植物MYC1基因擬南芥表達載體構建及鑒定分析
2022-11-29 10:21:06耿慶鎏曹樹青
合肥工業大學學報(自然科學版) 2022年11期
關鍵詞:植物
耿慶鎏, 宋 慧, 胡 敏, 曹樹青
(合肥工業大學 食品與生物工程學院,安徽 合肥 230601)
0 引 言
植物在生長發育過程中,為適應復雜多變的自然環境進化出響應不同環境脅迫的生理代謝反應系統,這些系統使植物及時響應環境變化,保障植物的正常生長發育[1]。植物面對的環境脅迫分為生物脅迫(例如真菌、細菌感染,動物啃食)和非生物脅迫(例如寒冷、干旱、營養缺乏、重金屬毒害、土地鹽堿化)[2]。近年來,環境污染、氣候變化等問題日益突出,植物在生長發育過程中更容易受到環境脅迫影響,因此為保障人類糧食生產和食品安全,植物抗逆境研究尤為重要[3]。
鐵元素是土壤中含量最高的4種元素之一[4],作為植物需求量最大的微量元素,鐵元素是許多蛋白酶的組成成分,在植物的生理代謝反應(如光合作用、呼吸作用)中起氧化還原反應的功能[5]。植物缺鐵一般發生在pH>7的堿性或石灰質土壤上[6],由于堿性土壤條件下游離態的鐵元素減少,鐵元素難以被植物吸收,使植物出現缺鐵反應。植物缺鐵反應表現為因葉綠素減少引起的葉片失綠,嚴重時可影響植物生長發育,并且當植物中鐵元素過高也會導致植物體內產生過量活性氧造成細胞損傷甚至壞死,植物體內的鐵元素含量也受到嚴格控制[7],因此研究植物的鐵吸收及體內鐵穩態具有重要意義。
有研究表明,植物BHLH轉錄因子家族MYC1基因可能介導不同植物非生物脅迫反應[8],本研究以植物MYC1基因為研究對象,構建擬南芥ProMYC1-GUS植株材料,研究該基因在植物缺鐵脅迫中的響應機制,為今后的植物非生物脅迫研究和生物技術的應用提供理論基礎。
1 材料與方法
1.1 實驗材料
1.1.1 植物材料
本實驗所用植物材料為模式植物擬南芥(Arabidopsisthaliana),哥倫比亞生態型(Columbia,col),購于美國擬南芥種質資源中心(TAIR),由合肥工業大學植物逆境分子生物學實驗室培養繁殖。
1.1.2 質粒載體與菌株
重組載體構建所用質粒pART27-GUS、轉基因植物材料構建用菌株農桿菌GV3101、大腸桿菌DH5α,均購買于北京全式金生物技術有限公司。
1.1.3 試驗用試劑
Ps酶(TaKaRa);San Taq mix、壯觀霉素、慶大霉素、卡納霉素、瓊脂糖、酵母粉、蛋白胨、MES(生工生物工程(上海)股份有限公司);膠回收試劑盒、質粒小提試劑盒(TransGen);T4連接酶、限制性內切酶(New England Biolabs);MS(Murashige Skoog)培養基(Sangon Biotech);氯化鈉、瓊脂、蔗糖、氯化汞、CTAB、氯仿、異戊醇、無水乙醇、苯酚(中國醫藥集團有限公司);goldview、Silwet L77(北京索萊寶科技有限公司);GUSBlue組織染色試劑盒(北京華越洋生物科技有限公司)。
1.2 實驗方法
1.2.1 野生型擬南芥種植
營養土經120 ℃、20 min高溫滅菌后與珍珠鹽、蛭石以3∶1∶9比例混合拌勻,分裝至植物培養用花缽,花缽置于托盤中。向托盤加入適量營養液,營養土完全浸潤,每缽播種擬南芥種子5~7顆,花缽由保鮮膜包裹放置于植物培養室,22 ℃、16 h光照/8 h黑暗條件下培養,擬南芥發芽后3 d去掉保鮮膜。
1.2.2 重組載體構建
在TAIR網站(https://www.arabidopsis.org/)查詢擬南芥MYC1基因啟動子區域序列,通過Primer premier 5.0軟件設計該序列上下游引物并合成。上游引物為:
5’-GGGGTACCGAAACATTGTTATTTACTTTGGGACA-3’,
下游引物為:
5’-CCGCTCGAGATCTAGATATCGTGAAGAACTGAATT-3’。
限制性內切酶為KpnⅠ、XhoⅠ,由生工生物工程(上海)股份有限公司合成。
以擬南芥野生型植株基因組DNA為模板對其啟動子區域進行聚合酶鏈式反應(polymerase chain reaction,PCR)擴增,反應結束后,擴增片段進行瓊脂糖電泳并回收,回收片段與待連接空載質粒共同進行限制性內切酶雙酶切,37 ℃、2 h酶切后片段與質粒回收后16 ℃連接過夜。連接產物全部加入預先制備大腸桿菌DH5α感受態細胞,混勻,熱激法轉化,均勻涂布于壯觀霉素抗性篩選LB固體培養基,37 ℃培養過夜,挑選單菌落PCR鑒定,陽性克隆送測序(生工生物工程(上海)股份有限公司),測序序列與基因啟動子區域序列比對成功,重組載體構建成功。
1.2.3 電擊法農桿菌轉化
電擊杯冰上預冷,2 μL重組載體質粒加入預先制備農桿菌GV3101感受態細胞混勻,將感受態細胞轉移至電擊杯中,放入電轉儀。電轉儀參數設置為電壓1 800 V,電脈沖25 μF,啟動電脈沖,待電擊結束后電擊杯中菌液轉移至500 μL LB液體培養基,28 ℃、200 r/min振蕩培養4~6 h,均勻涂布于壯觀霉素和慶大霉素抗性篩選LB固體培養基,28 ℃培養48 h,挑選單菌落PCR鑒定,陽性克隆菌株甘油保菌-20 ℃保存待用。
1.2.4 浸花法擬南芥侵染
吸取100 μL已鑒定ProMYC1-GUS農桿菌菌液接種于3 mL壯觀霉素和慶大霉素抗性LB液體培養基,28 ℃、200 r/min振蕩培養活化。取2 mL菌液接種于100 mL壯觀霉素和慶大霉素抗性LB液體培養基,28 ℃、200 r/min振蕩培養至菌液OD600為1.0~1.3,4 000 r/min離心8 min,取上清液保留菌體,用侵染緩沖液重懸洗滌菌體,重復2次,最后用侵染緩沖液將菌液稀釋至OD600為0.5~0.6,并加入終體積分數為0.02%~0.03%的SilwetL-77,混勻待用。選取6周左右,生長發育正常野生型擬南芥植株,提前剪除果莢和已授粉花,花苞浸泡于菌液15~20 s,所有植株完全浸潤后保鮮膜包裹并黑暗處理過夜,除去保鮮膜。1周后再次侵染,2周左右收種,烘干后4 ℃春化待用。
1.2.5 轉基因陽性植株篩選
配制1/2MS固體培養基,高溫滅菌(120 ℃,20 min)后待其冷卻至50~60 ℃,加入終質量濃度為50 μg/mL卡納霉素,混勻后倒入滅菌培養皿,冷卻待用。超凈臺內0.1%氯化汞溶液洗滌侵染植株子代種子1次,無菌水洗滌3次。將洗滌后種子均勻灑在卡納霉素1/2MS固體培養基,封口膜密封后4 ℃春化3 d,光照培養14 d,選取體型較大,葉子較綠,根系較深植株轉移至營養土繼續生長,2周后提取基因組DNA,PCR鑒定,去除鑒定陰性植株,4周后單株收種,子代種子點在卡納霉素1/2MS固體培養基進行抗性分離,篩選純合體。
1.2.6 GUS染色
將已獲得的ProMYC1-GUS陽性植株純合體種子點在MS固體培養基,4 ℃春化3 d,光照培養7 d后將萌發幼苗轉移至MS固體培養基和缺鐵MS固體培養基,分為對照組和實驗組。培養3 d后用GUS染色液將ProMYC1-GUS陽性植株幼苗染色處理,37 ℃、6 h,75%乙醇溶液脫色后觀察。
2 結果與分析
2.1 目的啟動子區域片段克隆
為獲得目的基因啟動子區域DNA片段,以野生型擬南芥基因組DNA為模板,PCR擴增(95 ℃,5 min預變性;95 ℃,30 s變性,55 ℃,30 s退火,72 ℃,2 min延伸,34個循環;72 ℃,5 min,50μL體系),瓊脂糖電泳檢測結果如圖1所示。
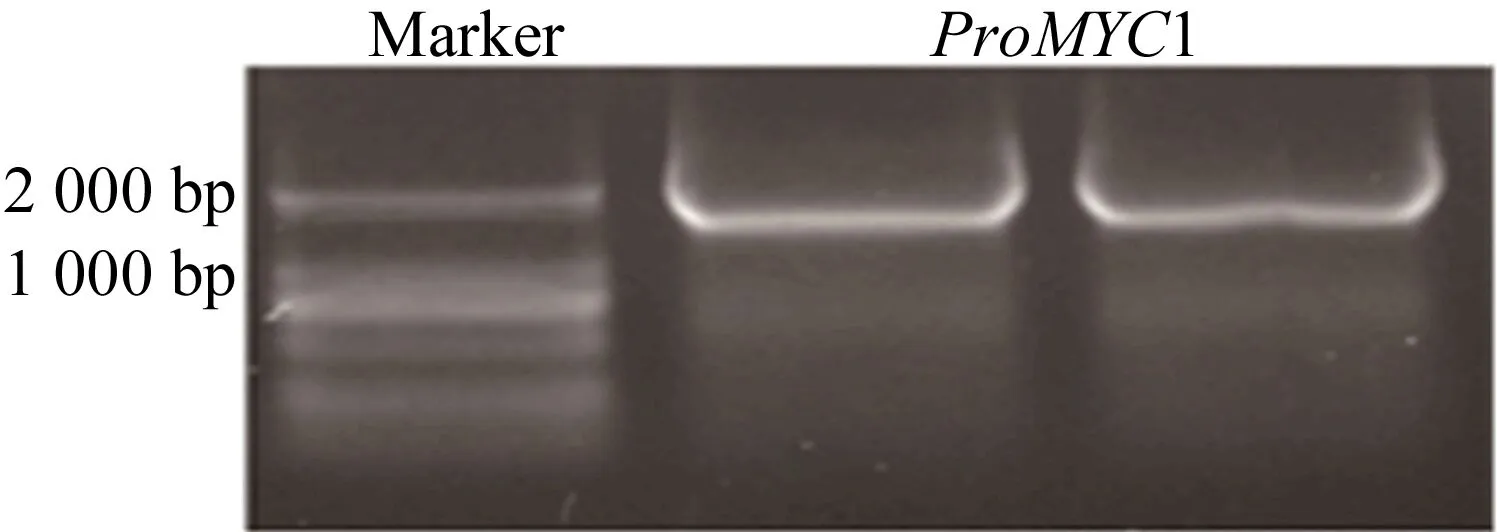
圖1 擬南芥MYC1基因啟動子片段的克隆
由圖1可知,獲得的目的片段大小在2 000 bp左右,與引物設計預期片段大小相同。
2.2 目的片段與質粒雙酶切連接
34 μL目的片段回收產物與pART27-GUS質粒置于1.5 mL離心管中加入4 μL CutSmart緩沖液,KpnⅠ和XhoⅠ各添加1 μL,混勻離心后37 ℃金屬浴酶切2 h。酶切后片段和質粒瓊脂糖電泳結果如圖2所示。
從圖2可以看出,酶切后條帶清晰,大小正常,未見拖帶、彌散、分層等異常結果,回收酶切片段與質粒。使用T4-DNA連接酶連接體系(酶切片段6.5 μL,酶切質粒2 μL,T4-DNA連接酶緩沖液1 μL,T4-DNA連接酶0.5 μL)連接回收產物,混勻離心后16 ℃金屬浴連接16 h,獲得連接產物。
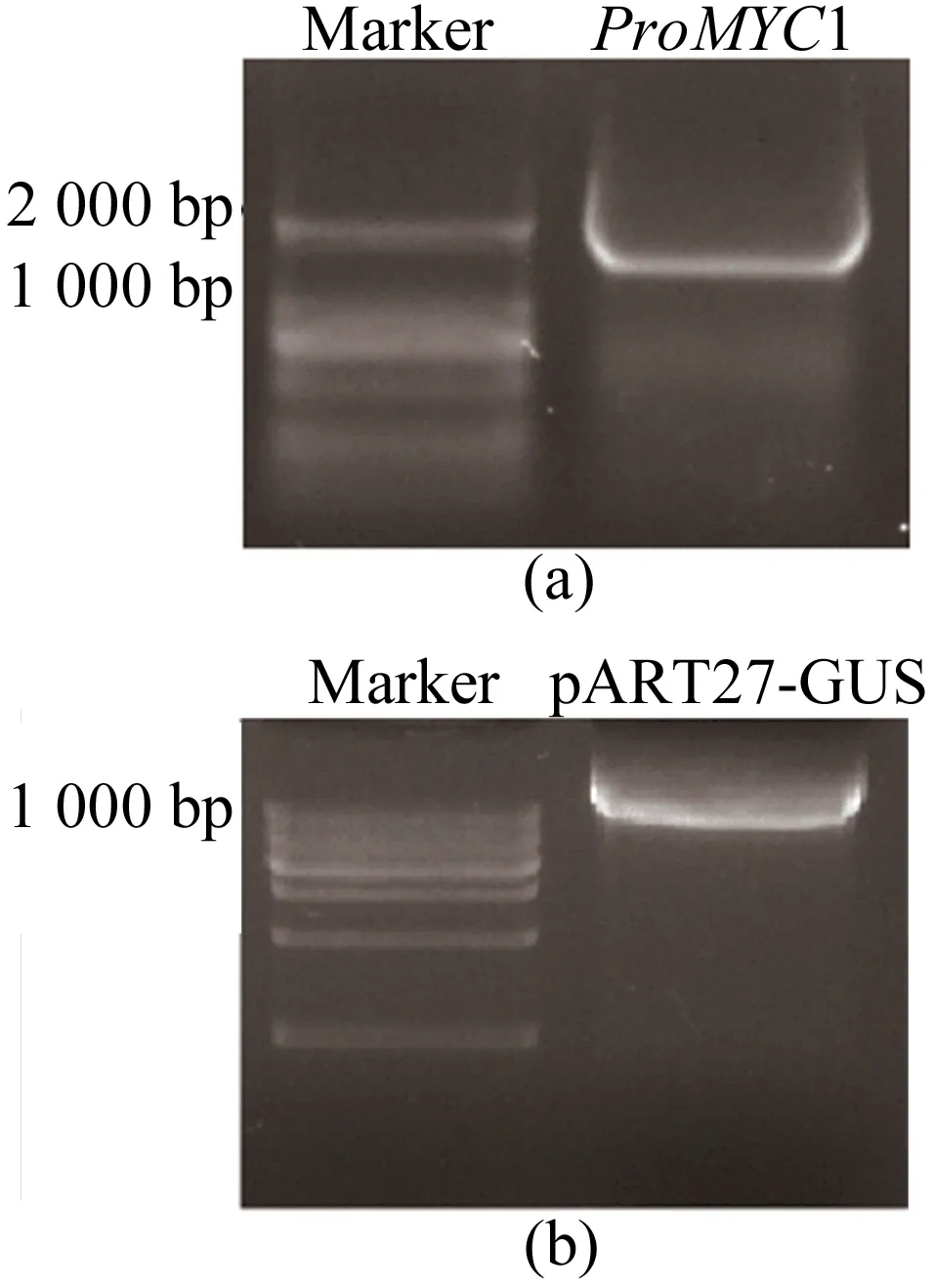
圖2 啟動子片段與質粒雙酶切
2.3 重組載體大腸桿菌轉化和陽性克隆鑒定
將10 μL連接產物加入50 μL預先制備的大腸桿菌DH5α感受態細胞,冰浴30 min,42 ℃熱激1 min,再冰浴2 min,加入500 μL無抗性LB液體培養基,37 ℃、200 r/min振蕩培養1 h,均勻涂布于壯觀霉素抗性篩選LB固體培養基,37 ℃培養過夜后挑取單菌落培養并進行PCR鑒定,瓊脂糖電泳結果如圖3所示,圖3中,1~12代表大腸桿菌的PCR產物。

圖3 大腸桿菌菌落PCR驗證
從圖3可以看出,2~7號泳道有明顯亮帶,并且與Marker比對,條帶大小與目的片段大小一致,選取2號、3號單菌落進行測序并于目的片段序列比對,2號、3號為陽性克隆。
2.4 重組載體農桿菌轉化與侵染
將5 μL 2號大腸桿菌測序質粒加入50 μL預先制備的農桿菌GV3101感受態細胞,電擊法轉化后挑取單菌落進行PCR鑒定,瓊脂糖電泳結果如圖4所示,圖4中,1~12代表農桿菌的PCR產物。
從圖4可以看出,所有泳道均有明顯亮帶,大小正確,均為陽性克隆農桿菌單菌落。選取1號農桿菌單菌落接菌,侵染4盆,共計20棵擬南芥野生型植株。

圖4 農桿菌菌落PCR驗證
2.5 ProMYC1-GUS陽性植株篩選與鑒定
浸花法侵染擬南芥野生型植株子代種子,灑在卡納抗生素抗性1/2MS固體培養基光照培養,2周后觀察種子萌發生長發育情況,如圖5所示,圖5中框選植株生長發育較為正常,移至營養土繼續生長,2周后提取植株基因組DNA進行PCR鑒定。

圖5 轉基因陽性植株抗性篩選
第1代轉基因植株成熟收種后繼續播種,子代植株分株單獨收種,第3代子代種子每種取20~25顆點在卡納抗生素抗性1/2MS固體培養基上,光照培養2周后觀察種子萌發生長發育情況如圖6所示。
由圖6可知,3組正常生長發育,而1、2、4組部分幼苗發黃,由遺傳分離定律可知,3組為純和陽性轉基因組。

圖6 轉基因植株抗性分離篩選
2.6 缺鐵脅迫轉基因材料GUS染色
為探究植物MYC1基因在缺鐵脅迫下基因表達的變化對植物缺鐵的響應,MS培養基正常生長7 d后ProMYC1-GUS轉基因植株移至MS和缺鐵MS固體培養基,3 d后對其GUS染色,通過觀察顏色反應的GUS基因表達量的不同,分析MYC1基因在缺鐵脅迫下轉錄水平表達量的變化。GUS染色結果如圖7所示,從圖7可以看出,缺鐵誘導條件下ProMYC1-GUS轉基因植株相較于對照組染色程度明顯降低,說明植物MYC1基因在缺鐵脅迫下表達被抑制,表達量明顯下降,該基因可能作為負調控因子介導植物缺鐵脅迫反應。

圖7 ProMYC1-GUS轉基因植株GUS染色
3 結 論
由于植物在自然界中和人類生產活動中扮演著非常重要且不可替代的角色,植物脅迫響應的研究與人類生活息息相關,通過對植物脅迫響應基因的功能研究幫助人類揭示植物逆境響應反應網絡和信號通路,并且以此為基礎解決環境保護、生態治理以及農業生產上的實際問題。
GUS基因作為一種常見的報告基因通過水解底物產生的顏色反應來指示目的基因轉錄水平,由于其幾乎不存在于高等植物中,并且表達穩定、結果分析簡單,經常作為一種融合基因標記來檢測植物基因在組織中的表達量[9]。有報道指出,擬南芥MYC1基因介導植物激素信號調控的非生物脅迫生理過程[10]。本研究通過構建擬南芥ProMYC1-GUS轉基因材料和缺鐵誘導條件下GUS染色,分析該基因誘導前后表達量的變化,結果表明MYC1基因在缺鐵條件下被抑制表達,進一步驗證了該基因與植物非生物脅迫(缺鐵脅迫)有關,為MYC1基因介導非生物脅迫的進一步研究打下基礎。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
