新疆巴楚地區奧陶紀中下統地層中造礁、附礁生物化石的發現及意義*
2022-09-27 00:31:30蘇學龍于國輝艾孜孜伊力哈木
甘肅科技 2022年11期
關鍵詞:生物
蘇學龍,于國輝,艾孜孜·伊力哈木
(喀什大學 生命與地理科學學院,新疆 喀什 844000)
1 概述
生物礁是造礁生物原地聚集形成的一種能夠抵御風浪的塊狀或丘狀生物巖體,對于油氣田勘探有重要意義[1]。塔里木盆地生物礁的分類、形成因素、分布特征一直是石油地質學家關注和研究的重點[2]。前人對奧陶紀生物礁進行了大量研究,并取得可觀成果,如李越等[3-4]初步總結了寒武紀到奧陶紀輻射演化的過程中海洋造礁生物也得到發展,此過程經歷了瓶筐石—苔蘚蟲—石海綿動物型和珊瑚—層孔海綿動物型兩次標志性事件,且在塔里木地區也有印證;李明明等[5]對巴楚生物礁研究發現,中上奧陶統階段巴楚地區生態環境非常適宜生物礁的發育及海洋居礁生物的生存;蔡習堯等[6]、徐康等[7]、Meng等[8]研究表明在巴楚隆起區內斷裂帶方向大致為NW-SE其中含大量生物建隆和生物層礁,且由達吾孜塔格向唐王城方向減少最終被生物灘取代。地層中的主要造礁生物為苔蘚蟲、海綿累、棘皮類動物、Calathium(瓶筐石)類,附礁生物主要是雙殼類、腹足類、腕足類、三葉蟲,生物礁的產生和發展受到這些生物礁群落的控制。
奧陶紀碳酸鹽巖地層在巴楚一間房、良里塔格、大坂塔格等斷層中出露較好,該地區也是研究奧陶紀古生物和古地理的重要地區,但對于化石屬種特征、分布反映的古鹽度、古水溫、古生態等方面的研究較少,此類研究對沉積相和沉積環境具有重要意義。基于此本研究在前人研究的基礎上,結合在巴楚一間房附近和大坂塔格采集的化石標本,對化石外部形態、組合及其指示的環境意義進行了深入研究,旨在加強巴楚地區奧陶紀地層中部分地質現象的解釋,為塔里木盆地奧陶紀生物礁環境及古生態提供寶貴證據。
2 地質背景及地層情況
巴楚隆起位于塔里木盆地西北緣,其構造總體受塔里木盆地構造控制。從晚震旦紀到奧陶紀盆地構控制。從晚震旦紀到奧陶紀盆地構造為被動陸緣盆地階段,盆地前后經歷了加里東、海西、印之—燕山、喜馬拉雅構造運動,其中加里東構造運動對巴楚隆起區的奧陶紀碳酸鹽巖古地理和沉積環境起重要作用[9]。且奧陶紀全球暖水臺地分布廣泛,生物大爆發的背景使得塔里木盆地中部隆起區中上奧陶統發育大量暖水礁灘及大量古生物群,是當時海洋生態和生物演化的有力證據[10]。
研究區位于巴楚東北部,見圖1。通過對采集化石組合和巖性分析,大坂塔格發現的化石產自鷹山組,良里塔格采集的化石產自一間房組。由于奧陶紀巴楚隆起的抬升吐木休克組與一間房組部分地層缺失,一間房組下部鷹山組之間也部分缺失。一間房組巖性較單一,為灰色灰巖、棘屑灰巖、生屑泥晶灰巖、瓶筐類礁灰巖,局部夾砂屑灰巖,化石主要為棘屑,托盤類,少量藻屑、鈣質骨針、三葉蟲、腕足等,可劃分為Pygod us serra 帶、Eolacognath us crassus 帶、Lenodus variabilis 帶、Microzarkodina parva四個牙形石帶。鷹山組巖性為淺褐、褐灰、灰色灰巖為主,夾灰色灰質白云巖、白云質灰巖,劃分為Paroistodus originalis—Serratognathus chuxianensis帶和Serratog nathusdiversus—Paroistodus proteus 帶兩個牙形石帶[11]。
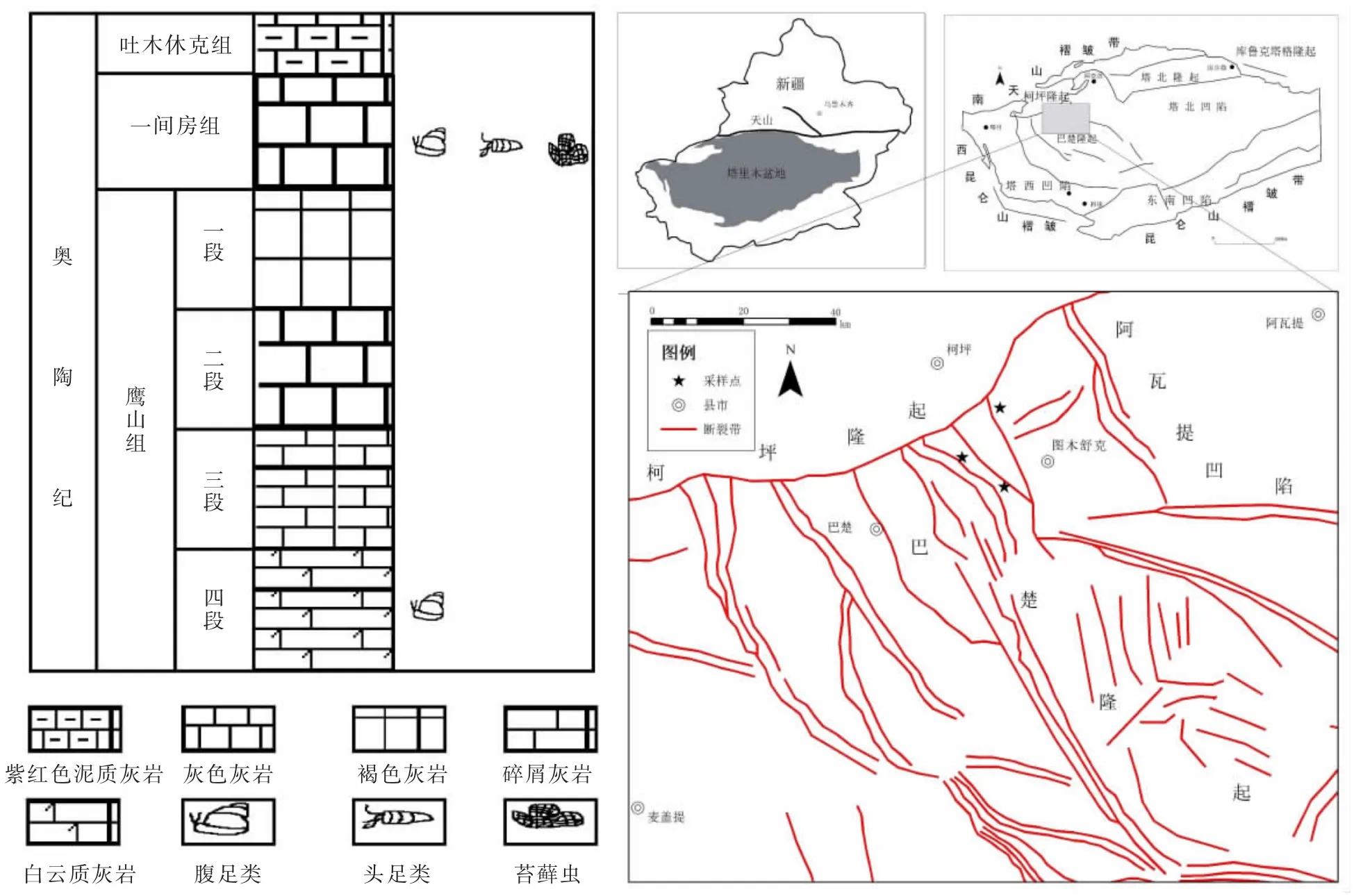
圖1 化石發現點位置圖及巖性柱
2 化石形態及埋藏特征
位于巴楚縣東北部圖木舒克市附近的一處剖面上,出露部分巖性為灰色厚層狀泥晶灰巖及生物碎屑灰巖的巖層,有明顯生物作用,中間穿插有硅質透鏡狀條帶,大小不一。1991年張師本等人對該處地層進行描述,將其劃分為丘里塔格群上亞群[12]。此處剖面中產有大小均一的腹足類化石,多嵌入層狀生物礁中,種類單一,數量較多,殼體較為完整,化石基本為殼頂朝上,或略微斜上埋藏于地層中,可將其劃分為原地埋藏,周圍有棘皮碎屑,牙形石等。此部分地層與東南部大坂塔格地層不一致,但腹足類化石相似程度比較高,見圖2—圖5。

圖2 化石發現地層野外照片

圖3 鷹山組腹足類化石圖片

圖4 一間房組苔蘚蟲化石圖片

圖5 一間房組化石圖片
本次研究的鷹山組地層出露部分為50 m,以白云質灰巖為主,地層中產一屬一種腹足類化石Liospira biconvexa.,豐度較高。化石基本都被白云石化,大小均一,呈中等凸起的圓形,直徑在2~4 cm,高為3 cm左右,殼體表面比較光滑,有4個逐漸變大的螺環組成,螺環上側面較為狹圓,臍孔幾乎全為殼質包裹。由于化石經過強置換作用化石細微構造沒有保存,僅保存外部形態,化石的內外唇、殼口、側面特征及內部結構不詳。化石保存為殼頂指向上、斜上,殼頂朝下等多種保存狀態。與上述地層腹足類化石保存方式不同,此地發現的螺殼保存完整程度不佳,僅個別保存完好,其余多為碎殼,周圍不見其他化石僅存在一些棘皮碎屑(圖3a-圖3d)。
一間房組地層野外出露5 m,巖性比較單一,為一套深灰色灰巖,向西北方向有生物礁出現,上部為紫紅色泥質灰巖。苔蘚蟲化石Batostoma sp.集中出現在一間房組下段,呈水平分布。化石硬體呈盤狀、塊狀,口室直徑在1~3 mm,呈圓角六邊形,排列不規律,外帶較窄,中間區體壁較薄,向外體壁變厚,本屬種的一大特點為間隙孔甚少。從野外的埋藏特征來看其外形多為近似球狀,成簇出現,從橫切面看內部結構重結晶嚴重,其蟲室和內帶構造不可見(圖4a-圖4h)。另外產苔蘚蟲化石的地層中產種類豐富的頭足類,具體有以下分子:葉松卷角石(比較種)Aphetceras Lobatum cf.一件(圖5c),櫛角石(未定種)Cetnoceras Noetling sp.一件,雙房角石(未定種)Dideroceras sp.兩件(圖5a-圖5b)。地層中還發現部分三葉蟲,托盤類(圖5d),其化石密度不大。
3 討論
3.1 腹足類化石分布成因及意義
研究區鷹山組地層為淺水碳酸鹽巖沉積,其中發現的Liospira biconvexa.同樣大小的個體埋藏在一起,分選性較好(圖3),碎殼和其他生物碎屑為在強水流沖刷或遠距離運輸過程中發生破碎導致。且化石埋藏形態多樣,為典型的異地埋藏特征。上述化石埋藏特征均表示當時臺地水流的比較湍急。早奧陶紀沿襲了寒武紀海進過程,一直持續到中—晚奧陶紀達到頂峰[13],此次鷹山組發現的生物碎屑灘也證實了這一點。通過與現代海洋生物對比發現,古海洋深水區域底棲的腹足類生物普遍偏小,本次發現臺地上的腹足類也為2~3 cm的小形圓殼形狀,生前應生活在水位較深地區,且在距離該地層不遠的生物礁中也發現類似腹足類化石,大小與之一致,但此處地層為深灰色灰巖和透鏡狀生物礁,為較深的水域沉積特征,依據張師本等[14]對該套地層時代的厘定,應為一間房組下段。前人的研究也證明塔里木盆地奧陶紀生物礁的附礁生物以腹足類為主,由此推測,鷹山組中發現的腹足類化石為臺地邊緣生物礁死去的腹足類螺殼被高能海水攜帶到開闊臺地上沉積下來。
3.2 苔蘚蟲化石指示的生態意義
苔蘚蟲為一類固著生物,附著于較硬的底質上,不同的生存環境對其形狀和分布狀況有影響。苔蘚蟲對于溫度及海水的適應性較強,其生活緯度較廣從赤道到兩極均有分布[15],由圖6可知奧陶紀時期赤道穿過塔里木盆地,巴楚地區發現的苔蘚蟲應為喜溫性物種,屬種組成單一,但相對豐度高。通過比較,研究區發現的屬種在北美、西伯利亞、烏拉爾山、西歐、中國湖北均有分布,古地理上形成一個溫帶、熱帶海域的緯度梯度,時間跨度從早奧陶紀到晚志留紀地層中均有其身影[16]。本研究采集的化石硬體結構多為塊狀、圓球狀和殼狀(圖4a-圖4d),說明當時海水比較湍急,浪擊頻繁,水體清澈。化石整體分異程度不高,指示當時海水鹽度適中。底質對于苔蘚蟲的生長和發展也是一個重要的因素,從圖4g看出,其生長以其本身硬體為底質,多個個體聚集發展,圖4h是以吸附灰泥為底質,上述固著生長特點為抵抗浪擊的特征,且其自身不斷向上發展累積成礁,發展動力應為水中浮游生物和陽光,生長在水下1~5 m[17],這種現象在熱帶水域非常明顯。從產于附近的其他種類生物來看,棲息在此處的生物有附礁生存的腹足類,游覓在海平面以下的頭足類,以及其他一些造礁的托盤類和海百合,均指示鹽度正常、清澈,光線條件好的的淺水環境,組成以Batostoma為優勢分子的群落生境類型,該類型與肖傳桃等[18]1993年報道的江漢平原隆礁生態系統比較相似,此類型應為臺地邊緣礁丘式生態系統(圖6b)。
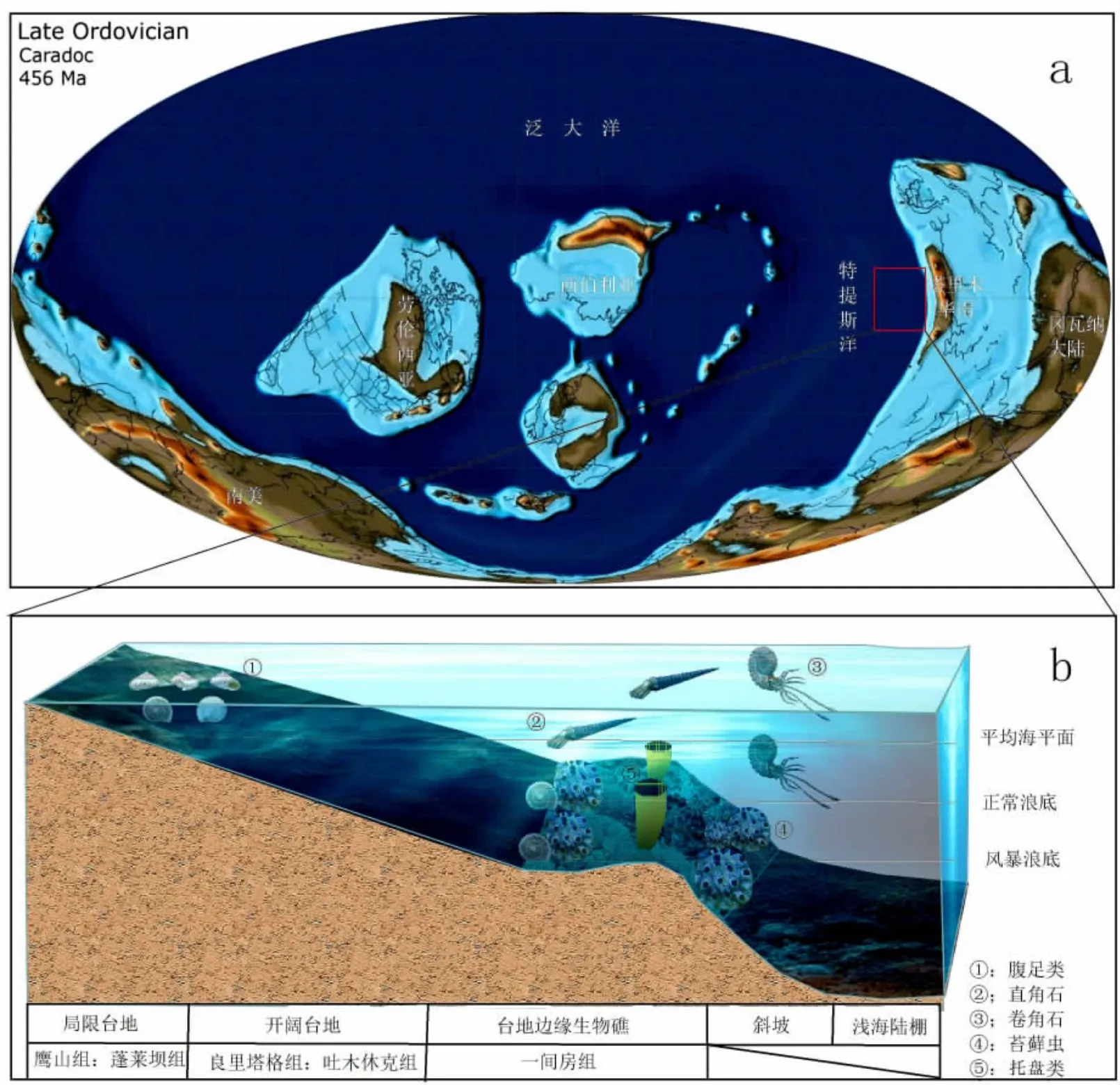
圖6 研究區生態示意圖(據Scotese,C R)
4 結論
(1)通過化石組合和巖性分析,大坂塔格發現的化石產自鷹山組,良里塔格采集的化石產自一間房組,通過生態系統對比,研究區為臺地邊緣礁丘式生態系統。
(2)巴楚鷹山組出露的豐度比較高的Liospira biconvexa.埋藏形態各異,殼頂指向不規律,系異地埋藏,代表較強的水動力條件,其圍巖白云巖含量較高,化石白云石化作用顯著,可見化石形成過程中置換速度小于溶解速度,指示比較強的沖刷環境,其化石來源極有可能為臺地邊緣死亡腹足類螺殼。
(3)一間房組的Batostoma sp.化石分異程度小,數目較多,硬體呈球狀、殼狀,指示比較動蕩清澈的水體環境,苔蘚蟲的棲息底質有一定的差異,表明奧陶紀早期海進,使得水體動力較足,比較動蕩,這一反應在古生物特征得到體現。當時半局限臺地和臺地邊緣生物礁所處環境為溫暖清澈水,流比較湍急的淺水環境。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
