miR-146a通過調節IRAK1影響急性胰腺炎炎癥自噬機制的研究
2022-09-21 06:38:26鄭傳明徐志鵬杜召輝竇賀賀王振杰
安徽醫科大學學報 2022年8期
關鍵詞:研究
鄭傳明,紀 忠,徐志鵬,杜召輝,竇賀賀,姜 海,王振杰
作者單位:蚌埠醫學院第一附屬醫院急診外科,蚌埠 233000
急性胰腺炎(acute pancreatitis,AP)是一種常見的外科急腹癥,其發病率和病死率都很高,目前對其發病機制知之甚少,也沒有特異性或有效的治療方法[1-2]。多項研究[3-4]表明,AP的發病機制與自噬失調和持續的炎癥密切相關。中文名小RNA(microRNA,miRNA)是長度為18~25 nt的單鏈非編碼小分子RNA,近年來其在AP中的作用受到了廣受關注。相關研究[5-7]表明,miR-146a在炎癥和先天性免疫應答中起著重要的調節作用,且有個別報道[8]顯示miR-146a有作為AP預后標志物的潛力,但其在AP中的具體作用機制尚未見研究。該研究利用牛磺石膽酸3磷酸鹽(taurolithocholic acid 3-sulphate,TLCs)處理AR42J細胞建立AP模型,探究miR-146a在AP中的作用及機制,為AP的機制研究提供新思路,更為AP精準診療提供潛在靶點。
1 材料與方法
1.1 材料TLCs(美國Sigma-Aldrich公司),miR-146a mimic及陰性對照(miR-ctrl)、si-IRAK1及陰性對照(si-ctrl)(廣州銳博生物技術有限公司),引物(蘇州金唯智生物科技有限公司),IL-1受體相關激酶1(IRAK1)、actin抗體(美國Abcam公司)、LC3、p62抗體、兔二抗(美國CST公司),白細胞介素6(interleukin-6,IL-6)和腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)試劑盒(杭州聯科生物技術有限公司),Ham's F-12K培養基、血清(美國Gibco公司)。逆轉錄試劑盒及qPCR試劑盒(日本Takara公司)。
1.2 方法
1.2.1細胞培養及AP模型構建 細胞復蘇于細胞培養瓶中,用含有20%的胎牛血清的Ham's F-12K培養基培養,細胞培養箱中設置的條件為37℃、5%CO2。細胞傳3代后,在細胞處于對數生長期時進行細胞功能學實驗。將細胞種于6孔板內,分別加入100、200 μmol/L TLCs,40 min后收集細胞進行后續實驗。
1.2.2RNA抽提及逆轉錄 將處理好的細胞均勻地鋪在培養板內,培養適當時間后棄去培養基,用PBS洗1次,每孔加入1 ml TRIzol,充分收集細胞,加入200 μl氯仿,充分混勻后室溫靜置5 min;4℃、12 500 r/min離心15 min;吸取上清液,轉移至新的1.5 ml離心管中,加入等體積異丙醇,冰上靜置10 min,4℃、12 500 r/min離心10 min;棄上清液,加入1 ml預冷好的75%乙醇溶液,重懸沉淀,4 ℃、12 500 r/min離心5 min,棄上清液,待沉淀變成無色透明狀,加入適量的RNase-free水溶解RNA,并檢測各組濃度。RNA逆轉錄及熒光實時定量PCR的步驟參照試劑盒說明書。引物序列見表1。

表1 引物序列
1.2.3蛋白提取 將處理好的細胞均勻地鋪在培養板內,培養適當時間后棄去培養基,用PBS洗1次,每孔加入300 μl細胞裂解液,冰上靜置15 min,4 ℃、13 000 r/min離心15 min;吸取上清液,轉移至新的1.5 ml離心管中,留取一部分用來測蛋白濃度,剩余液體按照比例加入5×loading,煮10 min,放入-20 ℃儲存備用。
1.2.4細胞轉染 細胞密度長至培養板孔的80%~90%,進行轉染操作。首先用50 μl opti-medium稀釋1 μl Lipofectamine 3000,再用50 μl opti-medium稀釋500 ng質粒,加入1 μl T3000;充分混勻后,將質粒和T3000混合物滴加到Lipofectamine 3000稀釋液中,室溫靜置15~20 min,滴加到24孔板培養基中。miRNA的轉染方法參照質粒轉染方法。
1.2.5熒光素酶報告基因實驗 構建pMIR-IRAK1 wt 3’UTR及結合位點突變pMIR-IRAK1 mut1 和mut2 3’UTR質粒。將上述3種質粒分別轉染到293T細胞中,再次轉染miR-146a mimics或陰性對照,檢測熒光素酶活性。
2 結果
2.1 TLCs誘導AR42J細胞炎癥采用不同濃度(0、100、200 μmol/L)TLCs誘導AR42J細胞,使細胞發生炎癥反應,檢測促炎因子TNF-α和IL-6的mRNA及蛋白水平的變化。結果表明,在用不同濃度TLCs處理的AR42J細胞中,TNF-α和IL-6的mRNA表達水平升高(圖1A)。ELISA分析顯示,在不同濃度TLCs刺激的AR42J細胞培養上清液中,TNF-α和IL-6的蛋白水平也顯著增加(圖1B)。
2.2 TLCs抑制miR-146a促進IRAK1表達并誘導AR42J細胞自噬用不同濃度的TLCs處理AR42J細胞,通過qPCR和Western blot測定miR-146a和IRAK1的表達。結果顯示:與對照組相比,用100 μmol/L TLCs處理AR42J細胞,miR-146a表達降低率為(40±1.22)%,用200 μmol/L TLCs處理AR42J細胞,miR-146a表達降低約(60±0.98)%(圖2A);但是,用100、200 μmol/L TLCs處理AR42J細胞,IRAK1 mRNA水平是對照組的(2.5±0.12)倍和(4±0.08)倍,蛋白水平也同樣有上升(圖2B、C)。表明miR-146a和IRAK1可能在AP的發展中起重要作用。此外,在用TLCs處理的AR42J細胞中,自噬相關標志物LC3-Ⅱ/Ⅰ比值和p62的表達水平也有所變化,即LC3-Ⅱ/Ⅰ比值增加、p62減少(圖2D),提示TLCs可以抑制miR-146a、促進IRAK1表達并誘導AR42J細胞發生自噬。
2.3 過表達miR-146a可以抑制TLCs誘導的炎癥和自噬為了研究miR-146a在AP中的功能作用,用Lipofectamine 3000分別向AR42J細胞轉染miR-146a mimic和miR-control,然后用TLCs(200 μmol/L)處理。qPCR和ELISA分析結果表明,在未加TLCs處理的條件下,過表達miR-146a未引起顯著的IL-6和TNF-α變化。在TLCs處理條件下,過表達miR-146a顯著降低IL-6和TNF-α的mRNA和蛋白水平(圖3A、B)。自噬試驗結果顯示,在轉染miR-control的處理條件下,TLCs引起LC3-Ⅱ/Ⅰ比值升高和p62表達下降,該結果與前述結果一致;與轉染miR-control且添加TLCs處理相比,過表達miR-146a,即轉染miR-146a mimic+TLCs處理,顯著降低LC3-Ⅱ/Ⅰ比值且上調p62表達(圖3C)。以上結果說明,上調miR-146a可以抑制由TLCs誘導的炎癥和自噬應答。
2.4 敲減IRAK1可以抑制TLCs誘導的炎癥和自噬為了進一步研究IRAK1在AP中的功能,在AR42J細胞中轉染si-IRAK1或siRNA-control,然后用TLCs處理(200 μmol/L)。Western blot結果表明,與siRNA-control組相比,si-IRAK1組中IRAK1的蛋白表達下降(圖4A)。敲減IRAK1顯著降低了TNF-α和IL-6的mRNA和蛋白水平(圖4B、C)。此外,敲減IRAK1減弱了TLCs誘導的LC3-Ⅱ/Ⅰ比值升高和p62表達降低(圖4D)。該部分結果顯示,敲減IRAK1與上調miR-146a表達對TLCs誘導的AP細胞模型中具有相似的作用。
2.5 miR-146a在AR42J細胞中靶向IRAK1為了進一步探索AP中miR-146a與IRAK1之間的關系,在AR42J細胞中過表達miR-146a,再用TLCs處理細胞,Western blot和qPCR檢測IRAK1表達。結果顯示:miR-146a可以明顯抑制IRAK1表達,TLCs可以促進IRAK1表達,同時加入miR-146a和TLCs,與只加TLCs組相比,IRAK1表達量也顯著降低(圖5A),這說明miR-146a可以抑制IRAK1的表達。為了更深入探究miR-146a與IRAK1靶向關系,通過Targetscan在線網站預測了miR-146a與IRAK1 3′UTR 的潛在結合位,并構建野生型和結合位點突變型質粒,通過熒光素酶實驗進一步驗證miR-146a與IRAK1靶向關系。結果表明,miR-146a的過表達強烈降低了IRAK1-wt的螢光素酶活性,而轉染miR-146a后,IRAK1-mut的螢光素酶活性未受到明顯影響(圖5B、C)。綜上所述,這些結果表明miR-146a可以直接靶向IRAK1。
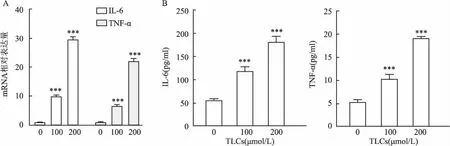
圖1 TLCs誘導AR42J細胞炎癥

圖2 TLCs抑制miR-146a促進IRAK1表達并誘導AR42J細胞自噬
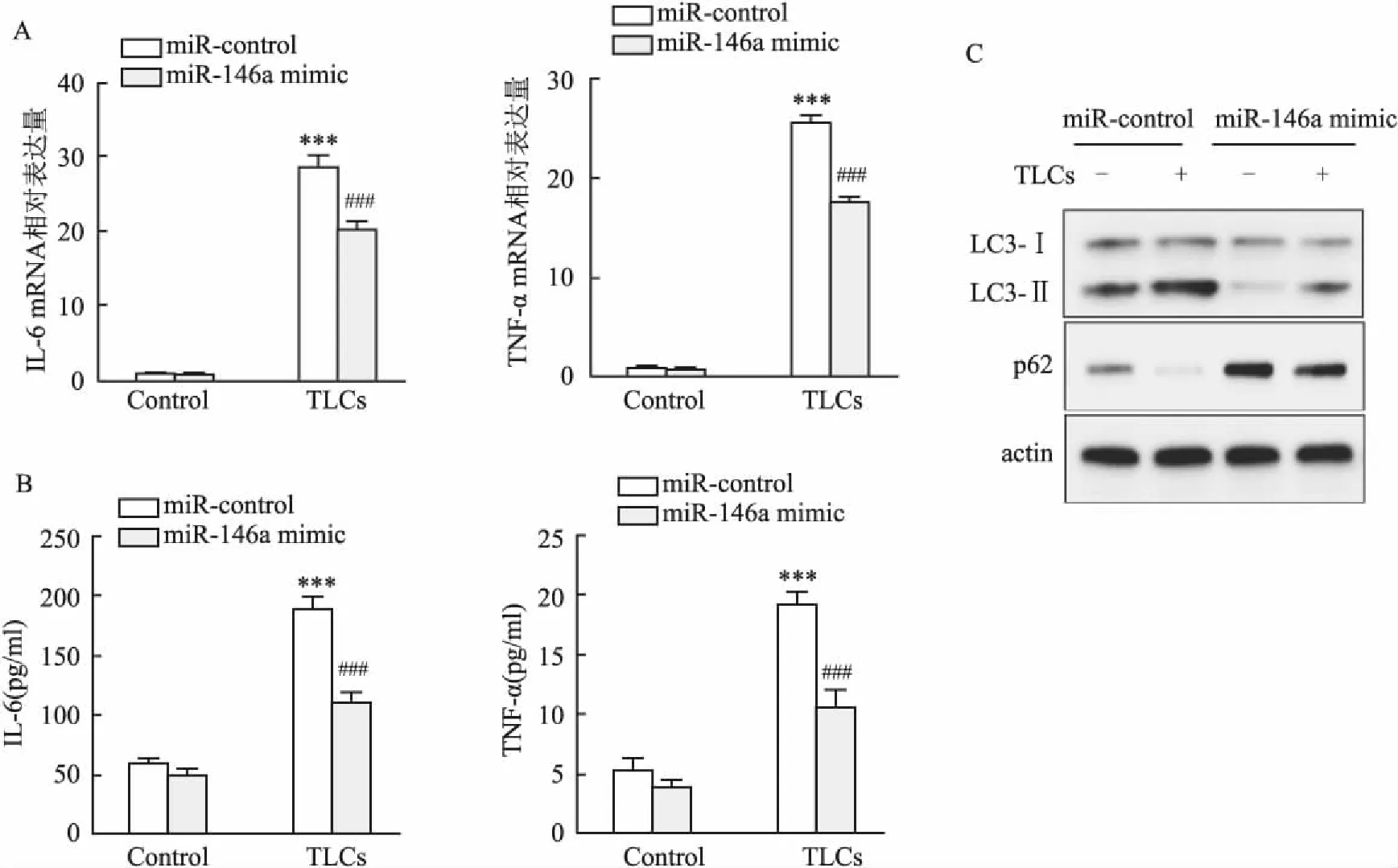
圖3 過表達miR-146a抑制TLCs誘導的炎癥和自噬
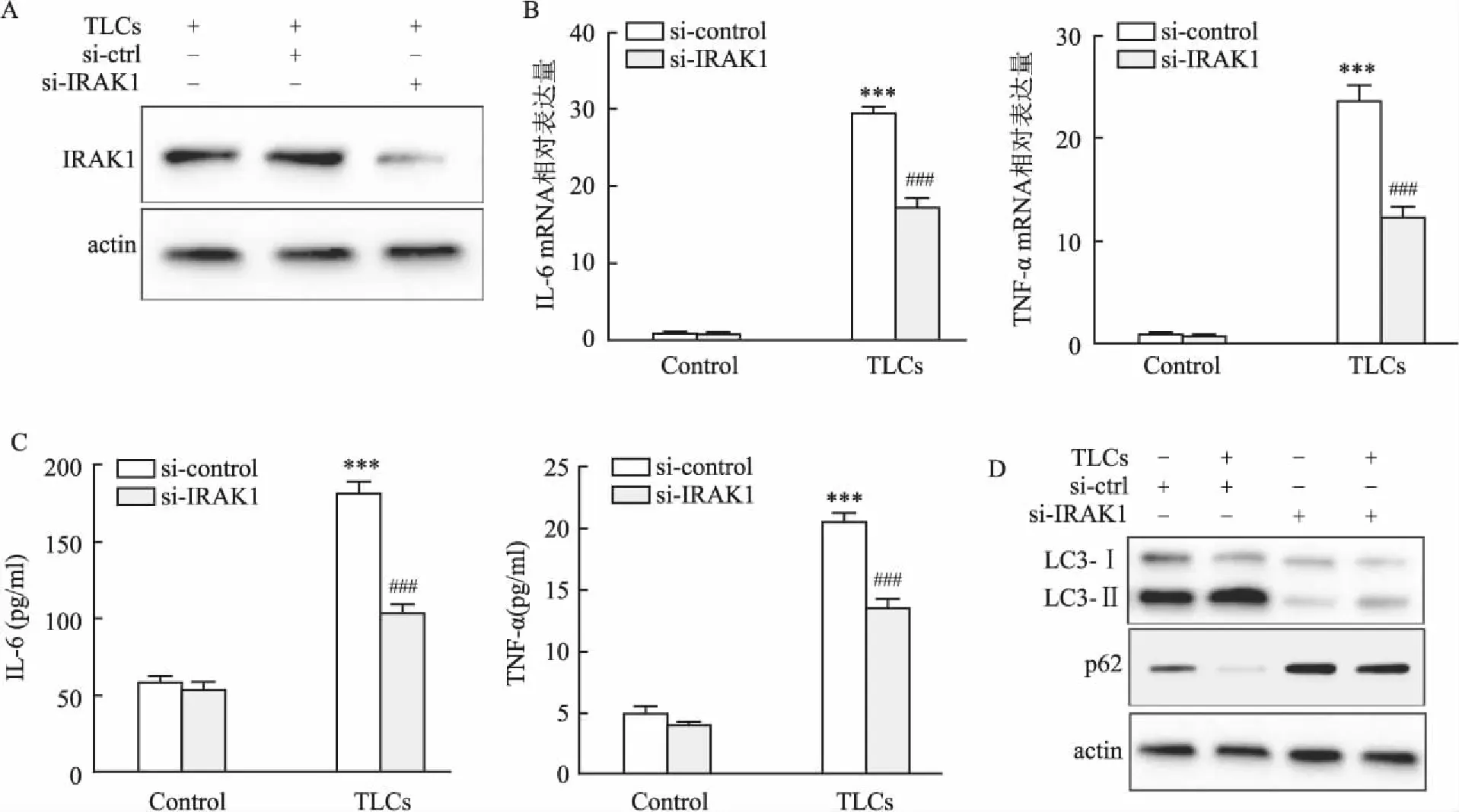
圖4 敲減IRAK1可以抑制TLCs誘導的炎癥和自噬
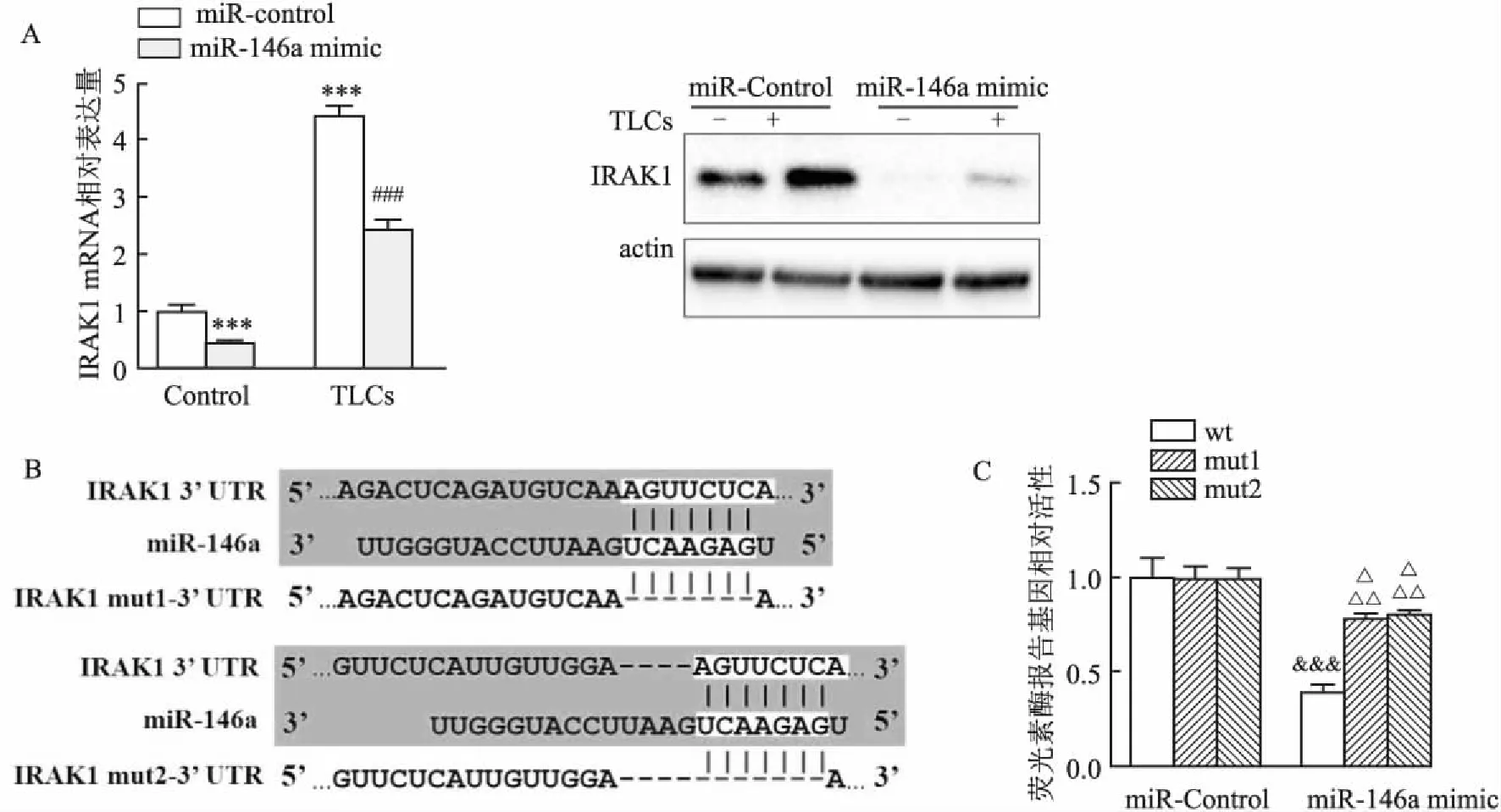
圖5 miR-146a在AR42J細胞中靶向IRAK1
3 討論
AP具有不同的臨床病程。大多數患者表現為輕度AP,具有自限性,通常在1周內消退。大約20%的患者發展為中度或重度AP,伴有胰腺或胰周組織壞死,或是器官衰竭,亦或兩者兼有,病死率高達 20%~40%[1-2]。更為重要的是,目前,臨床尚無可以改變AP疾病進程的藥物。因此,深入研究AP的發病機制,可能為AP的機制研究提供新思路,更可能為其治療提供新的靶點。
有研究[3]顯示,AP會導致腺泡細胞的細胞器(內質網、線粒體和溶酶體-自噬系統)受損,并使細胞器功能紊亂,進而導致胰蛋白酶原和炎癥通路的激活。自噬是細胞內主要的分解代謝過程,細胞通過該過程消除受損、有缺陷或不需要的細胞器、蛋白質和脂質等,并產生能量滿足細胞代謝需要。相關研究[9]顯示,在轉基因GFP-LC3小鼠中,GFP-LC3的表達對肝、肺和脾的自噬影響很小或沒有影響,但可以顯著增加胰腺腺泡細胞中的自噬體形成。另一項研究[10]顯示,抑制AP小鼠的自噬可以改善小鼠的預后。本研究也表明,在AP的細胞模型中,隨著TLCs濃度的逐漸增加,LC3-Ⅱ/Ⅰ比值逐漸升高、p62逐漸降低,同時,在炎癥和先天性免疫應答中起著重要負性調節作用的miR-146a表達量也逐漸降低。上調miR-146a可以顯著抑制TLCs誘導的AR42J細胞自噬。說明miR-146a可調控TLCs誘導的AR42J細胞自噬,且miR-146a的表達與AP的進展密切相關。
眾所周知,巨噬細胞在炎癥反應中起重要作用。巨噬細胞產生各種細胞因子和炎性介質,如IL-1β、IL-6、IL-8和TNF-α,它們可損傷附近組織和遠處器官。抑制胰腺炎相關巨噬細胞的活化不僅減輕了胰腺本身的病理損傷,而且在一定程度上減少了全身炎癥反應綜合征和多器官功能衰竭的發生[11]。研究[12]顯示通過腹腔灌洗清除巨噬細胞后AP小鼠的TNF-α、IL-6和IL-10血清水平降低。本研究顯示,TLCs可以促進炎癥因子IL-6和TNF-α分泌,且隨著TLCs的濃度逐漸增加,IL-6和TNF-α的表達量也逐漸增加。同時,TLCs可抑制AR42J細胞內miR-146a的表達、促進IRAK1表達。IRAK1是介導的NF-κB活化途徑的重要元件,NF-κB對于炎性反應和介導炎性因子(例如TNF-α、IL-1β和IL-6)的釋放是重要的,并可加劇炎性損傷[13]。NF-κB在AP相關巨噬細胞激活過程中起重要作用[14]。本研究在AR42J細胞中過表達miR-146a,顯著抑制TLCs誘導的炎癥及IRAK1的表達。結合熒光素酶報告基因實驗,進一步證明了miR-146a可以靶向調控IRAK1的表達。這些結果提示miR-146a和IRAK1極可能是AP治療的潛在靶點。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
