7還含有MYB結(jié)合位點參與光反應(yīng)(MRE)相關(guān)的作用元件。DlPIF基因家族啟動子區(qū)包含5種激素響應(yīng)相關(guān)的作用元件。DlPIFs家族成員均包含茉莉酸甲酯(CGTCA-motif、TGACG-motif)響應(yīng)元件和脫落酸(ABRE)響應(yīng)元件;DlPIF1-1、DlPIF1-2、DlPIF3、DlPIF5、DlPIF7和DlPIF8含有生長素(TGA-element、AuxRR-core)響應(yīng)元件;DlPIF1-1、DlPIF3、DlPIF5和DlPIF7含有水楊酸(TCA-element)響應(yīng)元件;DlPIF3、DlPIF4、DlPIF5、DlPIF7還含有赤霉素(GARE-motif、P-box、TATC-box)響應(yīng)元件。
DlPIFs基因家族還含有種子特異性(RY-element)、胚乳特異性(GCN4_motif)、生物鐘(Circadian)、分生組織表達(CAT-box)、玉米蛋白代謝(O2-site)、α-淀粉酶等相關(guān)的作用元件和低溫脅迫(MBS)、防御和應(yīng)激(TC-rich repeats)、MYB轉(zhuǎn)錄因子結(jié)合位點相關(guān)的作用元件,并含有參與干旱誘導(dǎo)(MBS)、厭氧誘導(dǎo)(ARE)、缺氧特異性誘導(dǎo)(GC-motif)等與非生物脅迫相關(guān)的作用元件。
由上述結(jié)果推測,DlPIFs家族最主要的功能為光響應(yīng)功能,同時在激素響應(yīng)、生物鐘調(diào)控和非生物脅迫等方面起重要作用。
2.5 DlPIFs家族的系統(tǒng)進化樹分析
使用龍眼、擬南芥、水稻和甜橙4個物種的PIF蛋白序列構(gòu)建系統(tǒng)進化樹(圖5)。根據(jù)其進化親緣關(guān)系的遠近,將PIFs家族初步分為6大類(Class Ⅰ~VI),DlPIF6屬于 Class Ⅱ,DlPIF7和DlPIF8屬于 Class Ⅲ,DlPIF1-1和DlPIF4屬于Class V,DlPIF1-2、DlPIF3和DlPIF5屬于Class VI。DlPIFs家族與擬南芥和甜橙親緣關(guān)系較近。
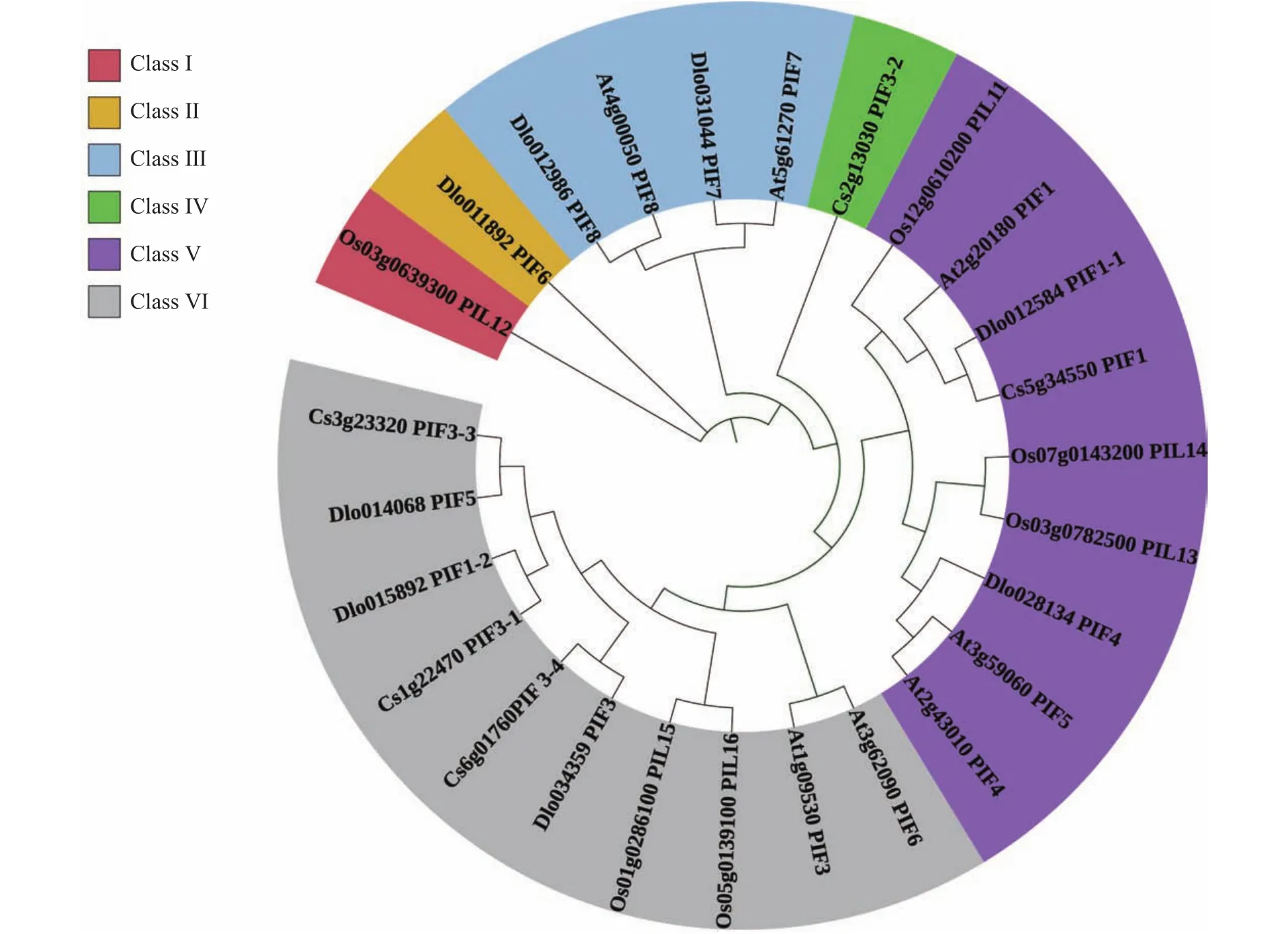
圖5 龍眼、擬南芥、水稻、甜橙PIFs家族系統(tǒng)進化樹Fig.5 Phylogenetic tree of PIF families in Arabidopsis thaliana, Oryza sativa L., Citrus sinensis, and D.longan
2.6 DlPIFs家族在不同組織器官的轉(zhuǎn)錄組基因表達水平分析
轉(zhuǎn)錄組基因表達水平結(jié)果(圖6)顯示,從整體上看DlPIFs在不同組織器官的表達水平在龍眼葉中最高,遠高于莖、花蕾、果肉、果皮、種子、幼果和花,在根中最低。DlPIF1-1的表達量在種子中最高,推測DlPIF1-1對種子的發(fā)育過程具有重要作用;DlPIF1-2的表達量在果肉中最高,推測DlPIF1-2對果肉的發(fā)育過程具有重要作用;DlPIF4的表達量在莖中最高,推測DlPIF4對莖的發(fā)育具有重要作用;DlPIF5的表達量在花蕾中最高,推測DlPIF5對花蕾的發(fā)育具有重要作用;DlPIF7和DlPIF8的表達量在葉中最高,推測DlPIF7和DlPIF8對葉的發(fā)育均具有重要作用。由上述結(jié)果可知,DlPIFs家族成員與龍眼不同組織器官的生長發(fā)育過程有著重要的聯(lián)系,在其中發(fā)揮著重要的功能。
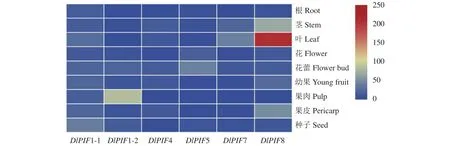
圖6 DlPIFs家族在非胚性和胚性培養(yǎng)物和不同組織器官的表達量熱圖Fig.6 Heat map of expressions of DlPIFs in non-embryogenic and embryogenic cultures and various tissues and organs
2.7 DlPIFs家族在不同光質(zhì)和不同溫度處理下的轉(zhuǎn)錄組基因表達水平分析
不同光質(zhì)處理下的轉(zhuǎn)錄組數(shù)據(jù)分析得出(圖7),除DlPIF7以外,其他DlPIFs家族成員均在不同光質(zhì)處理下有不同程度的表達;DlPIF1-1、DlPIF5和DlPIF8在藍光處理下的表達量明顯高于對照,推測DlPIF1-1、DlPIF5和DlPIF8能夠響應(yīng)藍光的調(diào)控;DlPIF4在白光和藍光處理下的表達量均明顯高于對照,推測DlPIF4能夠響應(yīng)藍光和白光的調(diào)控;DlPIF1-2在3種光質(zhì)處理下的表達量差別不明顯,推斷DlPIF1-2對不同光質(zhì)的響應(yīng)程度差別不明顯。DlPIFs成員在不同光質(zhì)處理下呈現(xiàn)不同的表達模式,推測DlPIFs成員能響應(yīng)光信號。
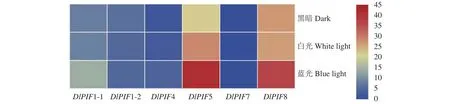
圖7 DlPIFs家族在不同光質(zhì)條件下的表達量熱圖Fig.7 Heat map of expressions of DlPIFs exposed to varied light
不同溫度處理下的轉(zhuǎn)錄組數(shù)據(jù)分析得出(圖8),相對高溫(35 ℃)促進DlPIF4和DlPIF6的表達,抑制DlPIF1-1、DlPIF1-2、DlPIF3和DlPIF8的表達;相對低溫(15 ℃)促進DlPIF1-1、DlPIF3和DlPIF5的表達,抑制DlPIF1-2、DlPIF4、DlPIF6和DlPIF8的表達。綜上所述,DlPIFs成員在不同溫度處理下有著不同的表達模式,說明DlPIFs大多數(shù)成員可能參與溫度的調(diào)控,并且對溫度的調(diào)控存在不同的作用。
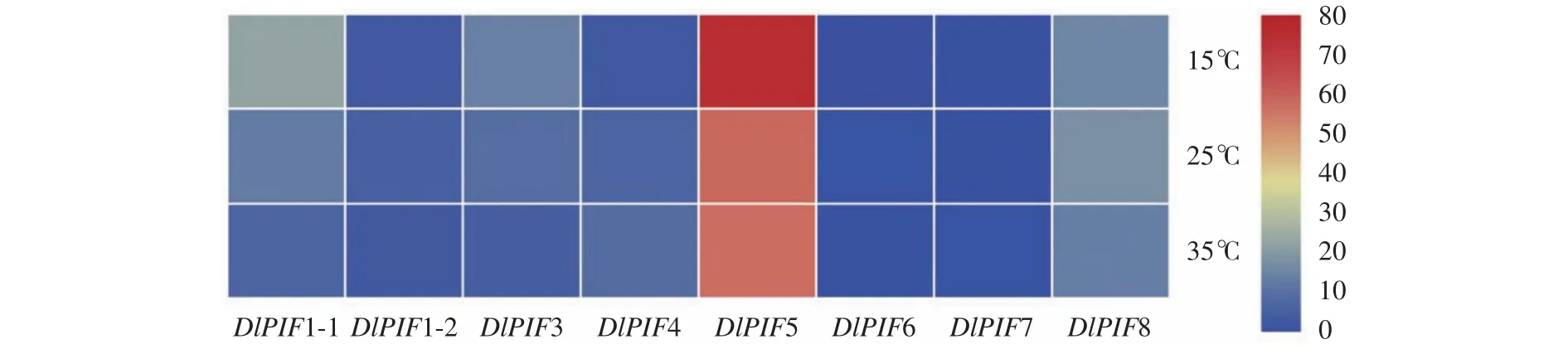
圖8 DlPIFs家族在不同溫度條件下的表達量熱圖Fig.8 Heat map of expressions of DlPIFs under different temperatures
2.8 DlPIFs家族在龍眼體胚發(fā)生早期的表達分析
龍眼體胚發(fā)生早期的qRT-PCR結(jié)果(圖9)顯示,DlPIF1-1、DlPIF5、DlPIF6和DlPIF8的表達量隨著龍眼體胚早期的發(fā)育不斷下降,推測DlPIF5、DlPIF6和DlPIF8DlPIF1-1、DlPIF5、DlPIF6和DlPIF8在龍眼體胚發(fā)育進程中具有相似的功能。DlPIF1-2和DlPIF3的表達量在EC到ICpEC階段下降,在ICpEC到GE階段上升,DlPIF4和DlPIF7的表達模式與之相反,推測DlPIF1-2和DlPIF3與DPIF4和DlPIF7對龍眼體胚發(fā)育進程具有相反的作用。綜上所述,DlPIFs家族不同成員在龍眼體胚發(fā)生早期具有不同的表達模式,可能通過相同或者不同的分子調(diào)控機制調(diào)節(jié)龍眼體胚的早期發(fā)育過程。
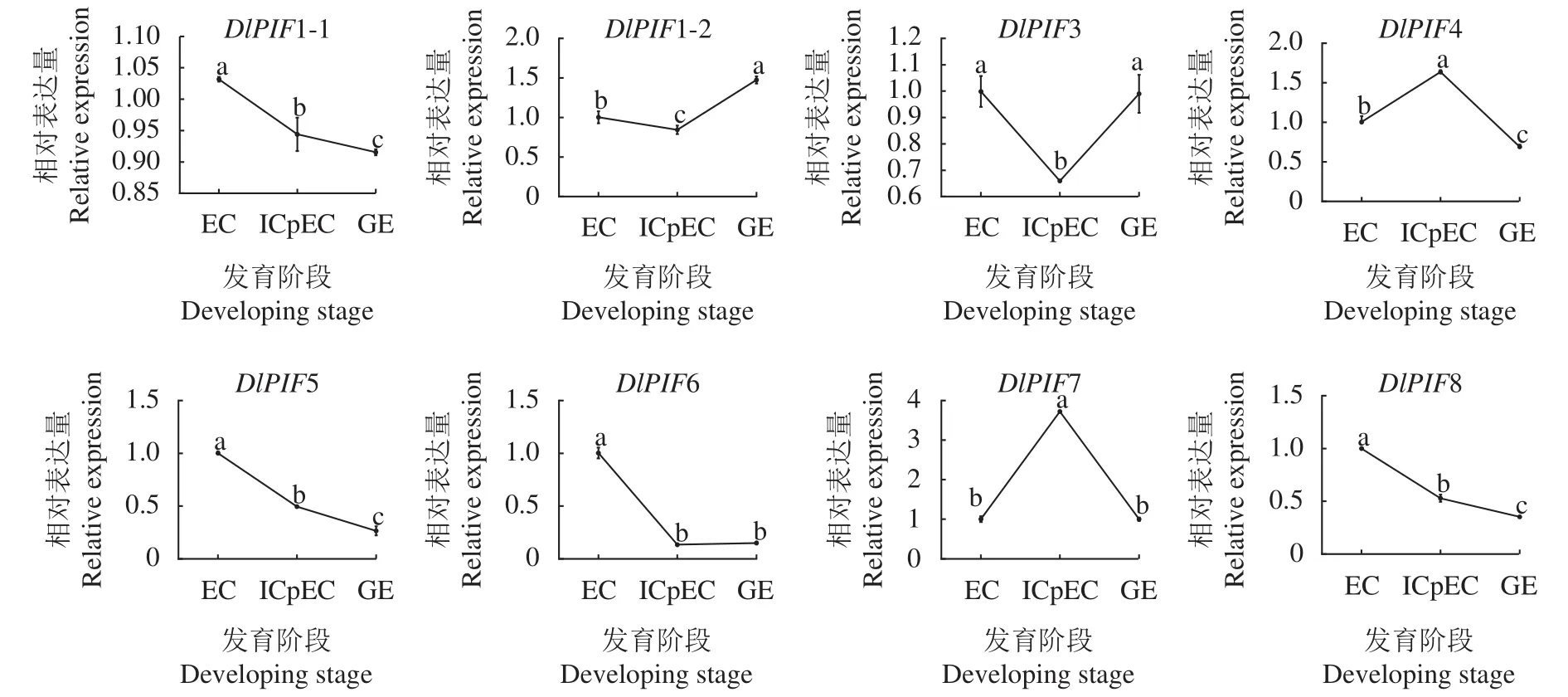
圖9 龍眼PIFs家族體胚發(fā)生階段表達分析Fig.9 Expressions of DlPIFs during somatic embryogenesis
2.9 DlPIFs家族在不同生長調(diào)節(jié)劑處理下的表達分析
利用qRT-PCR技術(shù)對不同生長調(diào)節(jié)劑處理下的龍眼愈傷組織中DlPIFs基因的表達水平進行分析(圖10),結(jié)果顯示,在GA3和ABA處理下,DlPIF3、DlPIF5和DlPIF7的表達量與對照相比有明顯提高,推測DlPIF3、DlPIF5和DlPIF7能響應(yīng)GA3和ABA的調(diào)控;2,4-D和SA處理下,DlPIF5和DlPIF7的表達量與對照相比有明顯提高,推測DlPIF5和DlPIF7能響應(yīng)2,4-D和SA的調(diào)控;在MeJA處理下,DlPIF1-2、DlPIF5和DlPIF7的表達量與對照相比有明顯提高,推測DlPIF1-2、DlPIF5和DlPIF7能響應(yīng)MeJA的調(diào)控。綜上所述,DlPIF5和DlPIF7的表達量在5種生長調(diào)節(jié)劑處理下都具有明顯變化,推測DlPIF5和DlPIF7是響應(yīng)激素調(diào)控的主要成員。

圖10 生長調(diào)節(jié)劑處理下DlPIFs基因的相對表達量Fig.10 Expressions of DlPIFs in response to exogenous hormones
顯著性差異分析發(fā)現(xiàn),在5種生長調(diào)節(jié)劑處理下,DlPIFs家族的大多數(shù)成員在不同時間處理下的表達量具有顯著性差異。
3 討論
3.1 DlPIFs家族的鑒定和功能的多樣性
近幾年,隨著對PIF基因家族研究的不斷深入,發(fā)現(xiàn)PIFs基因家族轉(zhuǎn)錄因子不僅在光信號中具有重要作用,在植物生長發(fā)育的多個方面都起到關(guān)鍵信號節(jié)點作用。目前研究發(fā)現(xiàn),擬南芥(Arabidopsis thaliana)中有7個PIFs家族成員(PIF1,PIF3~PIF8);水稻中有6個PIFs家族成員(OsPIL11~OsPIL16);玉米(Zea maysL.)中有7個PIFs家族成員(ZmPIF1~ZmPIF7)[31-32];香蕉(Musa nanaLour.)中有7個PIFs家族成員(MaPIF1、MaPIF1-1、MaPIF3、MaPIF3-1、MaPIF4、MaPIF4-1、MaPIF)[33];馬鈴薯(Solanum tuberosumL.)中有7個PIFs家族成員(St PIF1a、St PIF1b、StPIF3、St PIF4、St PIF7a、St PIF7b、St PIF8)[34];白菜(Brassica rapa)中有10個PIFs家族成員(BrPIF1-BrPIF10)[34]。本研究鑒定出龍眼PIFs家族8個具有bHLH結(jié)構(gòu)域的成員。基因結(jié)構(gòu)和保守基序分析發(fā)現(xiàn),部分成員之間具有相似的基因結(jié)構(gòu)和保守基序,推測成員的功能存在一定的保守性。亞細胞定位發(fā)現(xiàn)DlPIFs成員均位于細胞核,這與袁凌云等[35]在白菜和吳廣霞[36]在玉米中的研究一致。植物細胞內(nèi)擁有3套獨立的基因組,即細胞核基因組、線粒體基因組和葉綠體基因組。除去擁有巨大遺傳信息的細胞核之外,線粒體和葉綠體是植物細胞中具有雙膜系統(tǒng)的細胞器,它們的發(fā)育和增殖是受細胞核基因組及其自身基因兩套遺傳系統(tǒng)的控制,稱為半自主性細胞器[37]。染色體定位發(fā)現(xiàn),大部分DlPIFs成員定位于染色體上,而DlPIF3未定位于染色體,推測該基因為葉綠體或線粒體基因。啟動子順式作用元件分析有助于預(yù)測基因潛在的功能,進化樹和啟動子順式作用元件分析發(fā)現(xiàn),親緣關(guān)系較近的成員所含有的順式作用元件種類相似,不同成員之前包含的順式作用元件數(shù)量和種類有所差別,說明DlPIFs成員之間具有相同或者不同的生物學(xué)功能。
PIFs通過介導(dǎo)光信號,參與植物的光形態(tài)建成[38]。在擬南芥中PIF3負調(diào)控或者正調(diào)控植物幼苗的光反應(yīng)[6,39-40],PIF1、PIF4和PIF5可以負調(diào)控幼苗的各種光反應(yīng)[11,15,41],PIF7是紅光下幼苗去黃化的負調(diào)控因子[42],PIF6是植物光反應(yīng)的正調(diào)控因子[43]。與前人研究相同,本研究對DlPIFs家族進行順式作用元件分析得出DlPIFs家族各成員均含有與光響應(yīng)有關(guān)的作用元件且對不同光質(zhì)處理下DlPIFs轉(zhuǎn)錄組基因表達水平進行分析,發(fā)現(xiàn)DlPIFs成員普遍在藍光下表達量最高,在白光和黑暗處理下表達量較低。
PIFs不僅能夠響應(yīng)光信號,影響植物的光形態(tài)建成,還響應(yīng)非生物脅迫。前人研究表明光敏色素在參與調(diào)控植物對干旱脅迫的耐受性的過程中,phyB是主要的調(diào)控因子,在擬南芥中,PIF3和PIF4可能在phyB下游調(diào)控氣孔的開合中起作用[44];在玉米中,ZmPIFs均受到干旱脅迫的誘導(dǎo),而且其中起主要作用的是ZmPIF1和ZmPIF3[45]。本研究對DlPIFs家族進行順式作用元件分析得出PIF1-1、PIF1-2、PIF3、PIF4、PIF5、PIF6和PIF8含有與干旱響應(yīng)相關(guān)作用元件。有研究發(fā)現(xiàn)在高溫條件下,PIF4 能通過激活生長素的合成基因YUC8和TAA1促進生長素應(yīng)答基因IAA19和IAA29的表達來調(diào)節(jié)植物下胚軸和葉柄伸長[46],并有研究發(fā)現(xiàn)高溫下phyB可以促進HMR(phyB的互作蛋白)的積累,HMR進一步與PIF4相互作用,從而促進高溫響應(yīng)基因的表達[47]。低溫條件下,植物低溫脅迫響應(yīng)路徑的調(diào)控因子EBF1和 EBF2蛋白(能夠通過26S蛋白酶體直接靶向降解PIF3)降解使PIF3蛋白更加穩(wěn)定從而 抑制CBFs及其下游低溫響應(yīng)基因表達。此外,擬南芥PIF4和PIF7也能通過光周期依賴方式下調(diào)CBFs基因的表達來降低植物抵抗低溫的能力[48]。本研究發(fā)現(xiàn)DlPIFs家族中PIF1-1、PIF1-2、PIF5、PIF6和PIF8含有與低溫響應(yīng)相關(guān)的作用元件,并且對溫度處理的轉(zhuǎn)錄組數(shù)據(jù)進行分析得出,相對高溫(35 ℃)促進DlPIF4和DlPIF6的表達,抑制DlPIF1-1、DlPIF1-2、DlPIF3和DlPIF8的表達;相對低溫(15 ℃)促進DlPIF1-1、DlPIF3和DlPIF5的表達,抑制DlPIF1-2、DlPIF4、DlPIF6和DlPIF8的表達,此研究結(jié)果與前人研究相符。還有研究發(fā)現(xiàn)CCA1/LHY屬于MYB家族轉(zhuǎn)錄因子,是生物鐘的核心組件,PIFs能夠結(jié)合CCA1/LHY啟動子的G-box區(qū)域,參與生物鐘的調(diào)控[49]。本研究結(jié)果表明DlPIF1-1、DlPIF3和DlPIF4存在與生物鐘響應(yīng)相關(guān)的作用元件,與前人研究相符。
3.2 DlPIFs家族可能參與龍眼激素信號途徑
植物激素能以微小的含量調(diào)控植物的生命活動,是植物感受外部環(huán)境變化、調(diào)節(jié)自身生長狀態(tài)、抵御不良環(huán)境及維持生存必不可或缺的信號分子。在擬南芥中,脫落酸(ABA)激活PIF3和PIF6的表達,但抑制PIF1的表達,生長素(IAA)和茉莉酸甲酯(MeJA)分別抑制PIF5、PIF4和PIF8[8]。在玉米中,外源施加GA能顯著誘導(dǎo)ZmPIF5和ZmPIF6表達,與在ABA脅迫下的反應(yīng)相反,兩者可能作為ABA與GA信號的連結(jié)點[44]。有研究表明,在番茄(Lycopersicon esculentum)植株中,PIF4可以通過誘導(dǎo)JA和ABA合成基因的表達,從而促進JA和ABA積累,抑制GA合成基因的表達,降低GA含量[50]。本研究發(fā)現(xiàn),PIFs具有多種激素響應(yīng)元件,如生長素、赤霉素、脫落酸、水楊酸、茉莉酸甲酯,且對不同生長調(diào)節(jié)劑下表達量水平進行分析發(fā)現(xiàn),DlPIF5和DlPIF7的表達量在5種生長調(diào)節(jié)劑處理下都具有明顯變化,與前人研究相符。因此,推測DlPIFs參與龍眼激素信號途徑。
3.3 DlPIFs家族可能參與龍眼不同組織部位生長發(fā)育的調(diào)控
PIFs家族在不同植物的不同組織部位存在不同水平的表達。在擬南芥中,PIF1、PIF3和PIF4在幼苗和葉片中的表達量較高,PIF6在花和果實中的表達量較高,在嫩芽中,PIF5的表達量高于其他成員,PIF3在不同器官中的表達水平相似,而PIF8在所有地上器官中的表達水平相對較高[8]。在楊樹中,PtPIF1、PtPIF3和PtPIF8a在成熟葉中的表達水平較高,PtPIF3、PtPIF8a和PtPIF9b在幼葉中表達水平較高,PtPIF3b、PtPIF4、PtPIF5a、PtPIF9a和PtPIF10在各組織中的表達水平較低,PtPIF4和PtPIF5b只在成熟葉中表達[23]。與上述研究相似,DlPIFs成員在龍眼不同組織部位具有不同程度的表達水平,其中,DlPIF1-1在龍眼不同組織部位均具有較高的表達量,DlPIF4、DlPIF7和DlPIF8 在莖和葉中高表達,DlPIF5和DlPIF8 在花中高表達,在花蕾中6個DlPIFs成員的表達量均具有較高表達,DlPIF4和DlPIF8 在幼果和果皮中高表達,DlPIF1-2在果肉中高表達,DlPIF1-2在種子中有較高的表達。整體來看,在葉中的表達水平最高,在根中的表達水平最低,這與Gao等[32]在玉米中的發(fā)現(xiàn)一致。因此,表明DlPIFs是龍眼生長發(fā)育過程不可或缺的調(diào)控因子。
3.4 DlPIFs家族可能參與龍眼胚胎早期發(fā)育的調(diào)控
溫度和光照時間是影響棉花離體胚成苗的關(guān)鍵因素,適宜溫度和光照時間有利于離體胚的成苗和根系發(fā)育,溫度30 ℃、光照時間24 h的培養(yǎng)條件下,離體胚的水培成苗效果好,可作為早熟陸地棉離體胚成苗培養(yǎng)的適宜條件[51]。蜜瓜外植體培養(yǎng)中,在不同的光質(zhì)條件下愈傷組織的形成率有明顯的差異,其中,藍光對愈傷組織的形成有明顯的促進作用[52]。還有研究發(fā)現(xiàn),溫度對山核桃體胚發(fā)生具有一定影響,當(dāng)培養(yǎng)溫度在30 ℃時誘導(dǎo)率最高,當(dāng)培養(yǎng)溫度高于30 ℃或低于24 ℃時,誘導(dǎo)率均較低,并且出現(xiàn)較多畸形胚[53]。本研究得出,DlPIFs家族對溫度和光照等條件均有不同程度的表達,可能參與光照和溫度的調(diào)控,并且DlPIFs家族不同成員在龍眼體胚發(fā)生早期具有不同的表達模式。所以,推測DlPIFs家族可能通過不同的分子調(diào)控機制調(diào)節(jié)龍眼體胚的早期發(fā)育過程。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
