低溫脅迫對4 個三角梅品種葉綠素熒光參數的影響
2022-09-16 06:05:14伍成厚余銘杰傅小霞鄭育芬吳佳慶陳秋香唐湘楠
湖南農業科學 2022年8期
伍成厚,余銘杰,溫 志,傅小霞,鄭育芬,吳佳慶,陳秋香,唐湘楠
(廣州市林業和園林科學研究院,廣東 廣州 510405)
三角梅(Bougainvilleaspp.)為紫茉莉科三角梅屬植物,在廣州地區俗稱簕杜鵑,是海南省的省花、深圳和廈門等城市的市花,也是我國南方廣為應用的一種木本花卉。因其適應性強、花期持久,在廣州、深圳等地是城市橋梁綠化的首選植物。三角梅原產南美巴西,喜溫暖濕潤的氣候,怕冷不耐寒,大多數品種對4℃以下的低溫敏感[1],半致死溫度為-3℃[2],但也有半致死溫度低至-4.91℃~-9.22℃的品種[3]。冬季低溫是北方地區引種三角梅的主要障礙[4]。在廣州地區,冬季低溫也會對天橋綠化的三角梅枝葉造成不同程度的損傷,影響橋梁綠化的景觀效果[5]。
有研究表明,低溫脅迫會導致三角梅葉片電解質外滲率升高,不同品種間的耐寒性差異與葉片丙二醛、游離脯氨酸、可溶性蛋白質和葉綠素含量以及保護酶活性等生理指標相關[3,6],通過嫁接[7]、施用防凍抗寒劑可以提高三角梅的抗寒性[7-10]。丁印龍等[11]研究了干旱脅迫對三角梅葉綠素熒光參數的影響,但沒有低溫脅迫下三角梅葉綠素熒光特性的研究報道。采用葉綠素熒光技術可快速、靈敏、無損傷地探測植物對低溫脅迫的響應,該技術目前已應用于秋茄(Kandelia obovata)[12]、 諾 麗(Morinda citrifolia)[13]等 熱 帶、亞熱帶植物的低溫脅迫鑒定。低溫會導致光系統Ⅱ(PS Ⅱ)最大光化學效率(Fv/Fm值)顯著下降,且抗寒性弱的品種降幅顯著大于抗性強的品種[12-15],因此葉綠素熒光參數可作為篩選耐寒品種的重要指標。為了探討三角梅的耐寒性機理,筆者以4 個三角梅品種的葉片為材料,研究了低溫脅迫下三角梅葉綠素熒光參數的變化規律,以期為三角梅耐寒品種的篩選提供參考。
1 材料與方法
1.1 供試材料
供試三角梅有小葉紫、水紅、玫紅和櫻花共4 個品種,均為三年生的盆栽苗,采用露天常規水肥管理,植株生長健壯。
1.2 試驗方法
試驗于2021 年9 月進行。分別選取4 個三角梅品種樹冠上層外圍枝條上倒數第5~8 片葉,在晴天9點前采樣,置于自封袋中迅速帶回實驗室,在不對葉片造成機械損傷的前提下,擠出袋內空氣。將自封袋在黑暗條件下放入3℃冰水混合液中水浴進行低溫脅迫,低溫脅迫時間設1、2、3 和4 d 共4 個處理,以不進行低溫脅迫為對照(CK)。然后將不同處理材料(葉片)從自封袋中取出,用濾紙或紗布擦干水珠后利用德國WALZ-便攜式調制葉綠素熒光儀PAM2500測定離體葉片的F0、Fm、Fv/Fm和Y(NO)等葉綠素熒光動力學參數值。每個處理設3 個重復,每個重復測定3 次取平均值。
1.3 數據處理
用Excel 2007 軟件對數據進行計算和制圖,用SPSS 19.0 軟件進行方差分析。
2 結果與分析
2.1 低溫脅迫對三角梅葉綠素熒光參數F0、Fm 值的影響
F0為固定熒光或初始熒光,也稱基礎熒光、0 水平熒光。從表1 可知,小葉紫和櫻花2 個三角梅品種不同低溫脅迫時間處理間的葉綠素熒光參數F0值差異不顯著;而玫紅和水紅2 個三角梅品種不同低溫脅迫時間處理間的葉綠素熒光參數F0值存在極顯著性差異,在低溫脅迫處理1 d 時玫紅品種的F0值略有上升而水紅品種的F0值急劇下降,但在低溫脅迫處理1~4 d 時玫紅和水紅2 個三角梅品種的F0值均呈先升后降的變化趨勢。
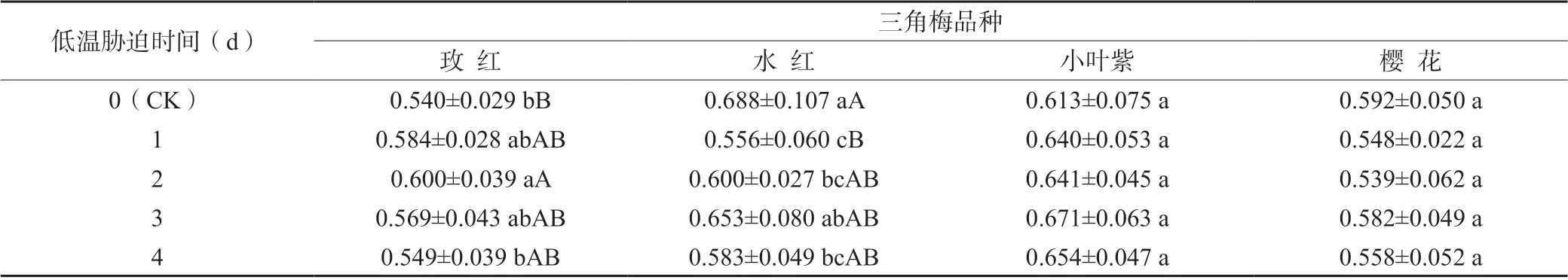
表1 不同低溫脅迫時間處理對三角梅品種葉綠素熒光參數F0 值的影響
Fm為最大熒光,是PS Ⅱ反應中心處于完全關閉時的熒光產量。低溫脅迫對4 個三角梅品種的葉綠素熒光參數Fm值均產生極顯著影響(表2),隨低溫脅迫時間的延長均呈現下降的趨勢,玫紅、水紅和櫻花3 個三角梅品種均在低溫脅迫1 d 后Fm值極顯著下降,而小葉紫品種則在低溫脅迫3 d 后才極顯著下降;當低溫脅迫4 d 時,玫紅、水紅、小葉紫和櫻花4 個三角梅品種的Fm值分別比CK 下降40.3%、25.6%、17.4%和30.8%。
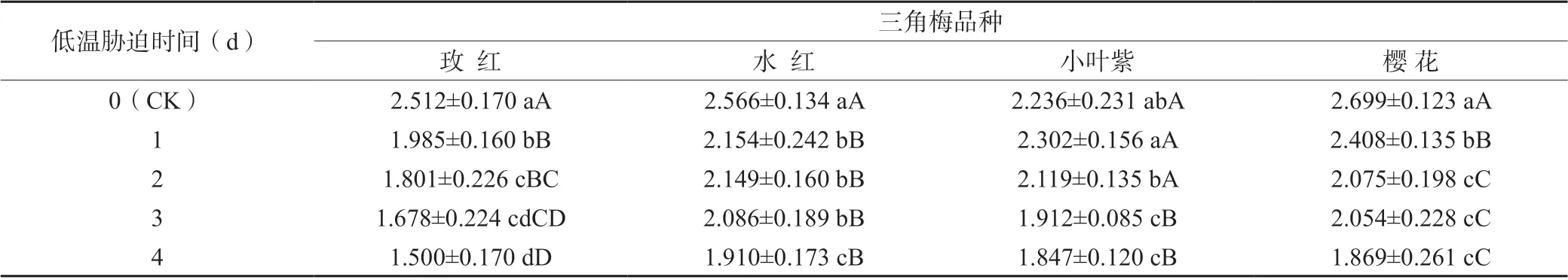
表2 不同低溫脅迫時間處理對三角梅品種葉綠素熒光參數Fm 值的影響
2.2 低溫脅迫對三角梅葉綠素熒光參數Fv /Fm值的影響
葉綠素熒光參數Fv/Fm反映PS Ⅱ反應中心內稟光能轉換效率或稱最大PS Ⅱ的光能轉換效率。低溫脅迫后4 個三角梅品種的Fv/Fm值均呈極顯著下降的趨勢(玫紅的F=33.500,P=0.000<0.01;水紅的F=6.426,P=0.000<0.01;小葉紫的F=17.349,P=0.000<0.01;櫻花的F=31.690,P=0.000<0.01)。從圖1 可知,玫紅品種在低溫脅迫1 d 時Fv/Fm有極顯著下降,然后下降趨緩,在低溫脅迫3 d 后再次極顯著下降;而水紅、小葉紫和櫻花3 個品種在低溫脅迫3 d 后才開始明顯下降,其中小葉紫在低溫脅迫3 d 后下降速度明顯減緩,而水紅在低溫脅迫4 d 時有上升的趨勢;當低溫脅迫處理4 d 時,玫紅、小葉紫、櫻花和水紅4 個三角梅品種的Fv/Fm值比CK 分別下降19.5%、10.8%、10.2%和5.2%。
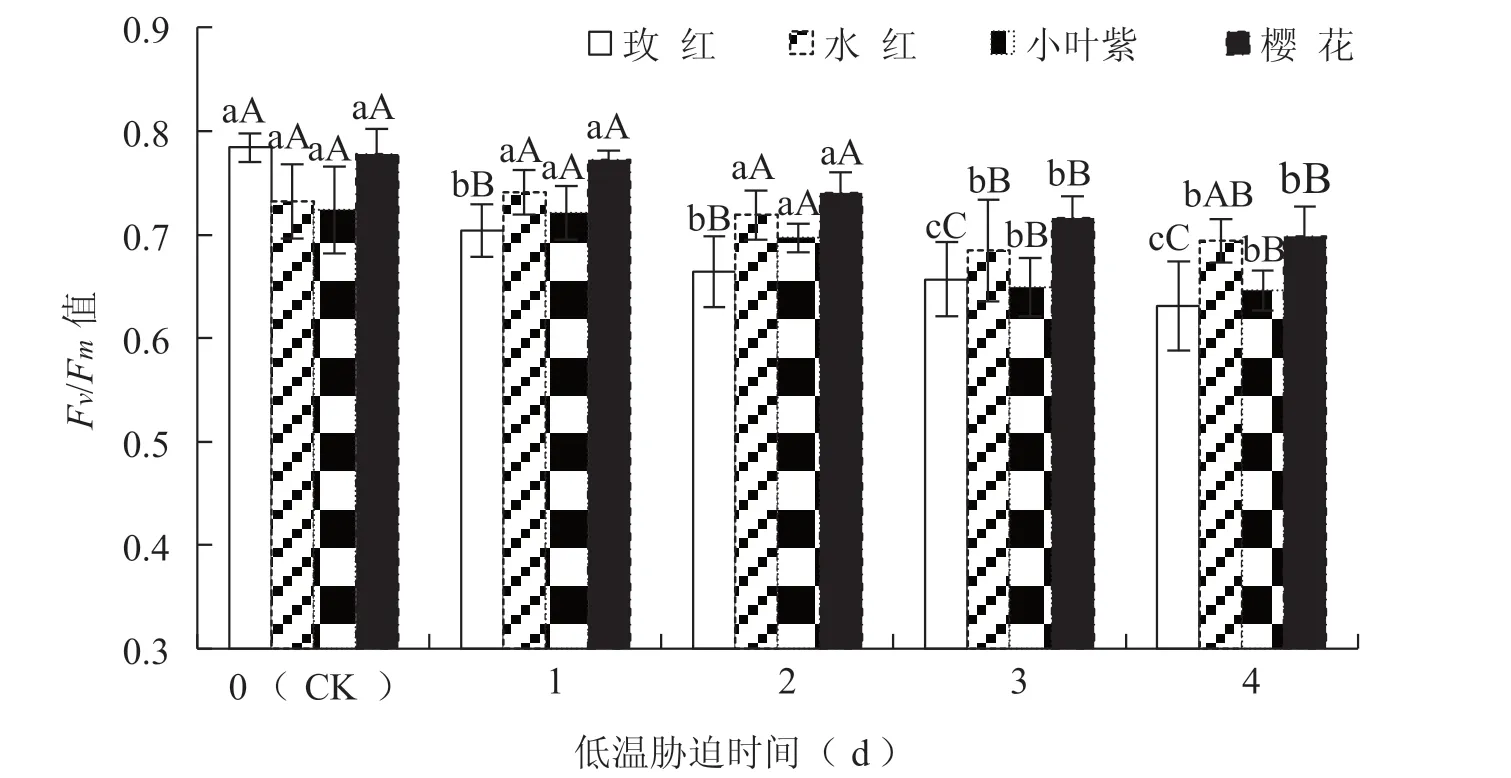
圖1 低溫脅迫對三角梅葉綠素熒光參數Fv /Fm 值的影響
2.3 低溫脅迫對三角梅葉綠素熒光參數Y(NO)的影響
Y(NO)為PS Ⅱ非調節性能量耗散。不同低溫脅迫時間處理對4 個三角梅品種葉綠素熒光參數Y(NO)值的影響均達到極顯著水平(玫紅的F=8.291,P=0.000<0.01;水 紅 的F=5.031,P=0.000<0.01;小葉紫的F=16.817,P=0.000<0.01;櫻花的F=23.010,P=0.000<0.01)。低溫脅迫處理使三角梅葉綠素熒光參數Y(NO)逐步升高,櫻花在低溫脅迫2 d 時極顯著升高,水紅和小葉紫均在低溫脅迫3 d 時極顯著升高,玫紅在低溫脅迫2 d 時顯著升高,4 d 時極顯著升高;當低溫脅迫處理4 d 時,櫻花、玫紅、小葉紫和水紅的Y(NO)值比CK 分別升高35.0%、24.5%、22.0%和17.5%(圖2)。
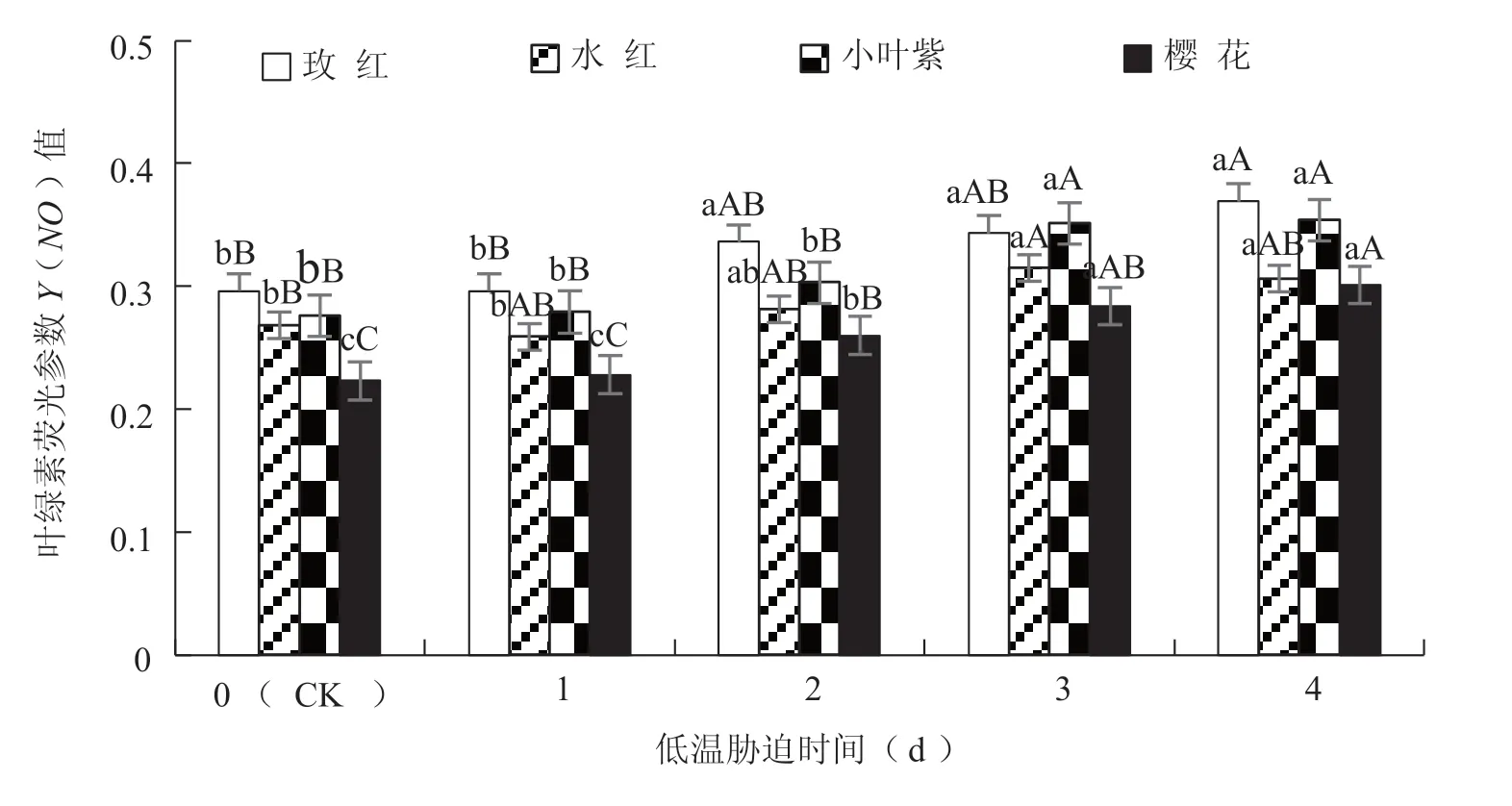
圖2 低溫脅迫對三角梅葉綠素熒光參數Y(NO)值的影響
3 討 論
初始熒光F0由PS Ⅱ捕光葉綠素蛋白復合物發出,代表不參與PS Ⅱ光化學反應的光能輻射部分,是PS Ⅱ光化學反應中心處于完全開放時的熒光產量,它與葉綠素濃度有關。一般情況下其數值保持穩定,當光合結構受到損壞時則會升高。F0的增加可能是植物葉片PS Ⅱ反應中心出現可逆的失活或出現不易逆轉的破壞,也可能是植物葉片類囊體膜受到損傷,而且F0增加量越多類囊體膜受損程度就越嚴重[16]。蝴蝶蘭(Phalaenopsis aphrodite)在6℃低溫脅迫下6 個品種的F0值均有不同程度的升高[14]。茄子(Solanum melongena)在10℃和15℃低溫脅迫下其F0值均顯著高于對照[15],但秋茄在5 和12℃低溫脅迫1~5 d后,其F0的變化率與對照的差異不顯著[12]。花椰菜(Brassica oleracea)的F0值在經受寒潮低溫脅迫2 d后顯著升高,但僅在不同熟性的品種間具有顯著差異,在相同熟性的品種間差異不顯著[17]。筆者的試驗結果表明,經3℃低溫脅迫后,小葉紫和櫻花2 個三角梅品種的葉綠素熒光參數F0值沒有顯著性差異,而玫紅和水紅2 個三角梅品種的F0值在低溫脅迫處理1~4 d 時均呈先升后降的變化趨勢,差異達到極顯著水平,可能是由于玫紅和水紅2 個品種的光系統在3℃低溫脅迫下較容易受損,而小葉紫和櫻花2 個品種因耐寒性較強而使光系統得到較好保護。
最大熒光Fm是PS Ⅱ反應中心處于完全關閉時的熒光產量,Fm值下降與PS Ⅱ關閉程度有一定關系[14]。在低溫脅迫下茄子的Fm值顯著低于對照[15]。隨著低溫脅迫時間的延長,葛藤(Pueraria lobata)、假連翹(Duranta repens)、老鴨嘴(Thunbergia erecta)和臺灣相思(Acacia confuse)幼苗的Fm值也呈逐漸下降趨勢[18]。筆者的試驗結果表明,經低溫脅迫處理后,玫紅、水紅、小葉紫和櫻花4 個三角梅品種的Fm值均呈下降趨勢,分別比CK 下降40.3%、25.6%、17.4%和30.8%,以玫紅下降最多、小葉紫下降最少,說明4 個品種中小葉紫的耐寒性最強、玫紅的耐寒性最弱。
PS Ⅱ最大光化學效率Fv/Fm在正常條件下變化極少,不受物種和生長條件的影響,但該參數在逆境下明顯下降,常被用于植物的抗逆性研究[16]。在低溫脅迫下,秋茄、諾麗、蝴蝶蘭、茄子等植物的Fv/Fm值均呈下降趨勢,且其變化幅度越大則植物的耐低溫能力越弱[12-15],筆者的試驗結果與之相似。當低溫脅迫處理4 d 時,玫紅、小葉紫、櫻花和水紅4 個三角梅品種的Fv/Fm值分別比CK 下降19.5%、10.8%、10.2%和5.2%,以玫紅的降幅最大,這也說明4 個品種中玫紅的耐寒性最弱。
非調節性能量耗散Y(NO)表示的是光鈍化所引起的非調節性電子產量,是光損傷的重要標志。盧廣超等[18]的研究表明,低溫脅迫后4 種幼苗的Y(NO)均有上升,且當低溫脅迫36 h 時Y(NO)均顯著大于對照,其中臺灣相思幼苗的Y(NO)與24 h 相比升幅最少,表明其光損傷的程度最低,抗寒性較好。何勇等[19]的研究認為,經6℃低溫脅迫后辣椒的Y(NO)均明顯升高,其中杭椒6 號的升幅較大,表明其光化學的能量轉換(用于光合電子傳遞)和光保護機制(調節性熱量耗散)不足,由于PS Ⅱ的過剩電子可能會引起光抑制,而杭椒1 號的Y(NO)變化較為平穩,說明其光合電子傳遞及光保護機制運轉正常。筆者的試驗結果表明,櫻花、玫紅、小葉紫和水紅4個三角梅品種的Y(NO)值分別比CK 升高35.0%、24.5%、22.0%和17.5%,以櫻花和玫紅2 個品種的Y(NO)值升高較多、小葉紫和水紅升高較少,表明玫紅和櫻花的耐寒性較小葉紫和水紅差。
4 結 論
經低溫脅迫處理后,4 個三角梅品種的Fm、Fv/Fm值均呈下降趨勢,Y(NO)值則呈上升趨勢,說明低溫脅迫對三角梅葉片的PS Ⅱ反應中心造成一定損傷,從而降低PS Ⅱ原初光能轉化效率,抑制PS Ⅱ反應中心的光化學活性,使得PS Ⅱ反應中心潛在活性受損,進而破壞三角梅葉片光合作用的原初反應過程。以玫紅的Fm和Fv/Fm值下降最多,櫻花的Y(NO)值升高最多,表明小葉紫和水紅的耐寒性較玫紅和櫻花的強。
