非洲豬瘟病毒特性
2022-09-29 03:56:02孟祥光譯自Vol287202010月
國外畜牧學(豬與禽) 2022年4期
關鍵詞:研究
孟祥光 譯自,Vol.287(2020),10月
王祎汀 校 潘雪男 審 張濤 制圖
非洲豬瘟(African swine fever,ASF)是一種病毒性、出血性疾病,對家豬和歐亞野豬具有極高的致死率。盡管引發該病的病毒非洲豬瘟病毒(African swine fever virus,ASFV)的宿主范圍有限,也不可能引發人畜共患病,但其對社會、經濟的影響巨大,許多利益相關者都被牽涉其中。出于這個原因,一旦豬群暴發該病,發病地所在行政管理部門應向世界動物衛生組織(Word Oranisation for Animal Health,OIE)通報。在最壞的情況下,ASF 會涉及到家豬、野生動物的病毒貯藏宿主(即野豬)、無生命的寄生體(如豬的尸體、被污染的棲息地、畜牧生產工具、其他機械載體)以及活的節肢動物載體(如軟蜱)。非洲豬瘟控制仍依賴于嚴格的衛生措施,因為目前既沒有獲得許可的疫苗,也沒有任何有效的治療方法。
2007 年,起源于撒哈拉以南非洲森林循環(sylvatic cycle)中的ASF 傳入格魯吉亞,隨后跨高加索地區侵入俄羅斯。該病毒進一步傳播,在2014 年傳入歐盟。2018 年8 月,ASFV 傳入世界上最大的生豬生產國——中國,目前該病毒正在亞洲一些國家或地區的豬群中蔓延。最近受影響的國家是澳大利亞和與之相鄰的巴布亞新幾內亞,以及2020 年暴發ASF的印度。因此,在過去的13 年,ASF 發生了空前絕后的傳播,并得到了各國或各地行業管理部門空前重視,目前的大流行甚至影響到了毫不相關的行業。亞洲的非洲豬瘟疫情暴露了當地獸醫和農業行政管理部門在防控方面的弱點,也暴露了養豬業與副產品回收使用之間的各種直接和間接聯系。非洲豬瘟疫情不僅導致肝素以及用于食品和糖果生產的明膠的供應受到影響,而且還導致動物脂肪、皮和毛的利用受到影響。此外,如果不小心將豬源性產品作為養豬生產中豬蛋白質營養的來源,那么這些產品將會成為ASF 的“加速器”。
在此背景下,本文力圖總結現有數據,特別是在過去5 年中獲得的知識,并就主要的知識空白進行歸納總結。對于后者,筆者借鑒了全球非洲豬瘟研究聯盟最近的差距分析報告(2018 年的報告可在網上查閱:https://www.ars.usda.gov/GARA/reports.htm)。
1 病毒特性
1.1 ASFV 的特征和復制
ASFV 是一種大型雙鏈DNA 病毒,屬于非洲豬瘟病毒科(Asfarviridae) 非洲豬瘟病毒屬(Asfivirus)。根據國際病毒分類委員會2019 年的分類公告(EC 51,德國柏林,2019 年7 月),非洲豬瘟病毒科已被納入痘疹病毒綱(Pokkesviricetes)、阿福病毒目(Asfuvirales)。除了這一官方命名外,人們還討論了將ASFV 納入暫定的巨型病毒目(Megavirales),該目包含單源但異質的核胞質大DNA 病毒(nucleo-cytoplasmatic large DNA viruses,NCLDV)支目。該暫定目還包括痘病毒科(現在屬于Chitovirales新目)、Iridoviridae科(現在屬于Pimascovirales目)、Asfarviridae科、Phycodnaviridae科(現在屬于Algavirales目)、Mimiviridae科(現在屬于Imitervirales目)、Ascoviridae科 (現在屬于Pimascovirales目)和Marseilleviridae科(現在屬于Pimascovirales目)。由于新的巨型病毒不斷被發現,如潘多拉病毒(Pandoraviruses)、福斯特病毒(Faustoviruses)、軟體病毒(Molliviruse)、高茅巴病毒(Kaumaoebavirus)、塞德魯特病毒(Cedratviruses) 以及帕克曼病毒(Pacmanvirus),這組病毒在不久的將來可能會增加新成員,命名法則仍在討論中。
ASFV結構非常復雜,直徑為175~215 nm。到目前為止,研究認為,ASFV 由一個核蛋白核心(直徑70~100 nm)、一個由內部脂質層包圍的核心殼、一個具有1 892~2 172 個帽狀體的二十面體蛋白衣殼和一個可有可無的脂質包膜組成。然而,有關ASFV 結構和構造的詳細情況仍不清楚。
最近,ASFV 粒子三維結構單粒子的低溫電鏡分析結果顯示,病毒核心(病毒基因組和相關蛋白,即DNA 結合蛋白p10、pA104R 和部分轉錄機制復合物) 實際上被兩個不同的二十面體蛋白衣殼和兩個脂蛋白膜包圍。一個是圍繞著對稱性二十面體的內殼;一個是外層,源于出芽過程(budding process)。內蛋白層以T=19 衣殼的形式組織,限制了該核心殼。它包含來自病毒多聚蛋白pp220(p5 蛋白、p14 蛋白、p34 蛋白、p37 蛋白和p150 蛋白) 和pp62(p35 蛋白、p15 蛋白和p8 蛋白)的多個蛋白質。鑒于β 鏈的含量,p15 蛋白可能是該層的主要衣殼蛋白。外層衣殼[T=277的三角(triangulation) 分數] 形成一個六邊形晶格(lattice),由主要衣殼蛋白p72 的8 280 個拷貝(以三聚體排列)和位于頂端的五聯體蛋白(penton protein) 的60 個拷貝組成。簡而言之,細胞外病毒由通過出芽獲得的外膜、外衣殼、二十面體膜、二十面體內衣殼以及封閉的核殼和類核組成,該病毒結構示意圖如圖1 所示。
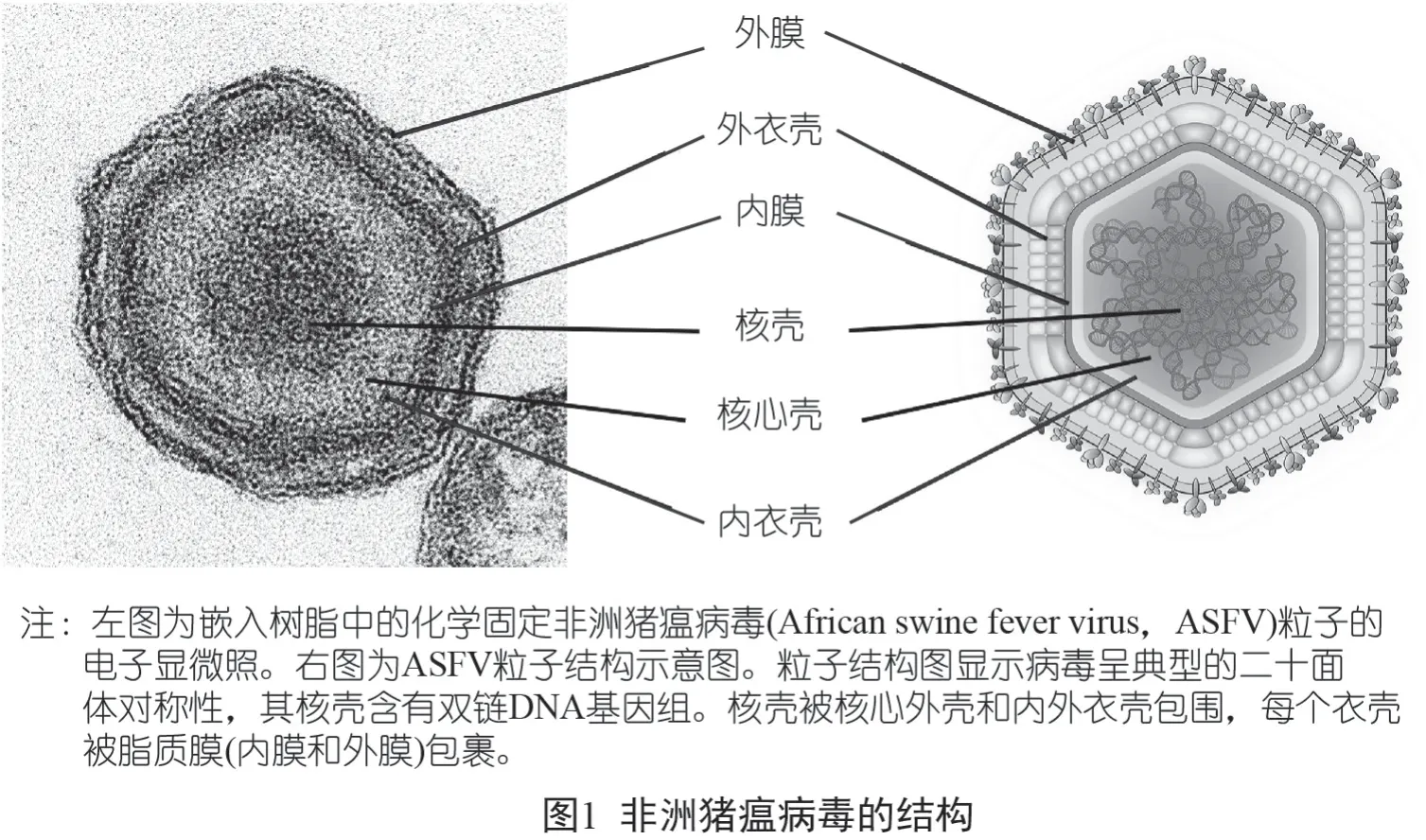
最近,ASFV 復雜的衣殼結構被進一步解開,顯示主要的衣殼蛋白(p72 蛋白) 和四個穩定的次要蛋白(H240R 蛋白、M1249L 蛋白、p17蛋白、p49 蛋白)以五邊形和三邊形對稱體的方式相互作用。在p72 蛋白的形成過程中,需要B602L 蛋白作為伴侶蛋白。
ASFV 基因組由一個170~190 kb 的雙鏈DNA 分子組成,包含151~167 個開放閱讀 框(open reading frames,ORFs),ORFs的數量取決于ASFV 的毒株。ASFV 基因組的兩端由末端反向重復序列(inverted terminal repeats) 和發夾環(hairpin loops) 組成。ASFV 基因組編碼了許多參與病毒組裝、DNA 復制和修復的蛋白質;還編碼參與免疫調節的蛋白質,如干擾I 型干擾素和細胞死亡途徑的蛋白質。大約一半的ASFV 基因仍無任何已知或可預測的功能。
直到最近,根據二維分析結果,已知ASFV擁有54 種結構蛋白。如上所述,ASFV 外膜是通過出芽方式從宿主細胞中獲得的,病毒附著蛋白p12 似乎定位于外膜。另一個值得關注的外膜蛋白是EP402R基因產物CD2v 蛋白。它與T 淋巴細胞表面黏附受體CD2 具有序列同源性。這種蛋白是血細胞吸附現象所必需的,可能對病毒的致病機制極為重要,對病毒在節肢動物體內的復制也很重要。它還與適配蛋白1(adaptor protein 1,AP-1)相互作用,并可能參與細胞通路重塑(cellular traffice remodeling)。ASFV 弱毒株通常為CD2v 蛋白截短。外膜中的另一個蛋白為名叫p24 蛋白的細胞蛋白。該衣殼的主要成分是p72 蛋白,還含有pB438L蛋白(p49蛋白)、pE120R蛋白(p17蛋白)、H240R 蛋白和M1249L 蛋白等。其內膜似乎來自于內質網,包含膜蛋白p54、p17、p12(在外膜中也被發現) 和pE248R 蛋白。所謂的核心殼,現在被定義為內衣殼,由pp220 蛋白和pp62 蛋白以及pS273R 蛋白組成。后者是一種處理病毒多聚蛋白的酶。順序切割生成p150蛋白、p37 蛋白、p34 蛋白、p14 蛋白、p35蛋白和p15 蛋白。如上所述,p15 蛋白可能是該內衣殼的主要衣殼蛋白。該核心殼中的核蛋白是p10 蛋白和pA104R 蛋白(組蛋白樣蛋白),核心應該包含轉錄機制。
最近,蛋白質組學工具的使用使人們能夠更好地掌握病毒蛋白和病毒宿主之間的相互作用。沿著這些思路,Alejo 等利用質譜和免疫電鏡對檢測到的蛋白質進行定位,建立了一個更新的ASFV 粒子與vero 細胞適應株“BA71V”的圖譜。該方案總共確定了68 種病毒蛋白,其中包括所有以前已知的結構蛋白(見上文)和44 種迄今未被識別的蛋白。有趣的是,23 種蛋白質與病毒的某些功能無法關聯起來,但這些蛋白質卻是病毒體的重要組成部分。
Kessler 等在另一項研究中利用ASFV 低致死毒株OURT88/3 株的重組衍生物研究ASFV在易感(野豬) 和非易感宿主(人、猴)的哺乳動物細胞系中的基因表達。在ASFV OURT88/3 株的157 個ORFs 中,94 個被鑒定為蛋白質。有趣的是,一些最豐富的蛋白質也未被定性,其中包括pK145R 蛋白和pI73R 蛋白,它們是今后需要研究的候選蛋白。Karger等總結并比較了上述兩項研究。可以看出,對于大多數預測的ORFs 來說,可以證明蛋白質的存在。然而,對于其他的ORFs 來說,例如多基因家族的成員,仍然缺乏表達的證據。這些ORFs 可能只在被感染的宿主或其主要靶細胞中發揮作用。這也可以解釋為什么這些基因在ASFV 的細胞培養適應株中經常被剔除。
上述研究是利用永久細胞系而非ASFV 主要靶細胞進行的,這可能會對研究結果產生重大影響。到目前為止,很難產生一個又大又純的受感染的巨噬細胞群。不過,相關研究機構正在探討分類、同步和改進的方案,以備將來研究使用。
ASFV 主要在單核吞噬系統的細胞中復制,并且通過網格蛋白介導和縊斷蛋白(dynamin)依賴的內吞作用和巨胞飲作用進行。研究表明,肌動蛋白依賴性的內吞作用和涉及微管活性的內吞通量也與此有關,即經典吞噬作用。其細胞受體及其病毒配體尚不清楚,一個假定的CD163 受體仍存在爭議,但CD163 基因敲除的豬對ASFV 仍然易感。Lithgow 等在與豬骨髓黏附細胞易感性相關的表面標記中發現了CD45 受體。
ASFV 進入細胞內后,要解開病毒的衣殼需要啟動整個內肢體途徑(endosomal pathway),衣殼在內體腔的酸性環境中分解。在pE248R 蛋白介導下衣殼和融合膜降解,病毒核心被釋放到細胞質中。病毒DNA 的釋放需要泛素-蛋白酶體系統的作用。病毒的復制和組裝在靠近細胞核的特殊病毒工廠中進行,新形成的病毒通過出芽從感染的細胞中釋放出來。至于強烈調控轉錄和RNA 加工,則需要病毒編碼的酶。Almazan 等指出,可以區分即時的、早期的、中期的和晚期的RNA 轉錄物。雖然病毒復制的主要階段在細胞質中進行,但Rojo 等指出早期的成核階段也發生在細胞質中,此階段牽涉核纖層網絡(lamina network) 的解體和核蛋白的重新分布。最近,Sim?es 等詳細回顧了成核階段的作用,并對目前掌握的信息進行了匯編。可以證明,ASFV 感染激活了DNA 損傷反應(DNA damage response,DDR) 途徑,改變了細胞核形態(nuclear landscape)和細胞表觀遺傳特征,促進了有效感染和后代的形成。例如,ASFV的有效復制取決于與細胞周期控制、凋亡和免疫反應相關的早幼粒細胞白血病核體的相互作用。此外,異染色質是經誘導形成的,很可能是為了使編碼不利于病毒復制的蛋白質的宿主基因保持沉默。這些病毒誘導的重排可能解釋了上述的核纖層網絡的裂解。綜上所述,ASFV破壞了亞核結構域和染色質結構,并誘導核結構改變,以加強壓制性核環境,使病毒能在宿主細胞中有效復制。然而,細胞核的全部作用仍不清楚,需要進一步研究。
最近,Galindo 和Alonso 對ASFV 一般復制進行了更詳細回顧,Rodríguez 和Salas 則特意對轉錄進行了詳細綜述。
差距和進一步研究的方向:
○對ASFV 毒力基因、宿主范圍和病毒-載體-宿主的相互作用知之甚少。需要大量ASFV 基因型高質量的全長基因組序列,以便能夠對病毒有更深入的了解。
○在ASFV 的150~170 個ORFs 中,大多數只是預測,很少有RNA 水平或蛋白質水平上的實驗性證據。因此,需要進一步研究以更好地了解病毒蛋白及其在不同條件下的表達。
○還需要了解宿主方面的反應,特別是病毒受體、先天性免疫反應以及病毒與宿主在細胞水平上的互作。
○應加快對ASFV 蛋白質功能的研究。
1.2 ASFV 活力的穩定性和滅活
ASFV 在環境和生豬肉制品中高度穩定,低溫、潮濕和富含蛋白質的環境有利于其存活。因此,ASFV 在冷藏豬肉中的感染期可長達15 周,在腌制的火腿中可長達6 個月,在帕爾瑪火腿中可達399 d。在液態豬糞中,ASFV能夠穩定存活100 d 以上;在豬的液態血液中,室溫下ASFV 可存活18 個月,在4 ℃下最多存活6 年。歐洲食品安全局(European Food Safety Authority,EFSA) 在《非洲豬瘟科學報告》中給出了詳細的清單。最近和正在進行的研究探討了與ASFV 間接傳播有關的材料,如飼料、食物、土壤和尸體。對于飼料原料,Stoian 等的研究重點是跨洋運輸時ASFV的穩定性。在這些條件下計算出的ASFV 的半衰期從9.6 d 到14.2 d 不等,具體時間取決于ASFV 所處的基質。此外,Niederwerder 等研究發現,ASFV 可以通過被污染的液體和干飼料傳播。在考察這些研究的結果時,病毒的載量和拷貝數是重要的影響因素,因此需要進行更多的研究。在一項關于ASFV 在受污染作物中活力穩定性的實驗室規模的研究中,研究人員探討了干燥和熱滅活對病毒活性的影響。總之,ASFV 在干燥的室溫環境中經過2 h 被滅活。關于ASFV 在豬胴體中的穩定性,最近立陶宛的研究人員進行了一項研究。Zani 等將于不同時間掩埋在不同地點的感染了ASFV的野豬尸體挖掘出,隨后利用體外檢測法檢測傳染性ASFV,用qPCR 法測定病毒的基因組。出乎意料的是,從尸體中只檢測出病毒基因組,而所有的病毒分離嘗試結果均為陰性。Carlson 等在最近一項尚未發表的研究中探討了土壤對ASFV 活性的影響。簡言之,在不同的土壤基質中摻入感染野豬的ASFV 陽性血液,隨后檢測病毒的活性。土壤pH、結構和環境溫度對ASFV 活力的穩定性起著重要的作用。對于沙地或花園類的土壤,幾周后仍可分離到活病毒;對于沼澤地的土壤,幾天后可分離到病毒;未能從森林的酸性土壤中分離出病毒。用檸檬酸或氫氧化鈣緩解土壤的酸堿性,結果顯示ASFV 在所有類型的土壤中均完全失活。
研究人員提出的另一個問題是ASFV 在以豬糞便為原料的沼氣廠廢棄物中的滅活情況。Davies 等調查了ASFV 在實驗性感染豬的糞便和尿液中的存活時間。根據計算出的半衰期可以認為,在37 ℃下,ASFV 經過約4 d(尿液)或3 d(糞便)后仍具有傳染力。在沼氣廠的沼氣生產環境中,應考慮如pH 和代謝物等次要因素,因此上述這些時間可以下調。Turner和William 的研究表明,在40 ℃下,豬糞中ASFV 經4 h 即可滅活。Moss 得出的結論是,如果沼氣廠采用正確的生產工藝,數小時(采用高溫生產工藝的沼氣廠)或數天(采用中溫生產工藝的沼氣廠) 可使ASFV 滅活。然而,必須考慮到(與高度安全的實驗室或現代熬煉廠相比),這些工廠在設計生產工藝時通常沒有考慮嚴格的黑/白分離。在工廠中,例如緊隨其后的是化肥廠,可以認為再次污染的情況也將不會發生,因為在真空生產環境中發酵基質的數量會逐步減少。
世界各地都在使用不同種類的消毒劑。在實驗條件下,過氧乙酸或甲酸消毒劑顯示出良好的殺病毒效果。一般來說,ASFV 對脂類溶劑、洗滌劑以及氧化劑敏感。EFSA 發表了一份科學報告,介紹了在田間條件下(與氫氧化鈉和碳酸鈉相比)被授權消毒劑功效的最新數據。原則上可以認為,對其他包膜DNA 病毒顯示出良好滅活功效的消毒劑對ASFV 也有效。
差距和進一步研究的方向:
○飼料、飲水和墊料對ASFV 傳播的作用仍有爭議,需要進一步研究。
○病死豬掩埋地是否需要采用消毒措施仍然是一個沒有定論的問題,需進一步測試ASFV 的穩定性,如使用不同種類的土壤,對改進風險評估是必要的。
○雖然抗包膜病毒的消毒劑在標準化條件下能夠有效殺滅ASFV,但需要找到切實有效的方法,以在資源有限的環境和表面結構復雜的物體(如木質物資、開放性混凝土或其他種類的地板)上測試消毒劑的有效性,并選擇合適的針對ASFV 的消毒劑。
1.3 遺傳多樣性和分型
在過去幾十年里,研究人員利用不同的遺傳區域,通過測定小DNA 片段中部分基因序列對ASFV 分離株進行定型。用于基因型命名的區域通常是p72 編碼區,對p54、p72 和pB602L基因進行測序,可以獲得更好的鑒別能力。基于這一思路,Boshoff 等已經在非洲鑒定出24 種ASFV 基因型。遺傳多樣性通過森林循環得到提高,但這些地區之外并不存在這種情況。此外,病毒的基因型與其毒力或致病性沒有關系。“國際”基因型是Ⅰ型和Ⅱ型。
總體而言,ASFV 非常穩定,基因突變率非常低。這導致該病毒在發病地區的遺傳變異性很低,即使使用下一代測序技術,通常也不能以更高的分辨率對病毒毒株進行分子追蹤,例如在發生疫情時的分子流行病學。只有全基因組才可能有所區別,并可推斷出毒力因子。同樣,通過全基因組測序也能更好地識別新毒株的基因缺失變種。在過去的十年中,一些科學實驗計劃已得到了優化,包括長片段牛津納米孔技術(Oxford Nanopore MinION)測序技術。
在目前的流行狀態下,ASFV 每天都會產生新的基因序列,控制檢測質量是最重要的。否則,測序的錯誤率可能高于病毒復制過程中的錯誤率,而測序錯誤會被誤解和過度解讀為重大差異。最近,為獲得可靠和高質量的全基因組序列,已經建立了一個深度測序流程(deep-sequencing workflow)。這個工作流程基于靶向富集和使用不同測序平臺來規避特定的缺點。國際合作是為未來研究提供可靠數據的關鍵。
差距和進一步研究方向:
○必須努力優化和協調樣本選擇、基因測序、生物信息學工作流程和數據文檔的協議,最有效地利用財政和技術資源。
○在一個有針對性的方法中,應該生成所有可用基因型和宿主物種的全基因組序列,作為進一步研究的基礎。
○應加快發現與非洲豬瘟病毒科有關的病毒,以便進行進化分析。
原題名:African swine fever -A review of current knowledge(英文)
原作者:Sandra Blome、Kati Franzke和Martin Beer
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
