自噬在脂多糖誘導的A549細胞炎癥反應中的作用及機制研究
2022-10-18 14:06:18時佳陶慧嫻郭艷鄒蕓蘇王木子盧志濤丁溢芳許衛東周曉光
中國當代兒科雜志 2022年10期
時佳 陶慧嫻 郭艷 鄒蕓蘇 王木子 盧志濤 丁溢芳 許衛東 周曉光
(1.南京醫科大學附屬兒童醫院新生兒醫療中心,江蘇南京 210008;2.張家港市第一人民醫院新生兒科,江蘇張家港 215600)
急性呼吸窘迫綜合征(acute respiratory distress syndrome,ARDS)是機體在遭受各種病理刺激(如缺氧、感染、休克等)后發生的一種急性肺損傷(acute lung injury,ALI),是新生兒臨床常見的危重癥。其特征是肺泡-毛細血管膜屏障遭到破壞,從而增強炎癥細胞的遷移能力和促炎細胞因子的產生,使炎癥反應失控[1]。迄今為止,ARDS的發病率和病死率一直居高不下。因此,迫切需要闡明新生兒ARDS的發病機制以制訂相應的防治措施。
自噬是一種古老的、進化保守的細胞內消化和再循環途徑,主要包括巨自噬、微自噬和分子伴侶介導的自噬3種類型,對于維持細胞的正常功能及細胞穩態具有重要作用。巨自噬以其處理大型細胞內結構的能力著稱,雙膜囊泡結構的自噬體也成為其區別于另外兩種自噬形式的特征。自噬參與人類多種疾病的發生、發展過程,本研究主要對巨自噬(以下簡稱“自噬”)進行研究。自噬被認為是應對炎癥的一種基本細胞反應,其相關途徑(如微管相關蛋白輕鏈3相關吞噬作用和非常規分泌等)是免疫和炎癥的核心穩態機制[2]。一般來說,自噬有助于宿主激活免疫以控制感染,同時限制有害的、不受控制的炎癥反應。在先天免疫系統中,Toll 樣受體4(Toll-like receptor 4,TLR4) 是一種重要的模式識別受體(pattern recognition receptor,PRR),PRR 通過識別病原體相關分子模式或危險相關分子模式,調節先天免疫或炎癥過程[3]。TLR4 也是第1 個被證明參與自噬 過 程 的 受 體[4]。 TLR4 經 脂 多 糖(lipopolysaccharide,LPS)激活后產生的細胞因子參與病原體的清除,對機體有益;但當該過程失調時,可能會導致危及生命的病理生理改變,如新生兒ARDS[5]。因此,TLR4 被認為是一個重要的藥理學靶點。目前有關自噬與新生兒ARDS發病關系的研究報道極少,自噬在新生兒ARDS炎癥反應所致肺損傷中的作用與機制尚不明了。本實驗利用LPS 誘導人肺泡上皮A549 細胞(簡稱“A549細胞”)炎癥反應來模擬新生兒ARDS發病過程中失控的炎癥反應與肺損傷變化,旨在探討自噬在炎癥反應導致肺損傷過程中的作用及其機制,為新生兒ARDS的防治提供新的途徑和實驗依據。
1 材料與方法
1.1 細胞和主要試劑
人肺泡上皮細胞株A549(上海中喬新舟生物科技有限公司);DMEM 高糖培養基(Gibco,美國);磷酸緩沖鹽溶液、內參β-actin(武漢賽維爾生物科技有限公司);胎牛血清(PAN,德國);LPS、兔抗LC3B、P62 抗體(Sigma,美國);兔抗Beclin-1 抗體(Abcam,英 國);兔抗NLRP3、TLR4抗體(北京博奧森生物技術有限公司);兔抗Caspase-1、ASC 抗體、內參GAPDH(武漢三鷹生物技術有限公司);辣根過氧化物酶標記山羊抗兔IgG抗體(南京巴傲得生物科技有限公司);3-甲基腺嘌呤(3-methyladenine,3-MA)、雷帕霉素(rapamycin,RAPA)(MCE,美國);CCK-8 試劑盒 (Dojindo, 日 本); LipofectamineTM2000(Invitrogen Life Technologies,美國);人源TLR4 過表達載體質粒、TLR4 siRNA(均由南京銳博生物科技有限公司設計);RIPA裂解液(上海碧云天生物技術有限公司)。
1.2 A549細胞培養及實驗分組
向人肺泡上皮細胞株A549 細胞中施加含10%胎牛血清的DMEM 高糖培養基,置于37℃、5%CO2的培養箱中培養。當細胞密度長至90%后棄培養液,磷酸緩沖鹽溶液清洗2 次,以0.25%EDTA 胰酶消化細胞,調整細胞密度接種于細胞培養板中,培養至細胞穩定貼壁。
(1)濃度梯度分組:加入不同濃度(0、1、5、10 μg/mL)的LPS 工作液,刺激12 h;時間梯度分組:加入濃度為1 μg/mL 的LPS 工作液,刺激不同的時間(0、4、8、12、24 h)。收樣前0.5 h加入三磷酸腺苷(adenosine triphosphate,ATP)(濃度為5 mmol/L)。收集各組細胞沉淀,用于后續蛋白檢測。
(2)細胞自噬抑制實驗分為:對照組、LPS組、3-MA組、3-MA+LPS組。3-MA組和3-MA+LPS組加入濃度為5 mmol/L 的自噬抑制劑3-MA 與細胞共培養12 h。細胞自噬激動實驗分為:對照組、LPS 組、RAPA 組、RAPA+LPS 組。RAPA 組 和RAPA+LPS 組加入濃度為100 nmol/L 的自噬激動劑RAPA 與細胞共培養12 h。LPS 的處理濃度為1 μg/mL,處理時間為12 h。收樣前0.5 h加入ATP。收集各組細胞沉淀,用于后續蛋白檢測。
(3)細胞轉染實驗分兩部分,每部分各分4組:TLR4 過表達對照組、TLR4 過表達組、TLR4過表達對照+LPS 組、TLR4 過表達+LPS 組;TLR4沉默對照組、TLR4 沉默組、TLR4 沉默對照+LPS組、TLR4沉默+LPS組。
1.3 細胞轉染
將細胞接種至6 孔板中,待細胞密度長至40%~60%時進行轉染。轉染前2 h 對細胞進行換液。通過構建質粒載體過表達TLR4,使用siRNA沉默TLR4,針對質粒和siRNA 進行轉染工作液的配制:將4 μg 的TLR4 過表達/空載質粒DNA 和10 μL 濃 度 為20 μmol/L 的TLR4 siRNA 及 其 對 照siRNA 加至250 μL 的無血清培養基中,混勻,再將5~10 μL 的LipofectamineTM2000 加至另外250 μL的無血清培養基中,混勻,室溫靜置5 min,再將兩者混合,室溫靜置20 min。從每孔中吸出500 μL培養基棄去,再按每孔500 μL 將上述配制的混合液加入,混勻,4~6 h 后換液,轉染48 h。收樣前0.5 h 加入ATP(濃度為5 mmol/L)。收集各組細胞沉淀,用于后續蛋白檢測。
1.4 CCK-8法檢測細胞存活率
將細胞接種于96孔板中培養12~24 h。以濃度為0、1、5、10 μg/mL 的LPS 刺激12 h;以濃度為1 μg/mL的LPS分別刺激0、4、8、12、24 h,收樣前0.5 h 加入ATP,后每孔加入10 μL 的CCK-8 溶液,1~4 h 后應用酶標儀檢測450 nm 處各孔吸光度。實驗獨立重復3次。
1.5 Western blot法檢測蛋白表達
采用RIPA 裂解液提取細胞總蛋白,并經二喹啉甲酸法測定蛋白質濃度。通過SDS-PAGE電泳分離蛋白后轉移到PVDF 膜上,5%脫脂牛奶室溫封閉2 h,分別孵育相應抗體:微管相關蛋白1 輕鏈3B (microtubule associated protein 1 light chain 3 beta, MAP1LC3B, LC3B) 、 Beclin-1、 P62、NLRP3、Caspase-1、ASC、TLR4(均為1∶1 000配制),4℃過夜。TBST 洗膜(10 min×3 次),室溫 孵 育 二 抗 (1∶5 000) 1 h, TBST 洗 膜(10 min×3 次),用ECL 發光劑熒光顯色,在化學發光凝膠成像系統曝光觀察蛋白條帶。以GAPDH(1∶5 000)或β-actin(1∶1 000)作為內參,分析各條帶灰度值。實驗獨立重復3次。
1.6 統計學分析
采用GraphPad Prism 8.0 進行分析和統計。正態分布的計量資料以均數±標準差(±s)表示,兩組間比較采用兩樣本t檢驗,多組間比較采用方差分析,組間兩兩比較采用LSD-t法。P<0.05為差異有統計學意義。
2 結果
2.1 LPS刺激對A549細胞存活率的影響
在培養的A549 細胞中以不同濃度(0、1、5、10 μg/mL) 的LPS 刺 激12 h, 與0 μg/mL 組(100.0%±0.0%) 相 比, 1 μg/mL 組(86.0%±1.2%),5 μg/mL組(72.8%±1.9%)和10 μg/mL組(63.6%±1.0%)的細胞存活率隨LPS 濃度的增加逐漸降低;以1 μg/mL的LPS在A549細胞中刺激不同 的 時 間(0、4、8、12、24 h),與0 h 組(100.0%±0.0%) 相比,細胞存活率在4 h 組(97.67%±0.23%)、8 h 組(94.21%±0.65%) 及12 h 組(95.15%±0.59%)僅有輕微降低,24 h 組(72.33%±1.59%)則明顯降低,差異均有統計學意義(P<0.05),見圖1。
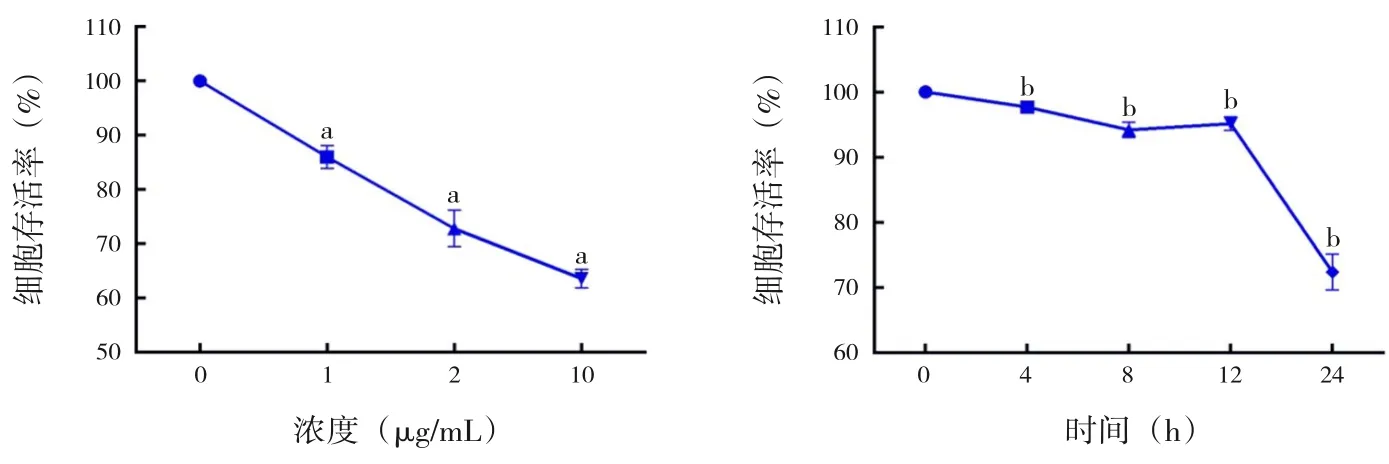
圖1 不同LPS 的刺激濃度和時間對A549 細胞存活率的影響
2.2 LPS對A549細胞炎癥水平的影響
在培養的A549 細胞中以不同濃度(0、1、5、10 μg/mL) 的LPS 刺 激12 h,炎 癥 指 標NLRP3、Caspase-1和ASC蛋白表達水平在1 μg/mL組升高并達到高峰,與0 μg/mL組比較,差異均有統計學意義(P<0.05);以1 μg/mL 的LPS 在A549 細胞中刺激不同的時間(0、4、8、12、24 h),炎癥指標NLRP3、Caspase-1 和ASC 蛋白表達水平均在12 h組升高并達到高峰,與0 h組比較,差異均有統計學意義(P<0.05),見圖2及表1~2。
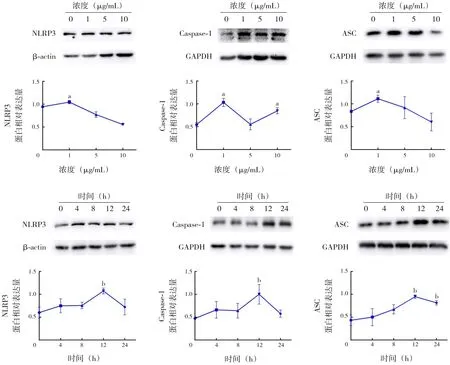
圖2 LPS對A549細胞炎癥蛋白表達的影響
2.3 LPS對A549細胞自噬水平的影響
在培養的A549 細胞中以不同濃度(0、1、5、10 μg/mL)的LPS 刺激12 h,自噬相關蛋白LC3B-Ⅱ/Ⅰ、Beclin-1 和P62 的表達水平均在1 μg/mL 組升高并達到高峰,與0 μg/mL組比較,差異均有統計學意義(P<0.05);以1 μg/mL 的LPS 在A549 細胞中刺激不同的時間(0、4、8、12、24 h),自噬相關蛋白LC3B-Ⅱ/Ⅰ、Beclin-1 和P62 的表達水平均在12 h組升高并達到高峰,與0 h組比較,差異均有統計學意義(P<0.05),見圖3及表1~2。
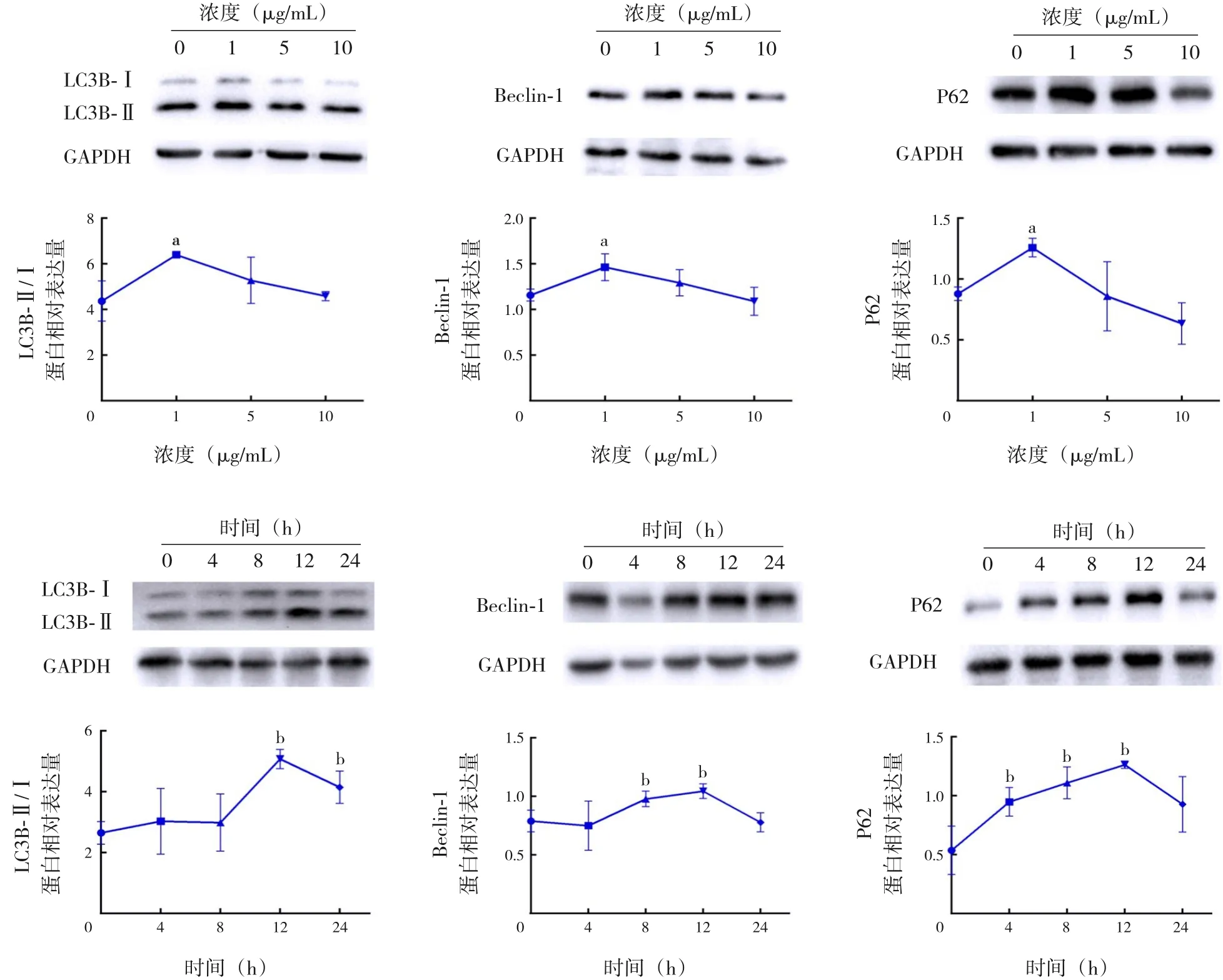
圖3 LPS對A549細胞自噬相關蛋白表達的影響
表1 不同濃度LPS刺激各組炎癥及自噬相關蛋白表達的比較 (±s,n=3)
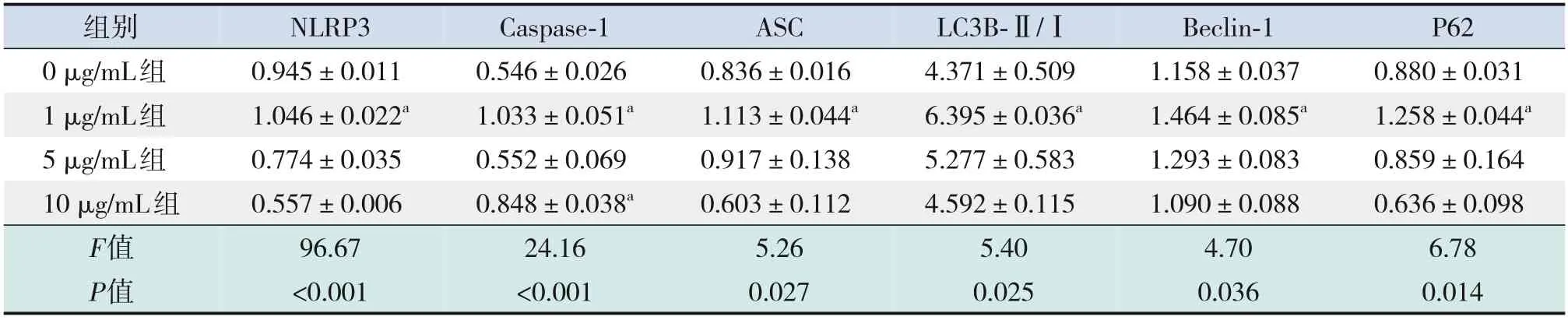
表1 不同濃度LPS刺激各組炎癥及自噬相關蛋白表達的比較 (±s,n=3)
注:a示與0 μg/mL組比較,P<0.05。
組別NLRP3 Caspase-1 ASC LC3B-Ⅱ/ⅠBeclin-1 P62 0 μg/mL組1 μg/mL組5 μg/mL組10 μg/mL組F值P值0.557±0.006 96.67<0.001 0.848±0.038a 24.16<0.001 0.603±0.112 5.26 0.027 4.592±0.115 5.40 0.025 1.090±0.088 4.70 0.036 0.636±0.098 6.78 0.014 0.945±0.011 1.046±0.022a 0.774±0.035 0.546±0.026 1.033±0.051a 0.552±0.069 0.836±0.016 1.113±0.044a 0.917±0.138 4.371±0.509 6.395±0.036a 5.277±0.583 1.158±0.037 1.464±0.085a 1.293±0.083 0.880±0.031 1.258±0.044a 0.859±0.164
表2 LPS刺激不同時間各組炎癥及自噬相關蛋白表達的比較 (±s,n=3)

表2 LPS刺激不同時間各組炎癥及自噬相關蛋白表達的比較 (±s,n=3)
注:a示與0 h組比較,P<0.05。
組別NLRP3 Caspase-1 ASC LC3B-Ⅱ/ⅠBeclin-1 P62 0 h組4 h組8 h組12 h組24 h組F值P值6.07 0.010 5.37 0.014 10.92 0.001 5.95 0.010 3.97 0.035 8.41 0.003 0.60±0.07 0.75±0.09 0.76±0.04 1.07±0.04a 0.72±0.10 0.475±0.005 0.661±0.105 0.643±0.092 1.002±0.123a 0.577±0.046 0.435±0.066 0.502±0.109 0.672±0.058 0.949±0.022a 0.815±0.028a 2.653±0.213 3.033±0.624 2.986±0.543 5.079±0.182a 4.146±0.306a 0.789±0.053 0.750±0.121 0.978±0.038a 1.045±0.036a 0.779±0.047??0.537±0.119 0.949±0.070a 1.110±0.077a 1.263±0.018a 0.928±0.136
2.4 LPS對A549細胞TLR4蛋白表達水平的影響
在培養的細胞中以不同濃度(0、1、5、10 μg/mL)的LPS 刺激12 h,TLR4 蛋白表達水平在1 μg/mL 組(0.585±0.054)升高并達到高峰,與0 μg/mL 組比較,差異有統計學意義(P<0.05);以1 μg/mL 的LPS 在A549 細胞中刺激不同的時間(0、4、8、12、24 h),TLR4 蛋白表達水平在12 h組(1.090±0.031)升高并達到高峰,與0 h 組比較,差異有統計學意義(P<0.05),見圖4。

圖4 LPS對A549細胞TLR4蛋白表達的影響
2.5 自噬對炎癥的影響
2.5.1 自噬抑制劑3-MA對炎癥及自噬相關蛋白表達水平的影響 在自噬抑制劑3-MA 的處理下,自噬相關蛋白LC3B-Ⅱ/Ⅰ蛋白表達水平明顯降低,P62 蛋白表達水平明顯升高;而炎癥指標NLRP3、Caspase-1、ASC蛋白表達水平明顯升高,與對照組比較,差異均具有統計學意義(P<0.05)。經LPS(1 μg/mL,12 h)刺激后,3-MA+LPS 組自噬相關蛋白LC3B-Ⅱ/Ⅰ表達仍呈降低趨勢,P62蛋白表達進一步升高;炎癥指標NLRP3、Caspase-1、ASC蛋白表達水平則進一步升高,與LPS組比較,差異均具有統計學意義(P<0.05),見圖5和表3。
表3 自噬抑制劑3-MA干預后炎癥及自噬蛋白表達的比較 (±s,n=3)
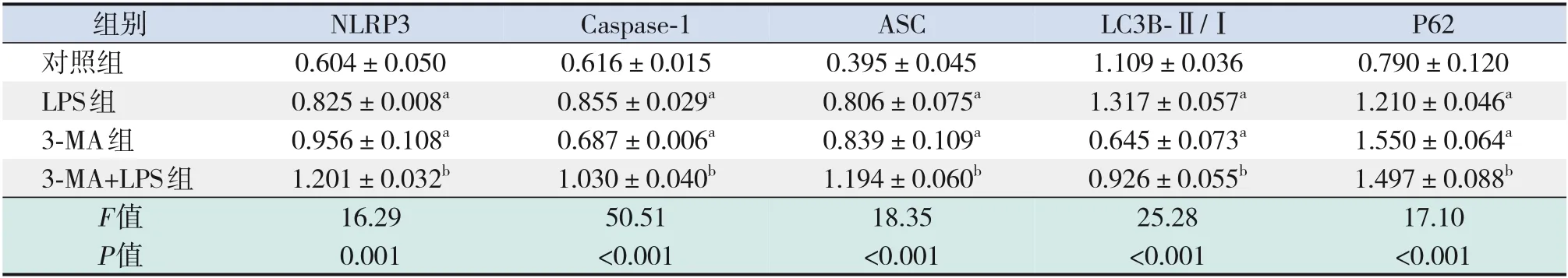
表3 自噬抑制劑3-MA干預后炎癥及自噬蛋白表達的比較 (±s,n=3)
注:a示與對照組比較,P<0.05;b示與LPS組比較,P<0.05。[LPS]脂多糖;[3-MA]3-甲基腺嘌呤。
組別對照組LPS組3-MA組3-MA+LPS組F值P值NLRP3 0.604±0.050 0.825±0.008a 0.956±0.108a 1.201±0.032b 16.29 0.001 Caspase-1 0.616±0.015 0.855±0.029a 0.687±0.006a 1.030±0.040b 50.51<0.001 ASC 0.395±0.045 0.806±0.075a 0.839±0.109a 1.194±0.060b 18.35<0.001 LC3B-Ⅱ/Ⅰ1.109±0.036 1.317±0.057a 0.645±0.073a 0.926±0.055b 25.28<0.001 P62 0.790±0.120 1.210±0.046a 1.550±0.064a 1.497±0.088b 17.10<0.001
2.5.2 自噬激動劑RAPA對炎癥及自噬相關蛋白表達水平的影響 在自噬激動劑RAPA 的處理下,自噬相關蛋白LC3B-Ⅱ/Ⅰ表達水平明顯升高,P62蛋白表達水平明顯降低;而炎癥指標NLRP3、Caspase-1、ASC蛋白表達水平明顯降低,與對照組比較,差異均具有統計學意義(P<0.05)。經LPS(1 μg/mL,12 h)刺激后,RAPA+LPS 組自噬相關蛋白LC3B-Ⅱ/Ⅰ表達進一步升高,P62蛋白表達則仍然是降低的;炎癥指標NLRP3、Caspase-1、ASC蛋白表達水平仍呈降低趨勢,與LPS組比較,差異均具有統計學意義(P<0.05),見圖5和表4。
表4 自噬激動劑RAPA干預后炎癥及自噬蛋白表達的比較 (±s,n=3)

表4 自噬激動劑RAPA干預后炎癥及自噬蛋白表達的比較 (±s,n=3)
注:a示與對照組比較,P<0.05;b示與LPS組比較,P<0.05。[LPS]脂多糖;[RAPA]雷帕霉素。
組別對照組LPS組RAPA組RAPA+LPS組F值P值NLRP3 0.525±0.057 0.735±0.022a 0.143±0.017a 0.358±0.013b 59.26<0.0001 Caspase-1 0.795±0.031 0.948±0.018a 0.570±0.075a 0.573±0.085b 9.61 0.005 ASC 0.722±0.027 0.944±0.027a 0.247±0.038a 0.349±0.065b 58.79<0.0001 LC3B-Ⅱ/Ⅰ2.111±0.078 4.845±0.446a 11.700±0.507a 8.001±0.621b 80.56<0.0001 P62 0.906±0.026 1.066±0.029a 0.432±0.109a 0.452±0.142b 12.29 0.0023
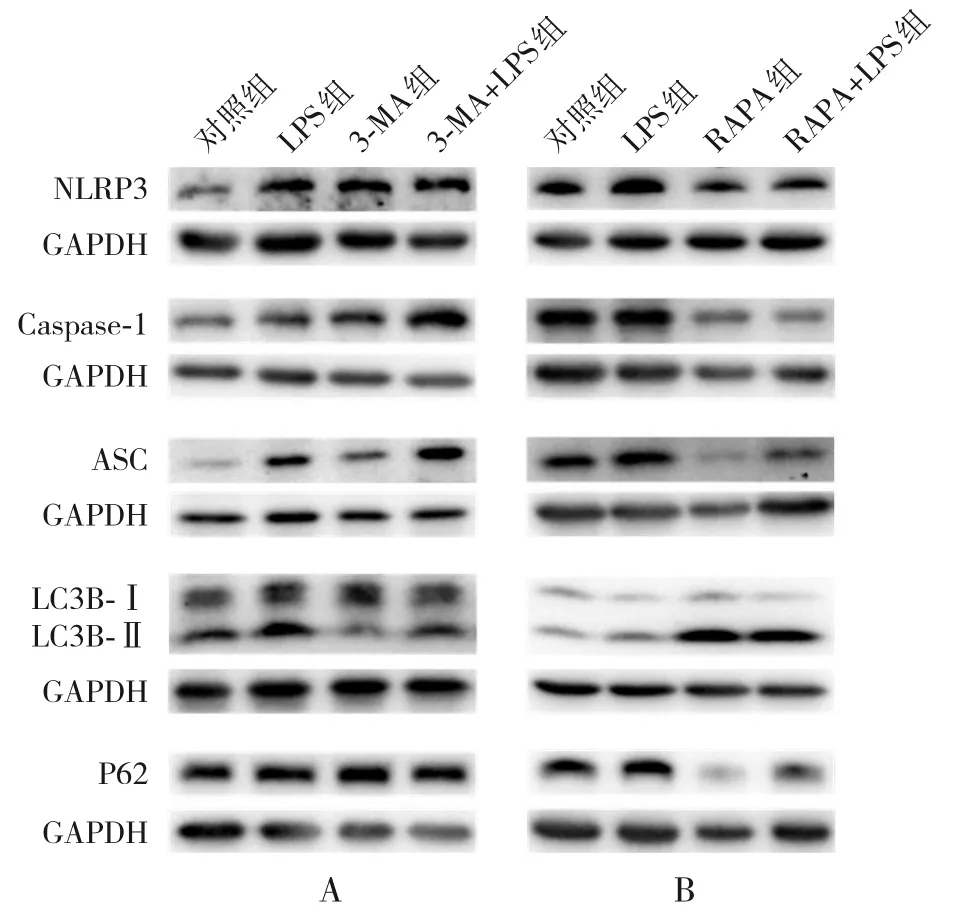
圖5 自噬抑制劑/激動劑對炎癥及自噬蛋白表達的影響
2.5.3 自噬抑制/激動劑對TLR4 蛋白表達水平的影響 在自噬抑制劑3-MA 的處理下,3-MA 組(0.531±0.036)較對照組(0.325±0.042)TLR4蛋白表達上調;經LPS(1 μg/mL,12 h)刺激后,3-MA+LPS 組(0.620±0.018)較LPS 組(0.475±0.032)TLR4蛋白表達水平進一步增加,差異有統計學意義(P<0.05);在自噬激動劑RAPA 的處理下,RAPA 組(0.392±0.088)較對照組(0.825±0.039) TLR4 蛋白表達下調;經LPS (1 μg/mL,12 h)刺激后,RAPA+LPS 組(0.445±0.129)較LPS 組(1.003±0.046)TLR4 蛋白表達水平降低,差異有統計學意義(P<0.05),見圖6。
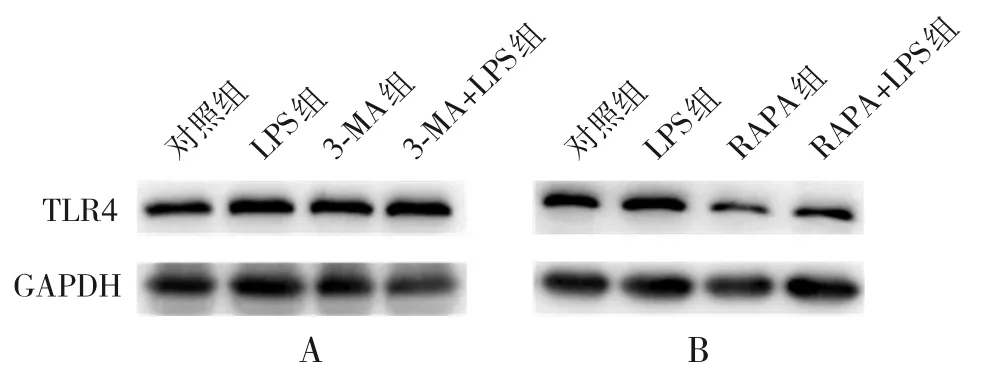
圖6 自噬抑制/激動劑對TLR4 蛋白表達的影響
2.6 TLR4對A549細胞炎癥及自噬水平的影響
2.6.1 過表達TLR4 對A549 細胞炎癥及自噬水平的影響 過表達TLR4 后,自噬相關蛋白LC3B-Ⅱ/Ⅰ表達明顯降低,P62蛋白表達明顯升高;炎癥指標Caspase-1、ASC 的蛋白表達水平呈升高趨勢,與TLR4過表達對照組比較,差異均有統計學意義(P<0.05)。 經LPS (1 μg/mL, 12 h) 刺 激 后,TLR4過表達+LPS組自噬相關蛋白LC3B-Ⅱ/Ⅰ表達仍是降低的,P62蛋白表達則進一步升高;炎癥指標Caspase-1、ASC 蛋白表達水平進一步升高,與TLR4過表達對照+LPS組比較,差異均具有統計學意義(P<0.05),見圖7和表5。
表5 TLR4過表達后炎癥及自噬相關蛋白表達的比較 (±s,n=3)
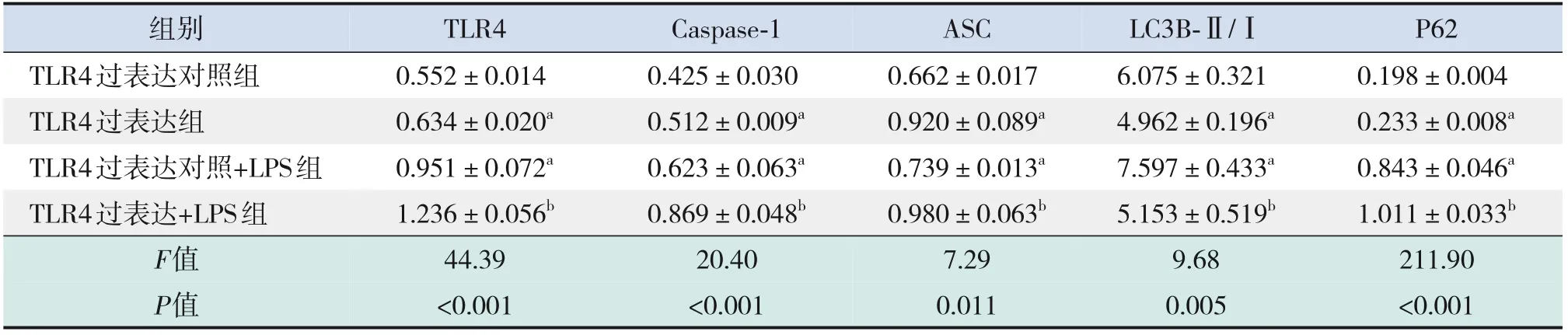
表5 TLR4過表達后炎癥及自噬相關蛋白表達的比較 (±s,n=3)
注:a示與TLR4過表達對照組比較,P<0.05;b示與TLR4過表達對照+LPS組比較,P<0.05。[LPS]脂多糖;[TLR4]Toll樣受體4。
組別TLR4過表達對照組TLR4過表達組TLR4過表達對照+LPS組TLR4過表達+LPS組F值P值TLR4 0.552±0.014 0.634±0.020a 0.951±0.072a 1.236±0.056b 44.39<0.001 ASC 0.662±0.017 0.920±0.089a 0.739±0.013a 0.980±0.063b 7.29 0.011 Caspase-1 0.425±0.030 0.512±0.009a 0.623±0.063a 0.869±0.048b 20.40<0.001 LC3B-Ⅱ/Ⅰ6.075±0.321 4.962±0.196a 7.597±0.433a 5.153±0.519b 9.68 0.005 P62 0.198±0.004 0.233±0.008a 0.843±0.046a 1.011±0.033b 211.90<0.001
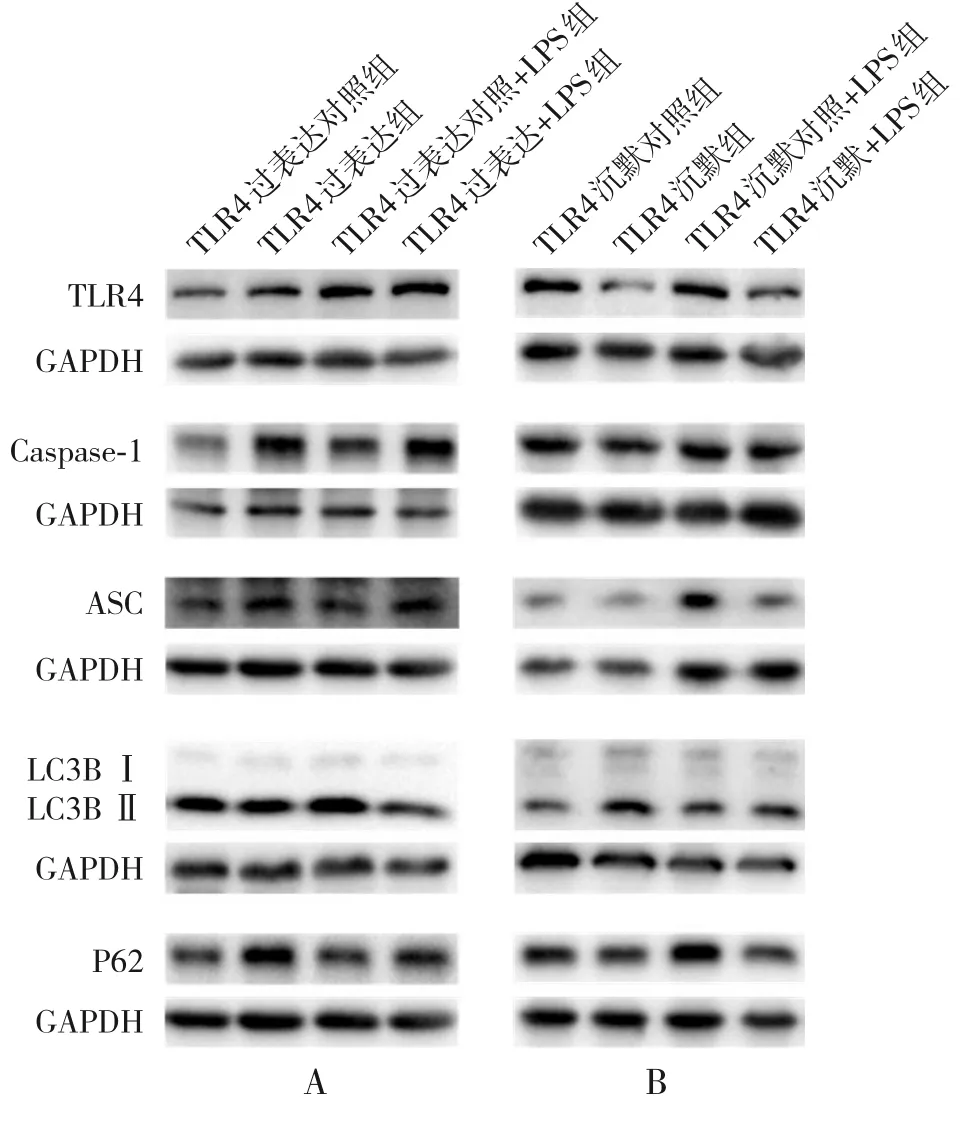
圖7 TLR4 過表達/沉默對A549 細胞中炎癥及自噬蛋白表達的影響
2.6.2 沉默TLR4 對A549 細胞炎癥反應及自噬水平的影響 沉默TLR4 后,自噬相關蛋白LC3B-Ⅱ/Ⅰ表達升高,P62 蛋白表達下降;而炎癥指標Caspase-1、ASC 蛋白表達水平明顯降低,與TLR4沉默對照組比較,差異具有統計學意義(P<0.05)。經LPS(1 μg/mL,12 h)刺激后,TLR4 沉默+LPS組自噬相關蛋白LC3B-Ⅱ/Ⅰ的表達進一步升高,P62蛋白表達水平降低;炎癥指標Caspase-1、ASC的蛋白表達水平進一步降低,與TLR4 沉默對照+LPS 組比較,差異均具有統計學意義(P<0.05),見圖7和表6。
表6 TLR4沉默后炎癥及自噬相關蛋白表達的比較 (±s,n=3)

表6 TLR4沉默后炎癥及自噬相關蛋白表達的比較 (±s,n=3)
注:a示與TLR4沉默對照組比較,P<0.05;b示與TLR4沉默對照+LPS組比較,P<0.05。[LPS]脂多糖;[TLR4]Toll樣受體4。
組別TLR4沉默對照組TLR4沉默組TLR4沉默對照+LPS組TLR4沉默+LPS組F值P值TLR4 0.831±0.038 0.563±0.087a 0.990±0.040a 0.756±0.062b 8.63 0.007 Caspase-1 1.024±0.026 0.900±0.023a 1.107±0.009a 0.982±0.042b 9.93 0.005 ASC 0.612±0.037 0.369±0.058a 0.794±0.037a 0.560±0.026b 18.07<0.001 LC3B-Ⅱ/Ⅰ4.185±0.155 8.890±1.185a 11.660±0.723a 30.010±5.661b 15.05 0.001 P62 0.842±0.009 0.699±0.020a 0.997±0.040a 0.765±0.031b 21.75<0.001
3 討論
ALI/ARDS 的病理生理特點是破壞肺泡毛細血管膜屏障,導致過度炎癥反應和肺功能障礙[6]。因此,抑制炎癥對ALI/ARDS 至關重要。LPS 是革蘭陰性菌細胞壁的主要成分,可刺激機體產生一系列炎癥反應。特別是當LPS影響肺部時,會誘發ALI,嚴重時可發展為ARDS。因此,LPS被廣泛用于ALI/ARDS臨床相關動物或細胞模型的建立[7-8]。Ⅱ型肺泡上皮細胞的炎癥損傷是ALI/ARDS發病過程中的主要病理生理變化,LPS暴露可導致這一變化[9]。而人肺泡上皮細胞株A549 與Ⅱ型肺泡上皮細胞特征最為相似,是體外構建ALI/ARDS炎癥模型使用最為廣泛的細胞系[10-11]。因此,本實驗采用A549 細胞培養模擬LPS 誘導的炎癥反應及肺損傷,是常用的替代原代細胞培養的方法。
在 本 研 究 中,A549 細 胞 經LPS (1 μg/mL,12 h)刺激后,炎癥指標NLRP3、Caspase-1和ASC蛋白表達水平均升高并達到高峰。這些結果表明由LPS 誘導的A549 細胞炎癥反應模型成功建立。同樣條件下,自噬指標LC3B-Ⅱ/Ⅰ、Beclin-1 和P62蛋白表達水平也升高達到高峰,這表明LPS雖然誘導自噬產生,但自噬流受到阻滯。自噬過程涉及幾個步驟:啟動和成核、擴增、自噬體形成、自噬體-溶酶體融合和降解。成核階段磷脂酰肌醇3-激酶(phosphatidylinositol 3-kinase,PI3K)復合物的核心成分Beclin-1被激活以調節自噬體的大小和數量[12]。成核后,LC3-Ⅰ與磷脂酰乙醇胺結合,形成僅在成熟的自噬體上表達的LC3-Ⅱ形式。LC3-Ⅰ向LC3-Ⅱ轉化的增加是自噬體形成的指標[13]。自噬體與溶酶體的融合是自噬貨物降解的關鍵事件[14]。P62/SQSTM1作為一種機制來傳遞泛素化的蛋白質,通過與LC3相互作用靶向物質于自溶酶體處降解[15]。當自噬被抑制時P62 會積累,而當自噬被誘導時可以觀察到P62 水平降低。因此,P62常用作研究動態自噬流的標志物。自噬流始于自噬體的形成,并以溶酶體的底物降解結束,LC3B 代表自噬流啟動,而P62 代表自噬流終止,完整通暢的自噬流才能使自噬真正發揮作用,達到降解物質的目的。本研究CCK-8 實驗結果顯示,LPS濃度過高或者刺激時間過長都會對細胞造成較大的損害,影響各實驗指標的表達。因此,結合上述實驗結果,我們認為1 μg/mL 濃度的LPS 刺激12小時是誘導A549細胞炎癥反應模型,探究自噬作用的最適條件。
為進一步探究自噬和炎癥反應之間的關系,我們選擇在細胞模型上調節自噬來觀察炎癥反應的變化。3-MA 是一種PI3K 抑制劑,通過阻止PI3KC3與beclin-1結合形成自噬體來抑制自噬,是常用的自噬抑制劑。RAPA通過抑制哺乳動物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)復合物的活性來激活自噬,是常用的自噬激動劑。在3-MA(5 mmol/L,12 h)的處理下,LC3B-Ⅱ/Ⅰ表達減少,P62表達增加,自噬受到抑制且自噬流受阻,而炎癥指標NLRP3、Caspase-1和ASC蛋白表達水平升高;在此基礎上,LPS的刺激使P62蛋白表達水平進一步升高,自噬流進一步受到阻滯,同時,NLRP3、Caspase-1 和ASC 蛋白表達水平也進一步升高。而當RAPA 激活自噬后,自噬流恢復通暢,炎癥反應減輕。這些結果說明自噬流對炎癥反應起到負調控的作用。這與許多研究結果一致,表明維持正常的自噬流具有潛在的抗炎作用,比如谷胱甘肽S-轉移酶P1 通過抑制PI3K-Akt-mTOR通路促進自噬流通暢對LPS誘導的炎癥反應發揮保護作用[16];雌激素相關受體α 通過增加自噬流和控制宿主腸道微生物群來改善結腸炎癥反應,從而充當腸道穩態的關鍵調節劑[17];LPS可誘導自噬流阻塞而促發炎癥反應,褪黑激素通過促進自噬流而具有有效的抗炎作用[18]。這也說明,維持一定水平的自噬是必要的,自噬在炎癥反應中可起到一定的細胞保護作用。
TLR4被稱為自噬的環境傳感器[19]。在本研究中,LPS的刺激和3-MA的自噬阻斷均可引起TLR4蛋白表達上調,而RAPA 的自噬激活則引起TLR4蛋白表達下調。為了確定TLR4對自噬及炎癥反應的影響,我們使用TLR4 過表達/空載質粒和針對TLR4的siRNA/陰性對照siRNA轉染A549細胞,用LPS 處理細胞并評估自噬流。研究結果顯示,TLR4過表達組,LC3B-Ⅱ/Ⅰ表達減少,P62表達升高,自噬水平降低,自噬流阻滯;TLR4 沉默組,LC3B-Ⅱ/Ⅰ表達升高,P62表達減少,自噬水平升高,自噬流通暢,表明TLR4對自噬流具有負調控作用。在LPS 的刺激下,TLR4 過表達+LPS 組LC3B-Ⅱ/Ⅰ表達仍是降低的,P62表達則進一步升高,自噬流進一步受到阻滯;而沉默TLR4+LPS組自噬水平進一步升高,自噬流通暢,說明LPS 經TLR4 對自噬流具有負調控作用。同時我們發現,當自噬流阻滯時,炎癥反應進一步升高;當自噬流通暢時,炎癥反應減輕。自噬被認為是炎癥反應的主要負調節因子[20]。本研究證明TLR4介導自噬流在LPS 誘導的A549 細胞炎癥反應中發揮負調控作用。自噬對細胞的保護作用是建立在自噬流通暢的基礎上,自噬流的通暢性抑制了LPS引起的炎癥級聯反應,這可能為ARDS有效的抗炎治療提供一種新方法。
本實驗仍有不足之處。在細胞系選擇方面,沒有使用原代肺泡上皮細胞,研究的借鑒意義相對受限,另外尚未進行在體動物實驗驗證,有待下一步深入研究。自噬流的檢測方法除了免疫印跡法,還有自噬mRFP-GFP-LC3雙標示蹤法等,均有待后續補充。本研究中,我們對自噬的研究著眼于自噬流這一關鍵點,初步闡述自噬對ARDS中炎癥反應的調控機制,這可能僅僅是自噬復雜調控網絡中的一小部分,關于肺上皮細胞自噬的上游信號或調節因子我們依然知之甚少。自噬在肺損傷中扮演著“雙刃劍”的角色,值得我們進行更多地探索。新生兒作為一個特殊的階段,新生兒疾病的防治一直在不斷更新與完善。通過自噬來了解和重新認識新生兒ARDS是一個新角度,可能為新生兒ARDS防治找到新的突破口,為降低新生兒疾病的發生率、病死率等作出貢獻。
利益沖突聲明:所有作者均聲明不存在利益沖突。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
家庭醫學(下半月)(2019年9期)2019-10-12 08:04:06
家庭醫學(下半月)(2019年8期)2019-09-25 09:02:00
人大建設(2019年12期)2019-05-21 02:55:32
媽媽寶寶(2017年3期)2017-02-21 01:22:12
罕少疾病雜志(2016年4期)2016-03-11 16:34:39
學苑創造·A版(2015年11期)2016-01-14 09:03:27
中國衛生標準管理(2015年8期)2015-01-26 18:08:35
中國火炬(2010年8期)2010-07-25 11:34:30
