高寒礦區采樣位置和生長期及坡向對草本植物根系力學特性影響
2022-09-29 11:23:46付江濤楊幼清趙吉美劉昌義李世珍胡夏嵩李希來余冬梅
草地學報 2022年9期
付江濤,楊幼清,趙吉美,劉昌義,李世珍,胡夏嵩*,李希來,余冬梅
(1. 青海大學農林科學院, 青海 西寧 810016; 2. 陜西理工大學土木工程與建筑學院, 陜西 漢中 723000; 3. 中國科學院大學, 北京 100000; 4. 青海大學地質工程系, 青海 西寧 810016; 5. 青海大學農牧學院, 青海 西寧 810016; 6.中國科學院青海鹽湖研究所, 青海 西寧 810008)
植物根徑、抗拉力、抗拉強度、極限拉伸應變率和拉伸模量等物理量是反映植物根系力學特性的重要指標,也是定量計算植物根系對土體抗剪強度貢獻的主要參數[1-4]。受植物生長環境,如溫度變化、土壤含水量、土壤營養元素供給、土壤理化生性質和植物生長期以及試驗條件等諸因素影響[5-9],植物根系力學特性通常具有較大變化幅度,這在很大程度上導致植物根系對土體抗剪強度貢獻值難以準確定量評價[9-10]。因此,為提高根系力學指標值測定結果準確度,定量分析生長環境對植物根系力學指標影響已成為生態護坡領域研究熱點問題之一。
目前,關于定量研究植物根系抗拉特性指標的主要方法為:采用單根拉伸試驗,在測定供試種植物每一條單根根徑、抗拉力和拉伸長度前提上,根據文獻[10]中公式(1)、(2)和(3)分別計算單根抗拉強度(MPa)、極限拉伸應變率(%)和拉伸模量(MPa)。在求得單根抗拉強度(MPa)、極限拉伸應變率(%)和拉伸模量(MPa)的基礎上,對上述試驗測定值(根徑、單根抗拉力和伸長量)和計算值(單根抗拉強度、極限拉伸應變和拉伸模量)求取平均值,最后利用各平均值反映各供試種植物根系實際的抗拉特性。根據上述研究方法,Schwarz等[11-12]借助于WWM模型和FBM模型計算了植物根系對土體抗剪強度的貢獻,進而對邊坡穩定性進行了評價。上述方法很大程度上提高了植物根系對土體抗剪強度貢獻的計算精度,然而亦存在諸多不足,例如在開展植物單根拉伸試驗時,不僅需對相同研究區域開展不同位置的采樣(以確保所選取的供試種植物根系力學特性指標具有代表性),且需對大量的根系進行重復性拉伸試驗[13-14],該過程耗時費力和效率低下,而且在相同研究區不同采樣點進行根系采集時,不同取樣點植物因生長環境存在差異,其根徑、單根抗拉力等指標是否存在差異尚不明確,需進一步深入研究[13-15]。
在丘陵地區,生長于同一區域的植物受其坡向等因素影響,其植物群落結構與多樣性等均存在差異[16],而作為體現植物水土保持能力的根系根徑、抗拉力、抗拉強度等指標是否也會產生差異尚不明確,需進一步確定。因此開展不同采樣位置、不同生長期以及不同坡向對植物根系抗拉特性的影響,對于準確計算植物根系對土體抗剪強度貢獻起到理論支撐作用,同時對于不同坡向處邊坡開展生態護坡建設亦有著實際的指導意義。基于此,本研究選取位于青海天峻大通河上游木里煤田江倉礦區復綠區為研究區,通過對生長期為1a(注:a表示年,下同),2a,3a,4a和5a的優勢草本垂穗披堿草根系開展不同位置、坡向和生長期的采樣,在此基礎上對所采樣根系開展單根拉伸試驗,同時測定其根徑、單根抗拉力和拉伸長度,并計算其單根抗拉強度、極限拉伸應變率和拉伸模量;最后通過單因素方差分析,明確采樣位置、坡度和生長期對供試種植物根系單根抗拉特性指標的影響。該項研究結果不僅有助于準確計算植物根系提高土體抗剪強度的貢獻,而且也為地處高海拔、寒旱環境礦區開展生態恢復提供理論支撐。
1 材料與方法
1.1 研究區概況
木里煤田位于青海省海北州天峻大通河上游,海拔高度為3 883 m,地理坐標東經99°28′54″、北緯38°20′49″,位置如圖1所示[17-20]。由該圖可知,研究區地處青藏高原東部地區,屬中祁連山高海拔地區,主要以高原冰緣地貌類型為主,氣候類型為典型的高原大陸性氣候[18-20]。區內沼澤、濕地和高山草甸較為發育,土壤類型以高山草甸土、沼澤草甸土為主,土層厚度10~50 cm,常年凍土覆蓋,植被類型以高寒沼澤類和高寒草甸類為主,主要建群種為藏嵩草(Kobresiatibetica)。區內發育河流主要有上哆嗦河、下哆嗦河、江倉曲及其支流等,均屬大通河水系,河水主要通過高山融雪水、泉水及大氣降水等形式補給。研究區獨特的地質條件及氣候類型使得該區生態環境極為脆弱,加之受人為工程活動等因素影響,該區采礦廢棄坑附近沖溝分布極為廣泛,水土流失、滑坡和泥石流等地質災害較為發育[18-20]。

圖1 研究區位圖示意圖[17]Fig.1 Location of the tested sites[17]
研究區木里煤田江倉礦區經多年開采,在位于遺棄采礦坑南北兩側形成面積分別為73.10 hm2和31.23 hm2的露天排土場[18,20],該排土場主要由腐植表土、風化巖土、堅硬巖石以及混合巖土、煤矸石、煤炭、凍土等組成。受降雨、降雪和凍融作用影響,該排土場極易發生坍塌,產生滑坡和泥石流等地質災害現象,這給當地生態恢復和居民群眾開展牧業活動構成潛在威脅。因此對該區排土場開展植被恢復已成為該區生態保護的主要任務。
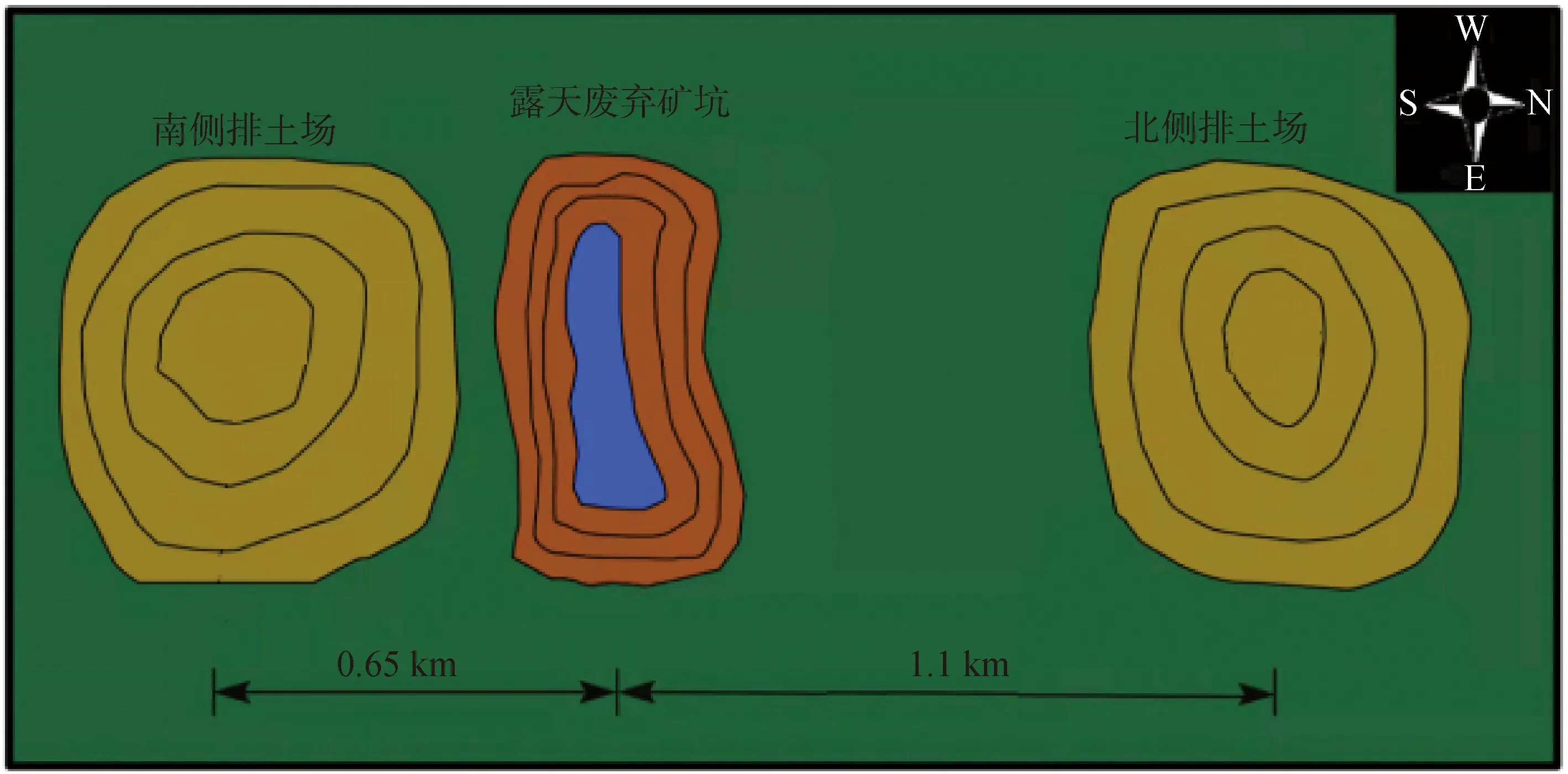
圖2 研究區廢棄采礦坑與南北兩側排土場邊坡位置分布圖(根據Li et al.[17];本研究略作改動)Fig.2 Sampling spots in the tested site (Originated from reference [17] with slight remediation)
1.2 試驗材料與試驗設計
在種植植物前,首先對區內排土場按照分級放緩邊坡的原則進行刷坡整形,待排土場邊坡處理完畢后,根據課題組前期所開展的區內植被類型及分布特征調查結果,本研究篩選出垂穗披堿草(Elymusnutans)、星星草(Puccinelliatenuiflora)和冷地早熟禾(Poacrymophila)等3種優勢草本作為生態恢復植物并開展種植、在此需說明的是,本研究制取試樣的礦區邊坡海拔高度、坡度等條件盡可能保持一致。
2018年8月,在研究區排土場按照不同坡向,即半陰半陽坡(對照組)、陽坡和陰坡確定采樣點。在制取試樣過程中,每處采樣位置隨機選取2~3個采樣點進行采樣,采樣過程為:待采樣樣方(圓形樣方,直徑為30 cm)確定后,用鐵鎬將樣方周邊土挖松,然后用小鏟將挖松的土取出,只保留中間的土柱(高度為:30 cm),最后用鐵鏟將土柱挖出,用木棍將土柱敲散,并對根系進行采集。待根系試樣采集完畢后,立即用保鮮膜將試樣包裹以有效防止水分蒸發,并及時將試樣帶回實驗室開展單根拉伸試驗,具體試驗過程根據文獻[10]進行。最后,為確保試驗數據的代表性和可靠性,本研究所開展的單根拉伸試驗在每個取樣點均進行61次重復。在此需要說明的是,本研究由于植物均為不同時間段種植,具體種植時間為2013,2014,2015,2016和2017年。本研究試樣采集時間為2018年,故植物生長期分別為5,4,3,2和1 a。因此,制取植物根系試樣時,除按照取樣點和坡向采集以外,還按照草本植物生長期進行取樣,每個生長期試樣重復2~3次。受試驗樣本數量限制,本研究難以全面對比同一生長時間條件下,不同坡向對垂穗披堿草根系抗拉特性指標的影響,例如種植于半陰半陽邊坡垂穗披堿草生長期為1,4和5 a,而在區內其它坡向中未采集到相對應生長期的試樣。基于以上實際情況,本研究所采用的分析方法為,首先對比同一生長期條件下,生長于陰坡和半陰半陽坡的垂穗披堿草根系抗拉特性指標;然后,對比生長于陽坡較短生長期和生長于半陰半陽邊坡相對較長生長期條件下,垂穗披堿草根系抗拉特性的差異性。
1.3 單根拉伸試驗與相關數據計算
本研究中,單根抗拉力FR通過單根抗拉試驗獲得,其操作步驟為:設定拉伸標距,在根系拉伸過程中,標距始終為一定值(l)。待標距設定完成后,將植物單根夾持于拉伸儀兩端,同時測定植物根系中部的根徑,進行3次重復,取其平均值(D),然后對植物根系進行拉伸。在根系受到張拉時,與拉伸儀相連的數據記錄儀自動記錄根系受到的拉張力和拉伸長度。當根系拉斷時,記錄的單根最大拉張力和拉伸長度分別為單根抗拉力(FR)和極限拉伸長度(Δl)。此外,在根系張拉時,為避免夾持部位對根系抗拉強度造成影響,僅選取斷裂發生于根系中部的根系為有效根系。當單根抗拉試驗完成后,通過公式(1)、(2)和(3)分別計算單根抗拉強度(tr)、極限拉伸應變率(εr)和拉伸模量(Er)。
(1)
式中,tr為單根抗拉強度(MPa);FR為單根斷裂時的抗拉力(N);D為單根根徑(mm)。
(2)
式中,εr為單根極限拉伸應變率(%);Δl為單根在軸向拉力作用下的伸長量(mm);l為根系的夾持長度(mm)。
(3)
式中,Er為單根拉伸模量或楊氏模量(MPa)。
1.4 數據處理與統計方法
以采樣位置為自變量(Independent variables),植物根徑、單根抗拉力、抗拉強度、極限拉伸應變率和拉伸模量為響應變量(Responsive variables),通過單因素方差分析(ANOVA),探討采樣位置對根系力學指標的影響;然后分別以坡向和生長期為自變量,以根徑、單根抗拉力、抗拉強度、極限拉伸應變率和拉伸模量為響應變量,探討坡向和生長期對響應變量的顯著(P<0.05)和極顯著性(P<0.01)影響。
2 結果與分析
2.1 取樣位置對植物根系抗拉指標影響
表1所示為生長在半陰半陽邊坡不同位置處垂穗披堿草單根拉伸試驗結果。由該表可知,單根平均根徑為0.16~0.21 mm,單根抗拉力為2.35~3.16 N,抗拉強度為88.16~245.46 MPa,極限拉伸應變率為41.05%~55.51%,拉伸模量為185.40~514.69 MPa,其中,根徑和極限拉伸應變率離差均較小(表2),而抗拉強度和拉伸模量離差則相對較大(表2)。此外,對該表進一步分析可知,除生長期為4 a時抗拉強度在不同采樣點表現出極顯著性(P<0.01)差異外,其余各指標均未表現出極顯著性差異,該結果表明,對照組邊坡在相同生長期條件下,采樣點位置不同總體未導致植物根系力學特性各指標產生極顯著性(P<0.01)差異。
如表3所示為種植于陽坡的垂穗披堿草單根拉伸試驗結果。由該表可知根徑為0.19~0.34 mm,單根抗拉力為2.61~3.52 N,抗拉強度為46.52~117.69 MPa,極限拉伸應變率為39.89%~49.65%,拉伸模量為116.88~255.10 MPa。上述試驗結果各指標離差相對較大,為9.92%~ 60.47%,如表4所示。此外,對該表進一步分析得到,對于種植于2015年的垂穗披堿草,其各指標均存在顯著性(P<0.05)或極顯著性(P<0.01)差異,該結果說明生長于陽坡垂穗披堿草單根抗拉性指標因采樣位置不同而產生差異性,造成該結果的主要原因可能與植物生長環境有關,即盡管本研究中植物生長環境(土質、土壤營養元素等)等條件基本一致,然而在植物種植后,由于受植物周圍生長環境因素(溫度、水土流失、營養元素組分)改變的影響,土壤含水率、土體密度和立地條件等均發生改變[12],從而導致垂穗披堿草單根抗拉特性指標值產生改變。
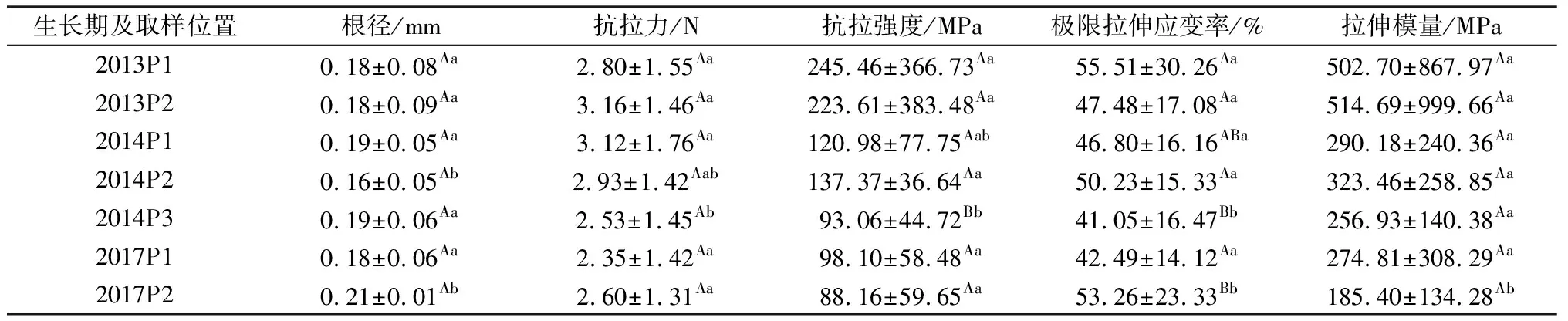
表1 研究區半陰半陽坡面(對照組)垂穗披堿草根系抗拉特性指標測定結果Table 1 Tensile mechanical indices of roots for E. nutants growing in north-south oriented slopes
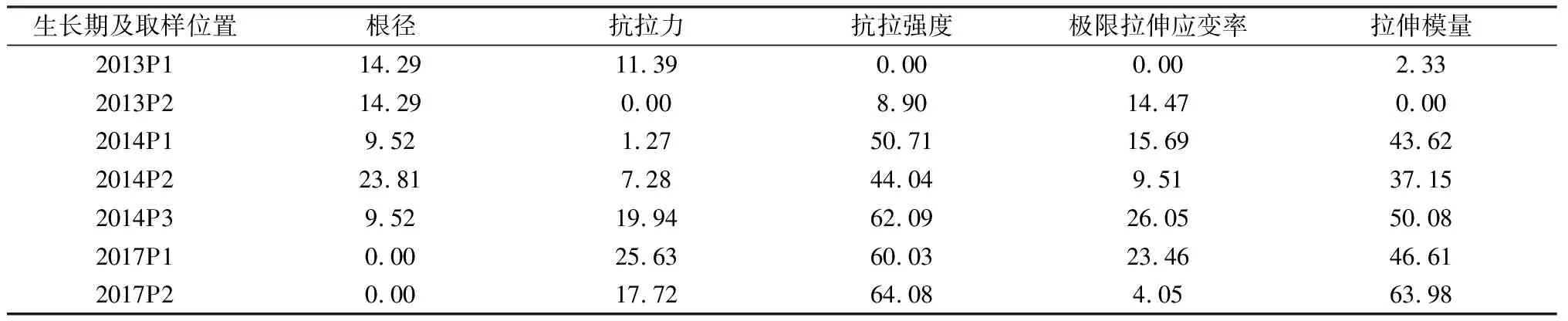
表2 研究區半陰半陽坡面(對照組)垂穗披堿草根系抗拉特性指標離差計算結果Table 2 Deviation of tensile mechanical indices of roots for E. nutants growing in north-south oriented slopes

表3 研究區種植于陽坡坡面的垂穗披堿草根系抗拉特性指標測定結果Table 3 Tensile resistance indices of roots for E. nutants growing in sunny slopes

表4 研究區種植于陽坡坡面的垂穗披堿草根系抗拉特性指標離差計算結果Table 4 Deviation of tensile resistance indices of roots for E. nutants growing in sunny slopes
由表5和表6可知,除單根抗拉力和極限拉伸應變存在極顯著性(P<0.01)差異外,其余指標均不存在差異性,該結果表明生長于陰坡的垂穗披堿草單根抗拉性指標總體上未表現出因其采樣點位置不同而產生差異的特征,且其離差相對較小。區內3種不同坡向邊坡中,在相同生長期條件下,對照組邊坡和陰坡,垂穗披堿草根系抗拉特性指標未表現出隨著采樣位置變化而發生顯著性變化的特征;但在陽坡坡向中,抗拉力、抗拉強度、極限拉伸應變率和拉伸模量等指標,則表現出隨取樣位置變化而發生顯著性(P<0.05或P<0.01)變化的特點,造成不同坡向位置處植物單根抗拉特性指標隨采樣點位置發生變化的原因除與前述營養元素差異有關外,也可能與植物受到的光照因素有關,即陽坡條件下植物受到光照相對較為充足,這在很大程度上促進了植物根系生長,從而提高了根系的抗拉能力;相應地,生長于對照組邊坡或陰坡植物,受到的光照相對較弱,造成垂穗披堿草生長相對較緩慢,從而導致該植物根系抗拉能力表現出相對較弱的特性。

表5 研究區種植于陰坡坡面的垂穗披堿草根系抗拉特性指標測定結果Table 5 Tensile resistance indices of roots for E. nutants growing in shady slopes

表6 研究區種植于陰坡坡面的垂穗披堿草根系抗拉特性指標偏差計算結果Table 6 Deviation of tensile resistance indices of roots for E. nutants growing in shady slopes
2.2 生長期對草本植物根系抗拉指標的影響
圖3所示為垂穗披堿草根系抗拉性指標與生長期之間的關系。由該圖可知,在半陰半陽邊坡中,植物根徑表現出隨著生長期增加呈逐漸降低且各組間不存在極顯著性差異(P<0.01)的特征(圖3a);單根抗拉力隨生長時間增加則表現出遞增的變化特征,且生長期為4 a和5 a的單根抗拉力間無顯著性差異(P=0.52),而與生長期為1 a的存在顯著(P<0.05)或極顯著性差異(P<0.01,圖3d);對于單根抗拉強度、單根極限拉伸應變率和拉伸模量,隨生長時間增加,這3項指標總體上均表現出增加的變化特征。其中,單根抗拉強度在不同生長時間段均表現出極顯著性差異(P<0.01,圖3g),極限拉伸應變率在生長時間為1 a和5 a以及1 a和4 a時不存在顯著性差異(圖3j),而在4 a的和5 a的之間存在顯著性差異。
類似于對照組,對于生長在陽坡的植物,隨著生長期增加,其根徑呈降低的變化特征,且存在極顯著性差異(P<0.01,圖3b),而其他指標則呈增加的變化規律;進一步分析可知,單根抗拉力未表現出顯著性差異(P>0.05),而抗拉強度(圖3e)、極限拉伸應變率(圖3k)和拉伸模量(圖3n)則表現出顯著性差異(P<0.01)。對于生長在陰坡的垂穗披堿草,其根徑為0.18 mm(圖3c)、單根抗拉力為2.30 N(圖3f)、單根抗拉強度為89.62 MPa(圖3i)、單根極限拉伸應變率為43.11%(圖3l)、單根拉伸模量為225.06 MPa(圖3o),且由于生長期均為1 a,在此難以進行比較與判斷;盡管如此,這方面的研究尚有待于進一步地探討。
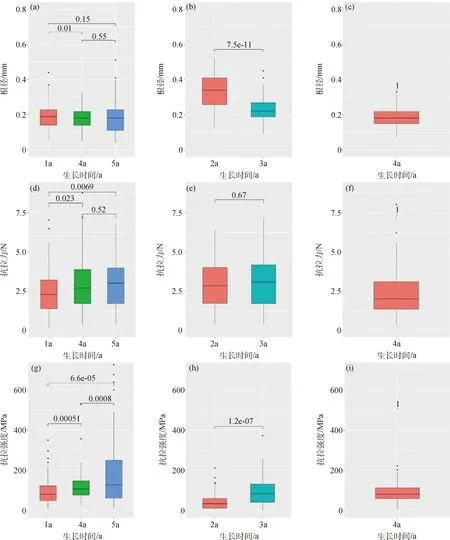
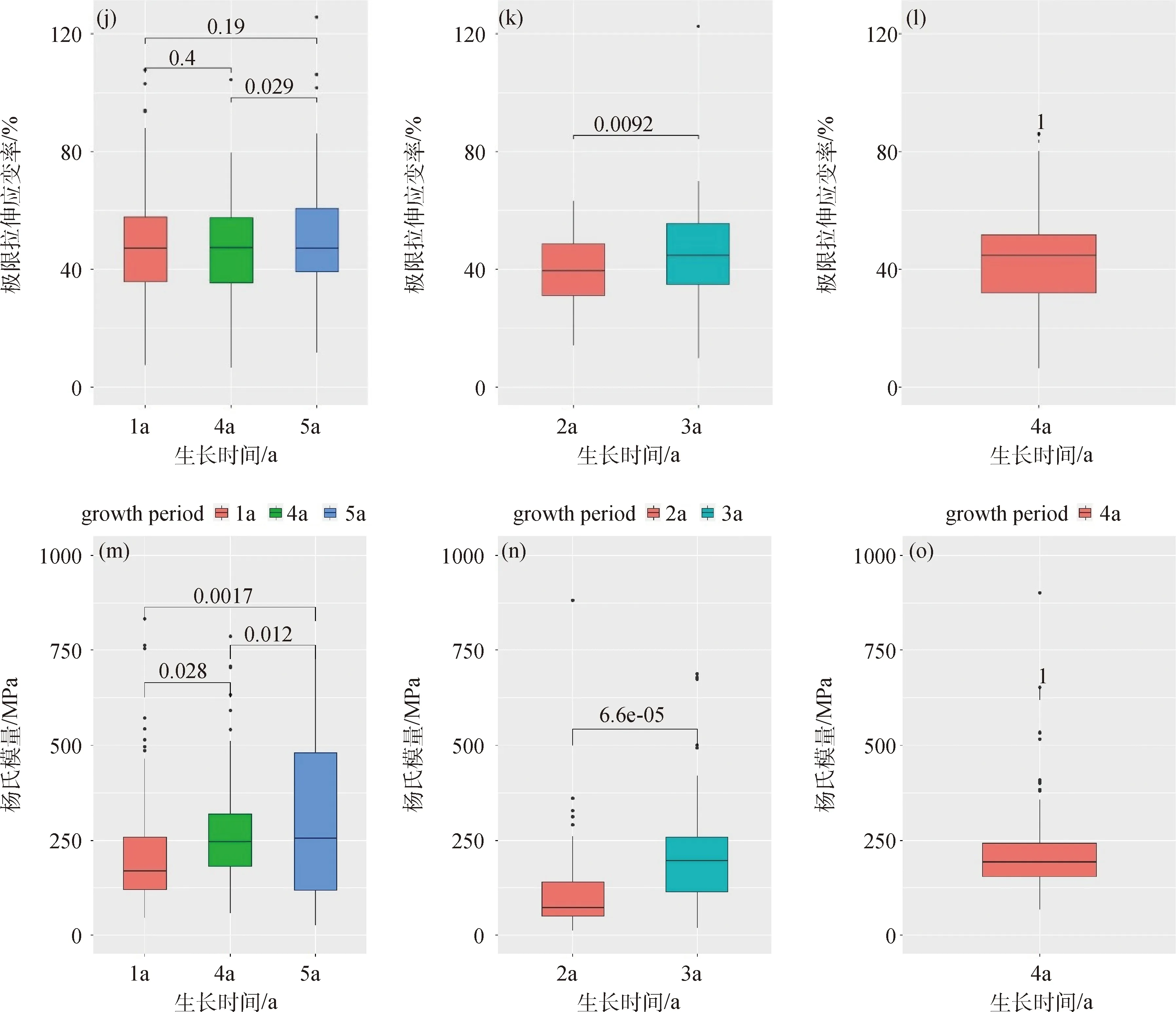
圖3 不同坡向條件下根系抗拉特性指標與生長期之間關系(左圖為半陰半陽,中圖為陽坡,右圖為陰坡)Fig.3 Relationship between tensile resistance indices and growth periods of root for E. nutans in different slope aspects (the left columns are control groups,the middle are sunny slopes and the right are shady slopes)
2.3 坡向對草本根系抗拉特性指標影響
如圖4a所示為生長期為4 a條件下,分別種植于陰坡和半陰半陽邊坡垂穗披堿草單根抗拉特性指標的對比結果。由該圖可知,在相同生長期條件下,生長于半陰半陽邊坡垂穗披堿草根徑、抗拉力、抗拉強度、極限拉伸應變率和拉伸模量分別為0.18 mm、2.86 N、117.39 MPa、46.03%和290.19 MPa,總體上大于生長于陰坡的垂穗披堿草相應數值(陰坡為0.18 mm、2.31 N、89.63 MPa、43.11%和225.06 MPa)。該結果表明,生長于半陰半陽坡的垂穗披堿草各項指標均大于陰坡,這在一定程度上反映出相對充足的光照可顯著提高垂穗披堿草根系抗拉力、抗拉強度和拉伸模量等指標。此外,通過對比生長于陽坡生長期為2 a和生長于半陰半陽邊坡生長期為4 a垂穗披堿草根系抗拉特性指標可知,生長于陽坡的垂穗披堿草根徑和抗拉力是半陰半陽邊坡的1.83倍和1.03倍,而抗拉強度、極限拉伸應變率和拉伸模量則分別為半陰半陽邊坡的0.40倍、0.85倍和0.40倍(如圖4b所示),該結果表明生長于陽坡的草本可有效提高其生長指標(根徑)和抗拉力,但同時亦可降低其抗拉強度、拉伸應變率和拉伸模量等抗拉特性指標值。
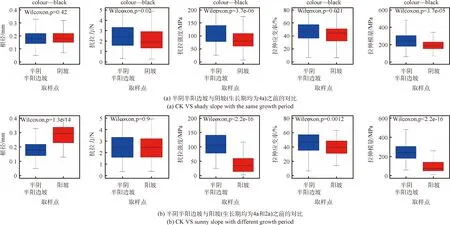
圖4 研究區生長在半陰半陽邊坡、陰坡與陽坡的垂穗披堿草根系抗拉特性之間關系Fig.4 Comparison between tensile resistance indices of root for E. nutans planted in CK,shady and sunny slopes
3 討論
3.1 取樣位置對草本植物根系抗拉指標影響
植物生長的立地條件主要包括氣候、土壤、地形等3個方面,是影響植物生長的重要因素[25-26]。由于垂穗披堿草種植區域面積相對有限,故種植區氣候條件基本相同。此外,種植層基質均由煤礦開采剝離原表土層經二次倒運而成,故植物種植基質理化生性質基本相類似[13-15],因此其土壤和立地條件亦基本相一致,唯一不同的因素為地形條件,即邊坡取樣位置和坡向。相關研究表明,不同立地條件可導致植物生長指標產生差異[24]。本研究表明生長于半陰半陽邊坡和陰坡的垂穗披堿草根徑、抗拉力、抗拉強度、極限抗拉應變率和拉伸模量盡管在量值上存在差異性,但總體上并未表現出極顯著性差異(P<0.01,如表1和表5所示),該結果說明,邊坡取樣位置的不同并未導致生長于半陰半陽邊坡和陰坡的垂穗披堿草根系抗拉特性指標值產生極顯著性差異。
此外,與此不同的是,生長于陽坡的垂穗披堿草各指標均表現出顯著(P<0.05)或極顯著性差異(P<0.01)。該結果表明盡管土壤和氣候條件基本一致,對于生長于同一坡向的垂穗披堿草因其采樣點位置不同,其根系力學特性產生顯著和極顯著性差異。綜上所述,致使生長于陰坡、半陰半陽邊坡和陽坡垂穗披堿草根系力學特性指標,在不同采樣位置表現出不同的差異性的主要原因可能在于陰坡和半陰半陽邊坡條件下,草本植物受到光照強度相對有限,加之區內全年溫度相對較低[18,20-21],每年僅有數月溫度條件適合植物生長,使得植物生長相對較為緩慢,造成植物生物量積累較少,從而導致植物根徑增加不顯著,而生長于陽坡的垂穗披堿草光照相對較足,生長較為迅速,即使不同采樣位置較小的土壤理化生性質的差異(如同一坡向,坡頂和坡腳處土壤含水量可能存在差異),也會導致植物生長指標表現出較大的差異,例如本研究陽坡條件下2個采樣點根徑離差(20.59%~44.12%)大于陰坡(5.26%)和半陰半陽邊坡條件下根徑的離差(9.52%~23.81%),變化幅度較大的根徑在一定程度上會導致其根系力學其它指標表現出顯著或極顯著性差異。
3.2 生長期對植物根系抗拉指標影響
植物生長時間是導致植物根系抗拉特性指標產生差異的重要因素,隨著生長時間增加,植物根徑呈逐漸增大的變化趨勢[22]。本研究中隨著生長時間增加,植物根徑出現降低變化特征,其原因可能在于以下方面:在研究區全年較低的氣溫以及夏季晝夜溫差相對較大的因素作用下[18,20],區內土壤極易發生凍融循環,從而造成該區嚴重的土壤凍融侵蝕,加之,植物種植區人工堆積邊坡坡長相對較長且無遮蔽物,夏季在坡面徑流作用下,土壤營養元素經坡面徑流搬運而發生流失,導致土壤養分虧缺,致使其后植物生長因為養分虧缺而發生退化現象,從而導致后續年份中植物新生根系較細,進而提高細根在總根系中的比例,使得測定的根徑隨生長期增加出現降低的現象。此外,區內全年氣溫相對較低[16],植物生長期相對較短,致使其生長速率較低,故導致植物生長量積累緩慢,垂穗披堿草種植后,邊坡土壤中可促進植物生長的養分在植物種植后因土壤侵蝕等因素而耗盡,使得植物發生退化,從而導致其根徑變細。
草本植物根系作為一種天然彈塑性材料,其變形包括彈性變形和塑性變形[10],故拉伸模量應采用割線模量(在公式(3)的基礎上,乘以0.4~0.7的折減系數以考慮根系的塑性變形)較為合適,但本研究并未采用割線模量,而是采用彈性模量的計算公式(公式3) 作為垂穗披堿草根系拉伸模量的計算公式,其原因在于不同植物根系其彈性模量和割線模量之間的相關系數相差甚遠[23-24];此外,植物在不同生長階段,其單根抗拉特性指標亦相差甚遠,其原因在于隨生長時間增加,植物根系木質素、纖維素和半纖維素含量均不同。本研究結果表明,生長時間愈長,植物根系拉伸模量和極限拉伸應變率均表現出增加的變化特征,但不同坡向處其增加幅度均不同(圖3),這可能與植物根系木質素、纖維素和半纖維素含量增加有關[7]。相關研究表明,植物生長時間愈長,其內部木質素含量愈高,而木質素含量較高的植物根系拉伸模量則愈高,但不同坡向作用條件下,植物根系木質素、纖維素和半纖維素是否存在差異尚不明確,需在今后的研究中做進一步深入分析。
3.3 坡向對植物根系抗拉特性指標影響
坡向是影響植物生長重要的地形因子之一,其主要通過影響土壤養分、水分、溫度和土壤微生物等生境條件改變植物生長[25-28]。陽坡通常受光照條件影響程度較強,具有較高的地溫,其蒸發相對強烈,故土壤濕度相對較低,而半陰半陽邊坡和陰坡所受光照相對較弱,地溫相對較低,蒸發相對較小,故土壤濕度較大。土壤是植物生長的物質基礎,植物生長發育和分布與不同坡面土壤特征存在密切關系[18]。植物生長與其生存環境之間是一種相互協同進化和適應的關系,植物在不同坡向和立地條件下生長特征會產生差異,亦會因生境不同而產生一些可塑性反應[21]。本研究結果表明相較于取樣位置,坡向對垂穗披堿草根系抗拉特性指標產生顯著性影響,即生長于陽坡生長期為2 a垂穗披堿草根系根徑和單根抗拉力均高于生長期相對更長,且生長于陰坡和半陰半陽邊坡的垂穗披堿草,該結果反映出陽坡相對較為充足的光照促進了垂穗披堿草的生長,這在一定程度上增加了垂穗披堿草根徑,提高了其單根抗拉力,從而有助于提高草本植物的固土護坡能力。陽坡促進植物護坡能力提高的機理可能在于陽坡受到光照直射時間長,使得土壤溫度相對較高[12],從而促進土壤微生物數量增加和活性提高,這可進一步促進植物根際土中營養元素的供給,間接地起到促進植物生長的作用,因此這在很大程度上導致陽坡植物根系相對較發達且根徑較粗,其結果則相應地增加了根系的抗拉力。相比較而言,陰坡和半陰半陽邊坡由于光照相對較弱,土壤溫度較低,根際微生物數量相對較少且活性相對較低,難以為植物生長提供充足的養分,從而導致生長于陰坡或者半陰半陽邊坡的植物根徑相對較細以及抗拉力相對較低的結果。
植物根徑粗細和數量作為體現植物地下部分生物量的2個基本指標,在一定程度可反映其護坡能力[4,22]。本研究未進一步深入探討坡向等因子對植物根系數量的影響,而是主要分析了根系抗拉特性指標值對坡向的響應。由本研究結果可知,區內陽坡草本植物具有相對較粗的根徑(如圖3所示),這與生長于陽坡植物具有較高的生物量基本相一致[18];除此以外,本研究亦發現種植于半陰半陽邊坡植物具有較高的單根抗拉強度(如圖3和圖4所示),造成該結果的原因可能在于生長于陽坡的植物因接受光照相對較強,其根系生長相對較快,導致根徑增加速率超過抗拉力增加速率,從而造成其單根抗拉強度相對較小,而生長于半陰半陽坡植物因接受光照條件相對有限,其根徑增長速率相對滯后于其抗拉力,從而導致其單根抗拉強度相對較大,而生長于陰坡的植物由于接受相對更為有限的光照,其根徑和抗拉力增加速率表現出相對更為緩慢的特征,導致其單根抗拉強度相對較低(如圖4b所示)。同樣地,對于單根極限拉伸應變率,種植于陽坡的植物略微大于對照組和陰坡,該結果說明陰坡相對較低的溫度和較弱的光照條件導致垂穗披堿草根系生長緩慢,造成其拉伸變形能力或延展性不足;對于單根拉伸模量,對照組邊坡最大而陽坡最小,該結果說明了生長于半陰半陽邊坡的垂穗披堿草根系抵抗變形能力最為顯著,而種植于陽坡的垂穗披堿草根系抵御拉伸變形能力最弱,該順序與極限拉伸應變率的結果表現出不一致性,這在一定程度上反映出了陽坡可促進植物根系生長,從而使得植物根徑亦得到增加,而根徑愈粗的植物其變形能力愈弱,故拉伸模量亦表現出愈弱的特征。
4 結論
研究區生長于半陰半陽邊坡和陰坡的垂穗披堿草根系抗拉特性指標,在不同取樣點未表現出顯著性差異;種植于陽坡的垂穗披堿草根系各指標除根徑以外,均表現出顯著性差異;隨生長時間增加,生長于半陰半陽坡和陽坡的垂穗披堿草,其抗拉力、抗拉強度、極限拉伸應變率和拉伸模量等抗拉特性指標則呈增加的變化特征;種植于半陰半陽邊坡的垂穗披堿草根徑、抗拉力、抗拉強度、極限拉伸應變率和拉伸模量均大于陰坡,生長于陽坡的垂穗披堿草根徑和抗拉力均大于半陰半陽坡,而其余各指標則表現出不一致的變化特征。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
