2019-2020年夏季舟山海域浮游動物優勢種生態位及其生態分化
2022-09-25 09:46:16宋晨孟周王曉波韓慶喜
海洋學報 2022年10期
宋晨,孟周,王曉波,韓慶喜*
( 1. 寧波大學 海洋學院,浙江 寧波 315832;2. 國家海洋局寧波海洋環境監測中心站,浙江 寧波 315012;3. 禹治環境科技(浙江)有限公司,浙江 寧波 315199)
1 引言
浮游動物是海洋中的次級生產者,其種類、分布和數量均可影響海域生產力,對漁業發展意義重大[1-2]。浮游動物的群落結構、生物量和優勢種分布可以反映水域環境的變化,常作為水域環境的指示動物[3]。浮游動物因其隨波逐流的特性而受到諸多因素的影響。例如,王慧君等[4]發現,不同水團會對浮游動物群落結構地理分布格局產生影響;何瑞等[5]指出,浮游植物可通過食物網影響浮游動物種類組成和分布;付顯婷等[6]指出,水環境變化會對浮游動物群落結構分布起主導作用。生態位是指種群在生態系統中所占據的時空位置以及它與其他種群之間的相互影響[7-8]。生態位理論常用于生物多樣性、種間關系、群落的結構和演替及種群進化等方面的研究[9-10],亦廣泛運用于海洋生態系統,常用于研究海域內生物的種群特征、生存狀況、利用各種資源的幅度及種對間的相互關系等[11]。
舟山海域位于浙江省北部,受長江和錢塘江兩大河口及臺灣暖流和沿岸寒流交匯的影響,水域環境復雜,水團交匯帶來大量餌料,是我國舟山漁場的主要海區。然而,由于20世紀70年代以來無節制的捕撈等原因,使舟山漁場漁業資源減少,此外,人類活動愈加頻繁,海域環境質量下降[12],使該海域浮游動物生物量呈下降趨勢[13]。近年來,“綠水青山就是金山銀山”的呼吁深入人心,合理有效地開發海洋資源及對環境的保護越來越得到重視。
目前對舟山海域浮游動物的研究主要集中在分布特征、群落結構等方面[1,4,13-14],而該海域浮游動物優勢種生態位的研究鮮有報道。此外對浮游動物的研究主要針對季節變化[3,9-10],對年際變化鮮有報道。浮游動物生態位分布在一定程度上能反映出海洋生態環境的變化現狀[9],因此研究浮游動物的生態位和年際變化具有重要意義。有鑒于此,根據2019年夏季和2020年夏季對舟山海域浮游動物的采樣調查,分析了舟山海域浮游動物優勢種組成及生態位的主要特征,并采用冗余分析(RDA)探究環境因子變化對浮游動物優勢種生態位分化的影響,該研究將加深對舟山海域浮游動物群落的認識程度,有望進一步了解該海域浮游動物群落中不同種群的相互作用關系,為闡述浮游動物應對環境變化的適應機制等研究提供數據基礎和科學依據。
2 材料與方法
2.1 樣品采集與分析
根據舟山海域的海域特點和本次研究目的,共設置相同采樣站位12個(圖1),調查范圍為舟山海域(29°~31°N,122°00′~123°30′E),Z1、Z3、Z5、Z6、
Z7、Z9、Z11站位分布在舟山近岸,Z2、Z4、Z8、Z10、Z12站位分布在舟山近海,租用漁船分別在2019年7月24-28日、8月14日和2020年7月26-31日及8月7日進行浮游動物和水質參數的調查采樣。采用淺水Ⅱ型浮游動物網具(網口內徑為31.6 cm,網長為140 cm,網口面積為0.08 m2),從底層至表層垂直拖網采集浮游動物樣品。將樣品用5%的福爾馬林固定之后,帶回實驗室去除雜質,在顯微鏡(奧林巴斯,SZX16,日本)和解剖鏡(奧林巴斯,SZ61,日本)下進行鑒定和統計。樣品采集和分析按《海洋調查規范》(GB/T 12763.6-2007)[15]進行。
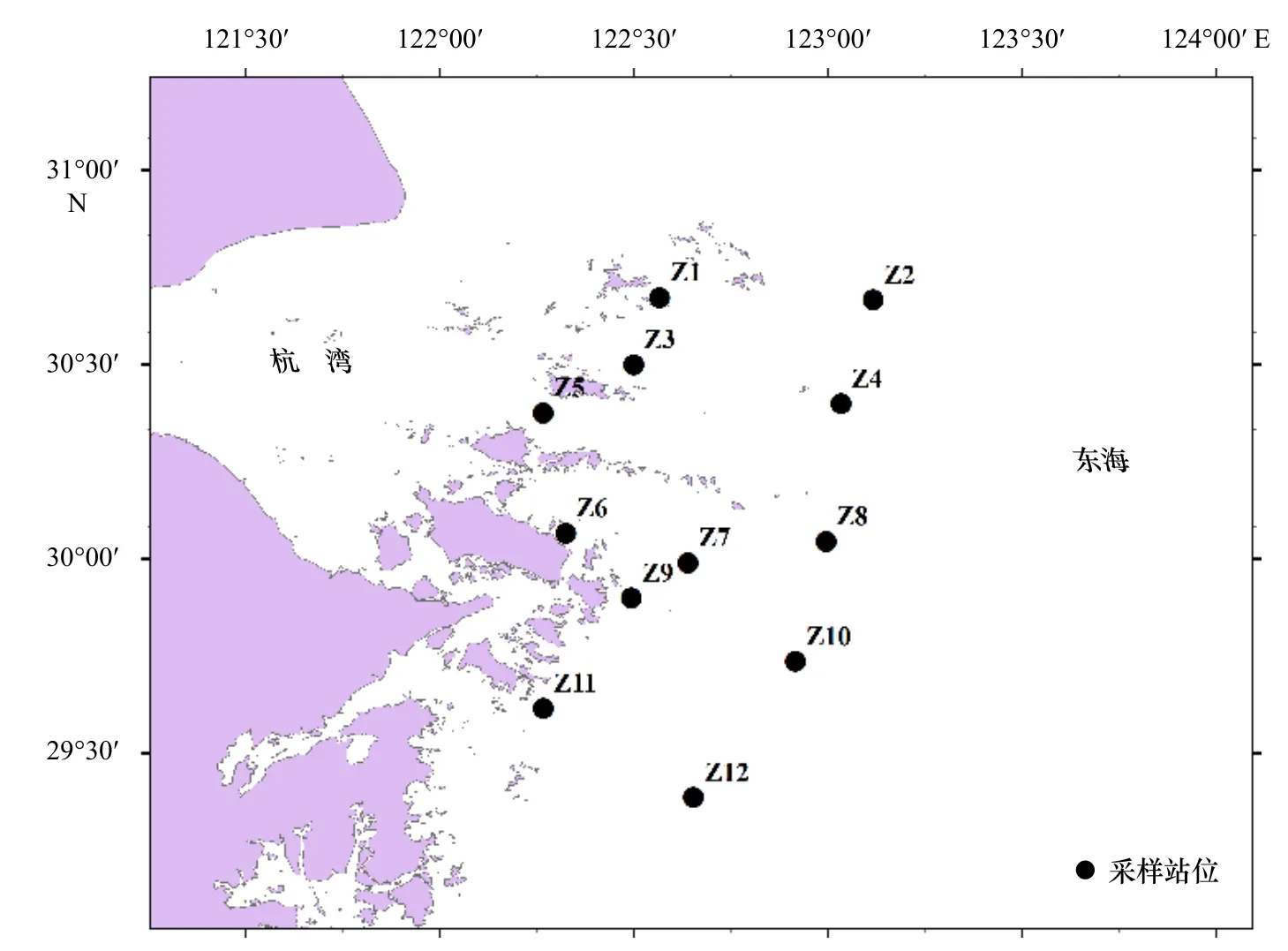
圖1 2019-2020年夏季舟山海域浮游動物采樣站位Fig. 1 Zooplankton sampling stations in the Zhoushan waters during the summer of 2019-2020
使用船載測深儀測量水深;使用溫鹽深儀CTD測量水溫和鹽度;使用pH計法測量pH;采用碘量法測量溶解氧(DO)濃度;采用熒光分光光度法測量葉綠素a(Chla)濃度;采用過濾物重量法測量懸浮物(SS)濃度;采用堿性高錳酸鉀法測量化學需氧量(COD);采用磷鉬藍分光光度法測量活性磷酸鹽(PO4-P)濃度;采用奈乙二胺分光光度法測量亞硝酸鹽-氮(NO2-N)濃度;采用鋅-鎘還原法測量硝酸鹽-氮()濃度;采用次溴酸鹽氧化法測量氨-氮濃度;采用硅鉬藍法測量活性硅酸鹽(SiO3-Si)濃度。此外,總無機氮(TIN)濃度等于亞硝酸鹽-氮、硝酸鹽-氮與氨-氮的濃度和。水樣采集與分析按《海洋監測規范》(GB 17378.4-2007)[16]進行。
2.2 數據分析
2.2.1 優勢種分析
采用優勢度指數(Y)[17]判斷浮游動物優勢種:

式中,ni為 第i種浮游動物的個體數;N為所有浮游動物種類總個體數;fi為該種出現的站位數與總站位數的比值。當Y>0.02[10]時,該種為優勢種。
2.2.2 優勢種更替率
優勢種更替率計算公式(S)[10]為
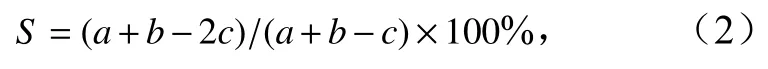
式中,a為2019年浮游動物優勢種種類數;b為2020年浮游動物優勢種種類數;c為兩年共同優勢種種類數。
2.2.3 生態位寬度
Shannon生態位寬度(Bi)計算公式[18]為
式中,Bi的取值范圍為[0,R];Pij為i種在j站位的個體數與該物種總個體數的比值;R為總站位數。
2.2.4 生態位重疊
Pianka指數生態位重疊指數(Oik)計算公式[19]為
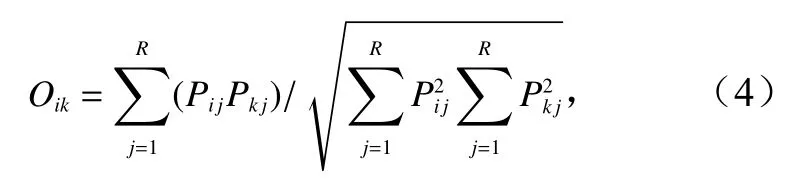
式中,Oik的取值范圍為[0, 1];Pij和Pkj分別為i種和k種在j站位的個體數占該物種總個體數的比值;R為總站位數;當Oik>0.6,視為顯著重疊[20],當0.3<Oik<0.6時,視為重疊有意義[11]。
2.3 數據處理
使用ArcGIS Desktop 10.6制作采樣站位分布圖;使用Microsoft Excel 2019計算優勢度指數和優勢種更替率;使用R4.2.1軟件計算生態位寬度值和生態位重疊值;使用SPSS 18.0 進行兩獨立樣本t檢驗;使用Canoco 5.0 進行冗余分析(RDA)。
在統計分析之前對除pH外的環境因子進行對數(lg(x+1))轉換[21],使其符合正態分布,然后對浮游動物優勢種數據進行去趨勢對應分析(DCA)。當排序軸最大梯度小于3時,使用冗余分析;排序軸最大梯度大于4時,使用典范對應分析(CCA);當排序軸最大梯度大于3小于4時,二者均可使用。結果顯示,兩年排序軸最大梯度均小于3,因此選擇RDA。
3 結果
3.1 浮游動物群落結構及優勢種
2019-2020年夏季在舟山海域共采集到浮游動物8個大類共130種,2019年采集到浮游動物103種,其中橈足類43種(41.7%)、浮游幼蟲16種(15.5%)、水螅水母類11種(10.7%)、毛顎類3種(2.9%)、多毛類3種(2.9%)、原生動物1種(1.0%)、背囊類1種(1.0%)、其他動物25種(24.3%);2020年采集到浮游動物82種,其中橈足類27種(32.9%)、浮游幼蟲16種(19.5%)、水螅水母類12種(14.6%)、毛顎類5種(6.1%)、多毛類2種(2.5%)、其他動物20種(24.4%)。2019年浮游動物平均豐度為3 866.6 ind./m3,Z5站最高(10 205.8 ind./m3),Z11站最低(1 164.4 ind./m3),舟山近岸海域平均豐度為4 521.1 ind./m3,舟山近海平均豐度為2 950.4 ind./m3;2020年浮游動物平均豐度為2 269.2 ind./m3,其中Z11站最高(8 452.2 ind./m3),Z9站最低(573.8 ind./m3)舟山近岸海域平均豐度為2 397.4 ind./m3,舟山近海平均豐度為2 089.6 ind./m3。
根據優勢度指數(Y>0.02),舟山海域共有浮游動物優勢種12種,其中2019年有優勢種6種,分別是針刺真浮螢(Euconchoecia aculeata)、針刺擬哲水蚤(Paracalanus aculeatus)、夜光蟲(Noctiluca scintillans)、橈足類橈足幼體(copepoda copepodite)、橈足類無節幼體(copepoda nauplius)、強額孔雀水蚤(Parvocalanus crassirostris);2020年10種,分別是刺尾紡錘水蚤(Acartia spinicauda)、肥胖箭蟲(Flaccisagitta enflata)、擬長腹劍水蚤(Oithona simills)、強額孔雀水蚤、橈足類橈足幼體、橈足類無節幼體、刷狀螢蝦(Lucifer penicillifer)、微駝隆哲水蚤(Acrocalanus gracilis)、針刺擬哲水蚤和中華哲水蚤(Calanus sinicus)。兩年共同優勢種(類)4種,分別為針刺擬哲水蚤、橈足類橈足幼體、橈足類無節幼體和強額孔雀水蚤(表1)。2019年、2020年優勢種更替率為66.67%。
3.2 優勢種的生態位寬度
2019-2020年夏季舟山海域浮游動物優勢種的生態位寬度范圍為0.45~2.34,根據生態位寬度值的大小將其劃分為3段,第1段浮游動物優勢種Bi≥1.60,為廣生態位種;第2段浮游動物優勢種 0.8≤Bi<1.60,為中生態位種;第3段浮游動物優勢種Bi<0.8,為窄生態位種。
2019年夏季舟山海域浮游動物優勢種的生態位寬度范圍為1.47~2.27,其中針刺真浮螢(1.60)、強額孔雀水蚤(1.75)、橈足類無節幼體(2.23)和橈足類橈足幼體(2.27)為廣生態位種,針刺擬哲水蚤(1.47)和夜光蟲(1.55)為中生態位種,無窄生態位種(表2)。廣生態位種和中生態位種分別占全部優勢種的66.67%和33.33%。
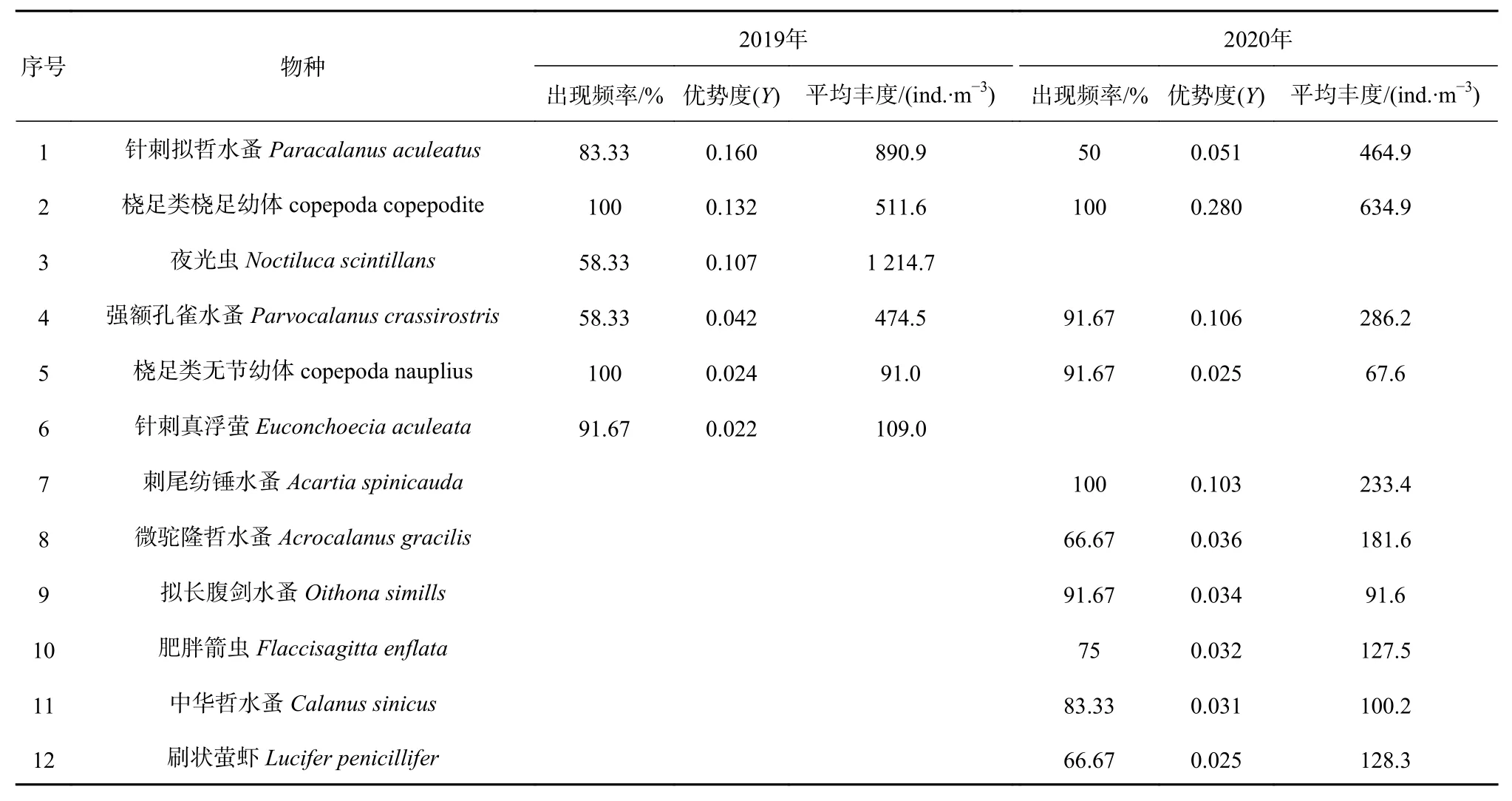
表1 2019年、2020年夏季舟山海域浮游動物優勢種Table 1 Dominant zooplankton species in the Zhoushan waters in summer 2019 and 2020
2020年夏季舟山海域浮游動物優勢種的生態位寬度范圍為0.45~2.34,其中微駝隆哲水蚤(1.61)、強額孔雀水蚤(1.73)、肥胖箭蟲(1.74)、橈足類無節幼體(1.82)、中華哲水蚤(1.92)和橈足類橈足幼體(2.34)為廣生態位種,刺尾紡錘水蚤(1.25)和擬長腹劍水蚤(1.41)為中生態位種,針刺擬哲水蚤(0.45)和刷狀螢蝦(0.76)為窄生態位種(表3)。廣生態位種、中生態位種和窄生態位種分別占全部優勢種的60.00%、20.00%和20.00%。
共同優勢種中,除針刺擬哲水蚤外,其他3種優勢種生態位寬度值變化不大,仍為廣生態位種,僅針刺擬哲水蚤生態位寬度值大幅度下降,由2019年的中生態位種(1.47)變為2020年的窄生態位種(0.45)。
3.3 生態位重疊
2019-2020年夏季舟山海域浮游動物優勢種的生態位重疊指數范圍為0~0.98。2019年浮游動物優勢種生態位重疊指數范圍為0.11~0.79,其中生態位顯著重疊(Oik≥0.6)有5種對,占33.33%,分別為橈足類橈足幼體和橈足類無節幼體(0.79)、橈足類橈足幼體和強額孔雀水蚤(0.60)、橈足類無節幼體和強額孔雀水蚤(0.63)、橈足類無節幼體和針刺擬哲水蚤(0.68)、針刺擬哲水蚤和強額孔雀水蚤(0.72)。生態位重疊有意義(0.3≤Oik<0.6)的有6種對,占46.67%,其中橈足類橈足幼體和針刺真浮螢(0.58)、橈足類橈足幼體和針刺擬哲水蚤(0.59)之間重疊指數均接近0.6。其余3個種對的生態位重疊指數均小于0.3,占20%(表2)。
2020年浮游動物優勢種生態位重疊指數范圍為0~0.98,生態位顯著重疊(Oik≥0.6)有11種對,占24.45%。其中微駝隆哲水蚤和刷狀螢蝦(0.87)、擬長腹劍水蚤和刺尾紡錘水蚤(0.96)、擬長腹劍水蚤和針刺擬哲水蚤(0.97)、刺尾紡錘水蚤和針刺擬哲水蚤(0.98)之間的生態位重疊指數達到了0.85以上。生態位重疊有意義(0.3≤Oik<0.6)的有11種對,占24.45%,其中橈足類橈足幼體和強額孔雀水蚤(0.58)之間重疊值接近0.6。其余23個種對的生態位重疊指數均小于0.3,占51.10%,其中微駝隆哲水蚤和針刺擬哲水蚤(0)、刷狀螢蝦和針刺擬哲水蚤(0)生態位重疊指數為0(表3)。
3.4 生態位分化
3.4.1 舟山海域環境因子
2019-2020年夏季舟山海域主要環境因子存在顯著差異(表4),其中2019年溫度、pH和Chla濃度顯著高于2020年(p<0.05),而2020年SS和SiO3-Si濃度則顯著高于2019年(p<0.05),其余各環境因子差異不顯著(p>0.05)。
3.4.2 冗余分析
利用前向選擇和蒙特卡羅檢驗對各環境因子進行篩選(p<0.05),根據篩選結果(表5),2019年篩選出SiO3-Si濃度、COD、溫度和Chla濃度進行排序分析,2020年篩選出SiO3-Si濃度和DO濃度進行排序分析(圖2)。
2019年篩選出4個環境因子與6個浮游動物優勢種進行RDA分析,分析結果如下:RDA1特征值為0.587 5,解釋物種變量為58.75%,RDA2特征值為0.232 6,解釋物種變量為23.26%,2個排序軸4個環境因子的累積解釋物種變量為82.01%。2019年浮游動物生態位分化主要受SiO3-Si濃度、COD、溫度和Chla濃度的影響。
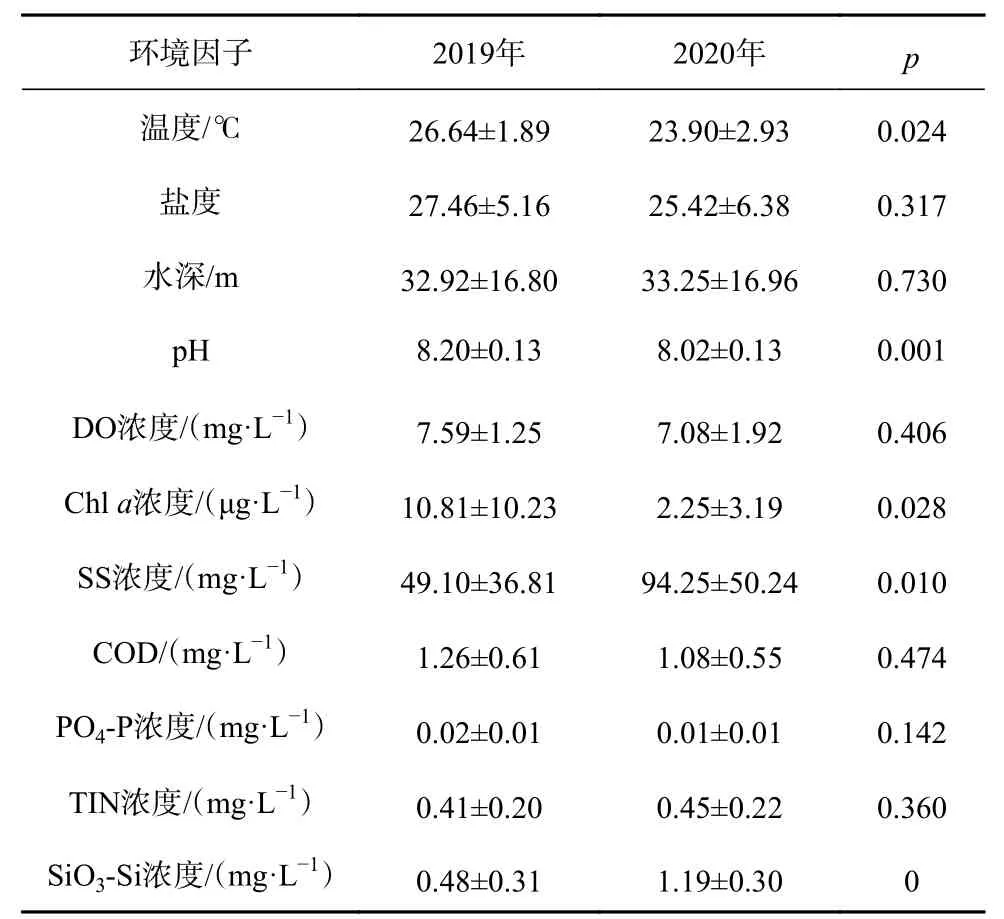
表4 2019年、2020年夏季舟山海域環境因子Table 4 Environmental factors of Zhoushan waters in summer 2019 and 2020
2020年篩選出2個環境因子與10個浮游動物優勢種進行RDA分析,分析結果如下:RDA1特征值為0.739 2,解釋物種變量為73.92%,RDA2特征值為0.147 4,解釋物種變量為14.74%,2個排序軸2個環境因子的累積解釋物種變量為88.66%。2020年浮游動物生態位分化主要受SiO3-Si濃度和DO濃度的影響。
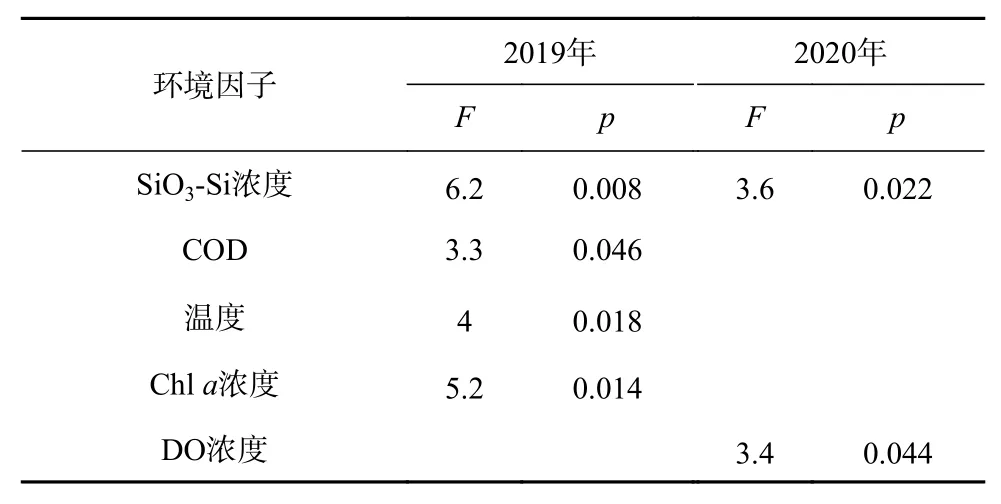
表5 環境因子篩選結果Table 5 Results of the selection on environmental factors
3.4.3 RDA排序圖
由圖2可知,2019年針刺真浮螢、針刺擬哲水蚤、橈足類橈足幼體、橈足類無節幼體和強額孔雀水蚤的分布主要受COD、溫度、Chla濃度和SiO3-Si濃度的影響,其中針刺擬哲水蚤、橈足類橈足幼體、橈足類無節幼體和強額孔雀水蚤分布與COD、溫度、Chla濃度和SiO3-Si濃度呈正相關,針刺真浮螢分布與COD、溫度、Chla濃度和SiO3-Si濃度呈負相關,夜光蟲分布與COD和Chla濃度呈正相關,與SiO3-Si濃度呈負相關,與溫度相關性較低。
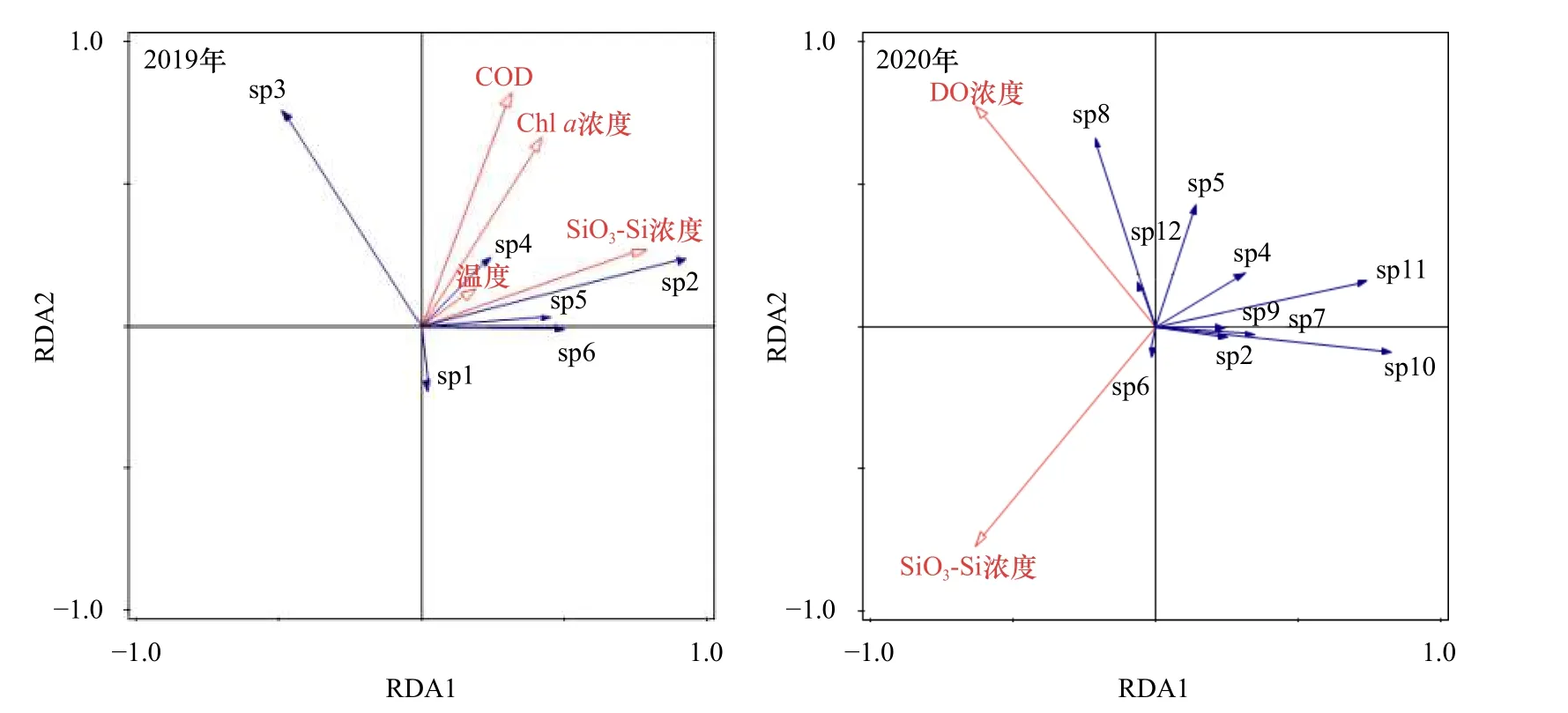
圖2 2019-2020年夏季舟山海域浮游動物優勢種冗余分析排序圖Fig. 2 RDA ordination graphs of the dominant zooplankton species in the Zhoushan waters in summer 2019-2020
2020年,針刺擬哲水蚤、橈足類橈足幼體、橈足類無節幼體、強額孔雀水蚤、刺尾紡錘水蚤、肥胖箭蟲、擬長腹劍水蚤、刷狀螢蝦、微駝隆哲水蚤和中華哲水蚤的分布主要受SiO3-Si濃度和DO濃度的影響,其中橈足類無節幼體、肥胖箭蟲、中華哲水蚤的分布與DO濃度呈正相關,除強額孔雀水蚤的分布與SiO3-Si濃度呈正相關外,其余優勢種的分布均與SiO3-Si濃度呈負相關。
4 討論
4.1 浮游動物群落結構及優勢種分析
本研究中,浮游動物群落組成均為橈足類最多,其次依次為浮游幼蟲和水螅水母類,這種群落結構與孟慶超和趙淑江[22]、駱鑫等[13]的研究結果相似。2020年浮游動物物種數、平均豐度均低于2019年,該結果與許卓昀等[23]發現“舟山海域浮游生物種類逐年減少、豐度逐年上升”的研究結果不完全一致,其原因可能與2019年夜光蟲暴發有關,其平均豐度遠高于其他優勢種,使得2019年浮游動物豐度高于2020年。兩年舟山近海浮游動物平均豐度低于舟山近岸海域,這種研究結果與陳小慶等[24]研究結果相一致。
2019年舟山海域浮游動物優勢種共6種,分別是針刺真浮螢、針刺擬哲水蚤、夜光蟲、橈足類橈足幼體、橈足類無節幼體、強額孔雀水蚤,平均優勢度約為0.08。2020年浮游動物優勢種共10種,分別是刺尾紡錘水蚤、肥胖箭蟲、擬長腹劍水蚤、強額孔雀水蚤、橈足類橈足幼體、橈足類無節幼體、刷狀螢蝦、微駝隆哲水蚤、針刺擬哲水蚤和中華哲水蚤,平均優勢度約為0.07。已有的研究表明,浮游動物優勢種越多且優勢度越小,則群落結構越復雜、穩定[25],簡單、脆弱的群落結構會給該海域海洋生態系統的穩定和健康帶來威脅[26]。本研究與侯朝偉等[21]2018年夏季在煙臺近海海域獲得浮游動物優勢種11種(平均優勢度0.07)、李浩然等[27]2009年夏季在黃河口及鄰近海域獲得浮游動物優勢種5種(平均優勢度0.11)、葉文建等[2]2014年夏季在舟山島附近海域獲得浮游動物優勢種6種(平均優勢度0.11)相比,煙臺近海海域浮游動物優勢種群落結構最復雜穩定,其次為舟山海域,且隨著年份增加,結構的復雜性、穩定性增加,最次為黃河口及其鄰近海域,群落結構相對簡單、脆弱。2019年和2020年共同優勢種共4種,分別為針刺擬哲水蚤、橈足類橈足幼體、橈足類無節幼體和強額孔雀水蚤。2019年、2020年優勢種的更替率為66.67%,更替率處于較高水平,說明2019-2020年舟山海域浮游動物的優勢種組成存在較為明顯的年際差異[21]。
4.2 生態位寬度分析
生態位寬度指物種可以利用的所有資源的總和,反映出其對生境的適應程度和對資源的利用能力[28-29]。2019-2020年舟山海域浮游動物優勢種的生態位寬度范圍為0.45~2.34,存在較大的差異,根據生態位寬度值的大小,可將其劃分為廣生態位種(Bi≥1.60),對環境有廣泛適應能力;中生態位種(0.8≤Bi<1.60),對環境的適應能力稍差;窄生態位種 (Bi<0.8),對環境的要求較高[21]。2019年和2020年浮游動物優勢種中,窄生態位種的占比分別為0%和20%,中生態位種的占比分別為33.33%和20%,廣生態位種的占比分別為 66.67%和60.00%,表明廣生態位種是舟山海域浮游動物優勢種的主要組成成分,這與侯朝偉等[21]在煙臺近海海域的研究結果相似。廣生態位種主要對空間因素做出響應,窄生態位種主要對環境因素做出響應[6],當環境發生變化時,物種對生境的適應程度和對資源的利用能力也會發生變化[21]。在4種共同優勢種中,橈足類橈足幼體、橈足類無節幼體和強額孔雀水蚤的生態位寬度值發生浮動,這3個物種生態位寬度值在2019年均占據前列,雖經歷變化但仍屬于廣生態位種,僅針刺擬哲水蚤的生態位寬度值大幅度下降,由2019年的中生態位種(1.47)變為2020年的窄生態位種(0.45)。分析其原因,強額孔雀水蚤為近海廣布種,其適合的溫度為5~33℃[30],舟山近海溫度的變化仍在其適合的溫度范圍內,夏季比較適宜浮游幼蟲的生存[31];因此,它們生態位寬度變化不大。針刺擬哲水蚤為近岸暖水種,2019-2020年溫度下降較為顯著,對針刺擬哲水蚤造成一定的影響,使其對舟山近海環境的適應程度和資源的利用能力下降,從而使其生態位寬度下降。同樣的情況發生在針刺真浮螢,其雖在2019年為廣生態位種(1.60),但其為暖水種,對溫度變化極為敏感[32],溫度的顯著下降使得該種大量減少。由此,廣生態位種一般更容易適應環境的改變,使得其競爭力增強從而更容易取得優勢地位,生態位寬度值在一定程度上可以判定物種的優勢度大小,該研究結果與前人的研究結果[10,33]相似。
4.3 生態位重疊分析
生態位重疊指數可以衡量物種間對資源環境利用的相似度和競爭性[33]。本研究中,2019年浮游動物優勢種Oik≥0.6的占33.33%,2020年Oik≥0.6的占24.44%,種對間顯著重疊現象不廣泛。梁淼等[10]和侯朝偉等[21]研究發現,廣生態位種之間往往生態位重疊程度也較高,但本研究結果并未發現這一現象,如2020年的橈足類橈足幼體(Bi=2.34)和中華哲水蚤(Bi=1.92)之間的生態位重疊指數僅為0.56,重疊不顯著。劉坤等[34]研究表明,環境資源的高異質性、物種的斑塊狀分布、種群生態學特性差異等因素,均會導致不同生態位寬度的物種間產生不同程度的生態位重疊。因此,僅靠生態位寬度值判斷物種間的競爭性可能存在一定的偏差。
2019年生態位重疊指數最高的為橈足類無節幼體和橈足類橈足幼體(0.79),橈足類發育分為無節幼體和橈足幼體兩個階段[35],二者之間關系密切,對環境的選擇重合度較高。2020年生態位重疊指數最高的為針刺擬哲水蚤和刺尾紡錘水蚤(0.98),從生態類群看,二者均為暖水近岸低鹽種[36-38],生存環境相似,資源不足時種間易產生較激烈的競爭。2020年微駝隆哲水蚤和刷狀螢蝦(0.87)等4組種對之間的生態位重疊指數達到了0.85以上,表明這些種類在同一空間競爭共同資源的現象極其激烈。2020年微駝隆哲水蚤和針刺擬哲水蚤(0)、刷狀螢蝦和針刺擬哲水蚤(0)生態位重疊指數為0,表明這些種類在同一生境中既相互重疊又相互獨立[39]。除2020年肥胖箭蟲和中華哲水蚤、肥胖箭蟲和橈足類無節幼體以及刷狀螢蝦和微駝隆哲水蚤之間生態位重疊顯著外,兩年中其余種對間生態位重疊顯著的均為橈足類生物。另外,2019年針刺擬哲水蚤的生態位寬度值為1.47,到2020年時已下降為0.45,其原因亦可能是其與刺尾紡錘水蚤(0.98)和擬長腹劍水蚤(0.97)的種間競爭有密切的關系。從生態類群看,前兩者均為暖水低鹽種,擬長腹劍水蚤雖為廣溫廣鹽種[40],但有研究證明擬長腹劍水蚤對低鹽度表現出一定的偏好,且對溫度和鹽度都表現出較高的耐受性[41],因此三者之間的種間競爭較為激烈,從而導致針刺擬哲水蚤的生態位寬度值下降。
4.4 浮游動物優勢種生態位與環境因子的關系分析
浮游動物體積較小,游泳能力微弱,其移動通常靠水的流動,浮游動物的生態位分布主要受環境因子的影響[4,6]。舟山海域位于長江及錢塘江兩大河口的交匯處,大量營養物質隨河水流入,同時受臺灣暖流和沿岸寒流的影響,使得該海域環境因子復雜多樣[42],因此浮游動物種類及分布也呈多樣化。冗余分析能很好地考慮環境因子對物種的影響,反映浮游動物對不同生境的適應性特征[34]。根據RDA結果分析, 2019年浮游動物優勢種生態位分化主要受SiO3-Si濃度、COD、Chla濃度和溫度的影響,2020年浮游動物優勢種生態位分化主要受SiO3-Si濃度和DO濃度的影響。
本研究發現,SiO3-Si濃度是影響舟山海域2019-2020年浮游動物優勢種變化的主要因素之一,2019年有多種浮游動物優勢種與SiO3-Si濃度成正相關,2020年多數與SiO3-Si濃度成負相關,存在一定的差異,姜會超等[43]和卞少偉等[44]均發現浮游動物豐度與硅酸鹽濃度呈正相關,但龐碧劍等[26]發現浮游動物豐度與硅酸鹽濃度呈負相關,朱旭宇等[45]發現浮游生物群落參數與硅酸鹽濃度等具有較強相關性,但因物種的不同而存在差異。氮∶磷∶硅比值為16∶1∶16是浮游植物生長過程中吸收營養鹽的最佳比值[45],且營養鹽濃度及結構不同,對浮游動物群落結構調控作用不同。本研究中,2019年氮∶磷∶硅比值為25∶1∶30,2020年氮∶磷∶硅比值為78∶1∶230,可見本海域存在嚴重的營養鹽比例失調現象。而SiO3-Si是許多海洋浮游植物尤其是硅藻必需的營養鹽之一,舟山海域浮游動物優勢種多為橈足類,喜食硅藻[46],一方面硅酸鹽等營養鹽通過影響硅藻等浮游植物的分布間接影響浮游動物分布[1],與浮游動物之間并非簡單的線性關系,另一方面舟山海域捕撈、開發和航行等繁密人類活動也可能對浮游動物產生影響,應在今后的研究中增加對浮游植物的測定及研究,進一步探討浮游動物群落變化機理。
本研究發現,舟山海域2019年多數浮游動物優勢種與COD、Chla濃度和溫度呈正相關,該結果與俞存根等[47]、駱鑫等[13]、邊佳胤等[14]和葉文建等[2]在舟山海域夏季所得結果相似。COD可反映出水體中有機物污染的程度和水體溶解氧的交換平衡,是評測有機物相對含量的重要指標,Chla濃度可評估海域初級生產力和浮游植物生物量,與浮游植物豐度相關性高[48],COD和Chla濃度均可通過影響浮游植物的分布間接影響浮游動物分布[49];海水溫度一方面可影響浮游動物的新陳代謝強度直接影響浮游動物分布[50-51],另一方面可影響浮游植物的生長分布間接影響浮游動物分布[52],2019年舟山海域存在厄爾尼諾現象,使得海水溫度較高,優勢種多為暖水種,如針刺擬哲水蚤等;2020年舟山海域多數浮游動物優勢種與DO濃度成負相關,該結果與卞少偉等[44]在遼東灣夏季及葉文建等[2]在舟山海域夏季所得結果相似,但也有一些浮游動物優勢種與DO濃度成正相關,邊佳胤等[14]發現舟山海域夏季浮游動物豐度與DO濃度成正相關。水中DO濃度是浮游動物生活和分布的重要因素,其濃度高低可以直觀表征生物的環境狀況[28]。葉文建等[2]監測時間、范圍為2014年7月舟山島附近,邊佳胤等[14]監測時間、范圍為2016年8月舟山及鄰近海域,時空分布不同,可能會導致結果存在一定的差異,可見浮游動物與溶解氧之間的關系比較復雜,存在一定的相關性,但這種相關性可能會因時空分布及人為因素等存在差異。
4.5 舟山海域浮游動物歷史變化及人類活動的影響
參考近年來舟山海域夏季浮游動物研究報道(表6),2011年與2014年相比,2019年與2020年相比,舟山海域浮游動物種類數和豐度均下降。2016年的研究范圍主要為舟山各島近岸海域,分布在近海的站位相對較少,因此導致種類數、豐度及生物量均偏高。2019-2020年的研究中,采集浮游動物使用的是淺水Ⅱ型浮游動物網具,網口面積小,因此導致物種數、豐度偏高,但這種情況下下降趨勢仍較為明顯。
舟山作為重要的港口城市和中國最大的漁場,人口繁密,人為干擾會對浮游動物造成巨大的影響。浮游動物是海洋水質監測的重要指標,且可以作為漁業資源的重要餌料,對舟山漁場漁業資源的維持具有重要意義。葉文建等[2]發現人類活動對舟山西部海域的生物群落干擾顯著;陸延等[53]發現舟山電廠排水口海域浮游動物種類數、密度、生物量高于對照點海域;駱鑫等[13]發現石油污染加劇會導致浮游動物豐度和生物量偏低。2020年舟山市生態環境狀況公報顯示:一、二、三、四類海水比例均上升,劣四類海水比例下降,保護環境政策初具成效,但是富營養化仍較為嚴重,下一步可針對舟山海域浮游生物和生態環境進行長期、多次檢測和評估,以期為恢復舟山漁場的海洋生態提供一些參考資料。
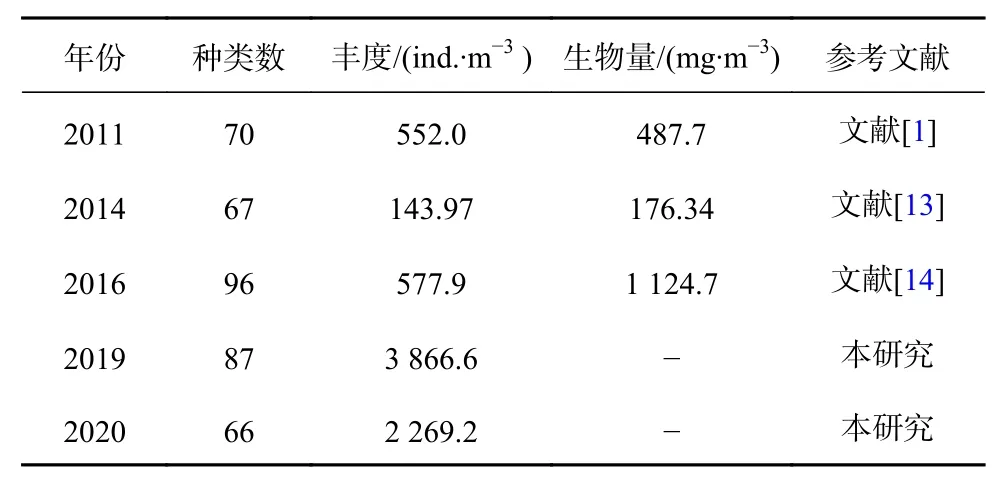
表6 不同年份夏季舟山海域浮游動物統計Table 6 Statistics of zooplankton in summer of different years in the Zhoushan waters
5 結論
(1)2019年和2020年夏季調查共采獲浮游動物8大類130種,橈足類是浮游動物主要類群。2019年浮游動物優勢種6種,2020年浮游動物優勢種10種,兩年共有優勢種4種,兩年浮游動物優勢種年際差異明顯,更替率為66.67%。
(2)2019年夏季浮游動物優勢種的生態位寬度范圍為1.47~2.27,生態位重疊指數范圍為0.11~0.79,廣生態位種占浮游動物優勢種的66.67%;2020年夏季浮游動物優勢種的生態位寬度范圍為0.45~2.34,生態位重疊指數范圍為0~0.98,廣生態位種占浮游動物優勢種的60.00%;兩年夏季廣生態位種物種是舟山海域浮游動物優勢種的主要組成成分,但未呈現出廣生態位種之間生態位重疊程度均較高的現象。
(3)根據RDA結果,2019年浮游動物優勢種生態位分化主要受SiO3-Si濃度、COD、溫度和Chla濃度的影響。2020年主要受SiO3-Si濃度和DO濃度的影響。
猜你喜歡
保健醫苑(2021年7期)2021-08-13 08:48:02
中國民間療法(2021年1期)2021-04-20 02:30:34
趣味(語文)(2020年3期)2020-07-27 01:42:46
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
領導文萃(2015年4期)2015-02-28 09:19:05
中國火炬(2014年11期)2014-07-25 10:31:58
中醫研究(2014年8期)2014-03-11 20:29:23
中國中醫藥現代遠程教育(2014年17期)2014-03-01 04:29:28
