揚子鱷FoxO1基因CDS序列的克隆及其在初孵鱷性腺組織的時空表達規律
2022-08-30 02:01:28胡誠誠汪仁平夏同勝何明月周永康張童欣沈新禹聶海濤吳孝兵
水生生物學報 2022年8期
關鍵詞:研究
溫 月 胡誠誠 汪仁平 夏 勇 夏同勝 何明月 周永康 張童欣 張 軍 龐 宇 王 浩 沈新禹 聶海濤, 吳孝兵,
(1. 安徽師范大學生命科學學院, 蕪湖 241000; 2. 安徽省揚子鱷繁殖研究中心, 宣州 242000;3. 安徽師范大學, 安徽省重要生物資源保護與利用重點實驗室, 蕪湖 241000)
揚子鱷(Alligator sinensis)在脊椎動物分類上隸屬于爬行綱鱷目(Crocodylia), 主要分布在長江中下游地區。其作為中國特有的爬行動物, 由于自然棲息地持續喪失、斑塊化及人類的肆意獵殺造成了近代以來野生揚子鱷數量持續下降, 導致現存野生數量非常稀少。1973年聯合國將其列為臨危種和禁運種我國已經把揚子鱷列為國家一類保護動物,嚴禁捕殺[1]。為防止揚子鱷滅絕, 目前采用人工繁殖的方式進行保護。歷經近半個世紀的人工繁育保護, 其種群數量得到一定的提升, 但是由于性成熟周期較長(雌性成年個體約需8—10年、體長1.5 m、體重約36 kg)等原因, 能繁殖成年個體種質資源尤為珍貴。近年來, 揚子鱷的多父性交配體系[2]、精子儲存[3]和卵黃發生等繁殖特性已逐漸被人們認知, 但其在性腺發育早期的配子發生進程則很少有人關注。原始卵泡的形成和發育直接影響到雌性動物生育階段的可用卵泡數量, 是雌性動物繁殖階段中可利用的全部資源[4], 其在重點保護物種的標簽下能繁殖的成年個體種質資源更顯珍貴。但至今未見有關鱷目動物卵巢中卵母細胞巢形成、裂解及原始卵泡形成過程和分子機制系統的報道。如何通過原始卵泡庫資源的有效調控, 追求其終生生殖配子的利用最大化, 始終是揚子鱷人工繁育與物種保護進程所面臨的重要課題之一。
1989年, 科學家利用染色體步移法在果蠅中克隆到一個控制胚胎頭部正常發育的基因Forkhead(FKH), 并證明該基因的編碼蛋白是細胞核中的一種轉錄調節因子[5,6]。隨后, 人們在小鼠(Mus musculus)、線蟲(Caenorhabditis elegans)、非洲爪瞻(Xenopus tropicalis)、酵母(Candida marina)和人(Homo sapiens)等幾乎所有真核生物中也都陸續發現這類具有保守序列的蛋白或轉錄因子, 并將其統一命名為Fox基因[7]。其中FoxO(Forkhead boxclass O)是研究最多的亞族之一, 且已被證實作為在細胞核與細胞質之間穿梭的轉錄因子, 通過調節凋亡、細胞周期、細胞分化、DNA修復、應激抵抗、長壽、葡萄糖代謝及肌肉生長等相關基因的表達, 發揮了廣泛的生物學效應[8]。目前FoxO轉錄因子只有4個家族成員, 分別為FoxO1(FKHR)、FoxO3a(FKHRL1)、FoxO4(AFX)和FoxO6, 盡管所有哺乳動物的FoxO家族都包含這4個成員, 但它們的編碼基因及染色體定位在不同物種中有很大差異[9]。FoxO蛋白是細胞存活的負調節因子, 通過抑制細胞増殖、細胞凋亡或促進細胞周期阻滯等機制調控哺乳動物的生殖細胞發育[10]。哺乳動物卵巢中含有大量處于不同發育階段的卵泡, 卵泡的發育既依賴于卵巢局部生長調控因子(如IGF-1和雌激素),又依賴于垂體分泌的促性腺激素(FSH和LH)。經證明, 在雌性哺乳動物中FoxO蛋白可以通過多條通路對卵泡的生長、成熟和閉鎖進行調控[11]。Richards等[12]最早發現, 大鼠卵巢中FoxO轉錄因子的表達和活性受IGF-1、雌激素及促性腺激素調控,表明這些內分泌因素會影響FoxO功能的發揮。另有研究表明FoxO1-/-小鼠在出生前即死亡, 且值得注意的是, 所有雌性小鼠的卵巢都出現了全體卵泡激活發育的現象, 導致卵母細胞死亡、卵泡池過早耗盡和繼發性不孕[13]。基于FoxO1在生殖系統和神經系統等中的重要性研究, 其序列已在大鼠和家雞等模式動物或經濟家畜動物中被成功克隆。然而, 截至目前, 少有揚子鱷相關研究中關于FoxO基因表達調控的報道。
本研究以初孵揚子鱷性腺組織作為材料, 通過對Forkhead box家族蛋白-1(FoxO1)基因CDS區序列的克隆分析; 結合免疫組織化學對FoxO1蛋白在初孵鱷性腺復合體、胃、腸和肺臟等組織表達譜進行研究; 利用免疫熒光和Western Blot技術對不同出殼后日齡(17d、63d和96d)雌鱷性腺組織中的FoxO1基因的蛋白質豐度進行定位和定量研究。相關結果將為系統解析FoxO1蛋白在揚子鱷卵子發生過程中的生物學功能提供參考, 對于豐富揚子鱷卵子發生調控機制具有重要的生殖生物學理論研究價值。
1 材料與方法
1.1 材料
分別取來自安徽省宣城市揚子鱷繁殖研究中心的17日齡、63日齡和96日齡3只初孵揚子鱷, 迅速斷頭處死, 解剖取一部分幼鱷性腺組織液氮保存–80℃備用; 另取其性腺、胃、大腸和肺等外周組織, 經多聚甲醛固定液處理后, 無水乙醇梯度脫水,透明、石蠟包埋, 用切片機連續切片(厚約6 μm), 于水浴鍋45℃清水展片并固定于潔凈載玻片上, 于60℃展烘箱中烘干2h, 可直接用于后續的研究。上述行為均得到了安徽師范大學動物倫理委員會的批準。
1.2 方法
總RNA的提取與cDNA的合成取?80℃凍存的幼鱷性腺組織按TRIzol抽提方法, 獲得總RNA,并測其濃度(OD值在1.6—1.8)、純度及完整度后,參照TaKaRa公司反轉錄試劑盒操作說明合成cDNA待用。
FoxO1基因cDNA序列的擴增通過NCBI在鱷目中查找高度同源序列(登錄號: KY713605),運用Primer 5.0軟件設計4對特異性引物(表 1), 以cDNA為模板進行PCR(35個循環, 每個循環94℃,30s; 55℃, 30s; 72℃, 30s; 72℃終止延伸10min)。PCR產物經1%瓊脂糖凝膠驗證條帶大小與引物設計條帶相符后, 送通用生物(安徽滁州)公司進行測序并拼接后獲得揚子FoxO1基因的完整CDS區序列。隨后利用DNAMAN軟件將測序得到的序列與在基因文庫中查找到的序列進行同源性比對分析,構建系統進化樹。

表1 引物序列信息Tab. 1 Information of primers sequences
免疫組化染色石蠟切片脫蠟脫水, 置于0.3%過氧化氫: 甲醇混合溶液中孵育30min, 清除內源性過氧化物酶, 組織打孔后置于抗原修復液中微波加熱至沸騰, 滴加封閉液, 孵育兔源FoxO1一抗(Bioss, bs-9439R, 1﹕300), 置于4℃冰箱過夜, 室溫孵育二抗(羊抗兔, 1﹕500)1.5h后, 用DAB試劑盒染色,蘇木素復染, 脫水透明封片, 光鏡下觀察并攝片。空白對照以PBS代替。
免疫熒光染色石蠟切片脫蠟脫水, 置于0.3%過氧化氫: 甲醇混合溶液中孵育30min, 清除內源性過氧化物酶, 組織打孔后置于抗原修復液中微波加熱至沸騰, 滴加封閉液, 孵育兔源FoxO1一抗(Bioss, bs-9439R ,1﹕200), 置于4℃冰箱過夜, 37℃避光孵育二抗(羊抗兔, 1﹕500)1.5h后, 用DAPI避光復染細胞核, 用抗熒光淬滅劑封片液封片、光鏡下觀察并攝片。
Western Blot取?80℃凍存的17日齡、63日齡和96日齡幼鱷性腺復合體組織, 按RIPA裂解液抽提方法, 獲得總蛋白質, 經SDS-PAGE電泳后將FoxO1及GAPDH內參蛋白目的條帶轉膜至PVDF膜上, 封閉后, FoxO1目的條帶孵育兔源FoxO1一抗(Bioss, bs-9439R, 5%脫脂奶粉1﹕1000稀釋),GAPDH內參蛋白條帶孵育鼠源GAPDH一抗(Bioss,D110016-0025, 1﹕2000), 在4℃條件下搖床過夜, 在室溫條件下孵育二抗(5%脫脂奶粉1﹕2000稀釋)2h,用高靈敏ECL發光試劑盒進行顯影觀察并攝片。
2 結果
2.1 揚子鱷FoxO1基因CDS區序列的克隆
經TA克隆測序和序列的拼接獲得長度為1941 bp的核苷酸片段、預測編碼646個氨基酸殘基, 蛋白理化預測分析表明揚子鱷FoxO1蛋白等電點(isoelectric point; pI)和分子量(Mw)分別為4.88和162 kD。
2.2 FoxO1氨基酸序列同源性比對及構建進化樹
通過將揚子鱷FoxO1基因的CDS區序列編碼的氨基酸序列與灣鱷(XP_019412112)、恒河鱷(XP_019359603)、密西西比短吻鱷(XP_014461938)、中華鱉(XP_014424549)、西部錦龜(XP_005285222)、原矛頭蝮(XP_015673988)、多疣壁虎(XP_015283276)、熱帶爪蟾(NP_001107165)、青鳉(NP_001098169)、小家鼠(AAI16898)、褐家鼠(XP_006252158)、智人(AAI26438)、原雞(NP_001074346)和斑胸草雀(ACK57931)的FoxO1蛋白及揚子鱷FoxO4蛋白的氨基酸序列進行同源性比對后, 發現揚子鱷的FoxO1基因的CDS區序列編碼的氨基酸序列與密河鱷等鱷目物種同源性為96.74%—92.21%; 與西部錦龜(85.87%)、中華鱉(85.34%)、原雞(85.65%)和斑胸草雀(85.02%)等物種的氨基酸同源性次之; 與智人(75.58%)、小家鼠(74.96%)、褐家鼠(74.81%)、熱帶爪蟾(72.79%)、壁虎(69.95%)和原矛頭蝮(68.61%)具有中等同源性; 而與青鳉的氨基酸同源性最低只有51.64%。此外, 經與揚子鱷FoxO4基因CDS區序列編碼的氨基酸比對發現兩者同源性僅為37.08%, 側面佐證所克隆序列確實為揚子鱷FoxO1基因編碼序列。
利用DNAman軟件構建脊椎動物FoxO1基因的分子進化樹結果表明(圖 1), 揚子鱷首先鱷目動物聚為一小類、再與斑胸草雀聚為第一大類、其他爬行類物種聚為第二大類, 提示鱷目物種FoxO1基因與鳥類的親緣關系相較中華鱉和原矛頭蝮等爬行動物更近。在其他脊柱動物中, 哺乳動物、硬骨魚和兩棲物種也各自聚類稱為子群, 表明FoxO1兼具功能保守性和種間特異性。
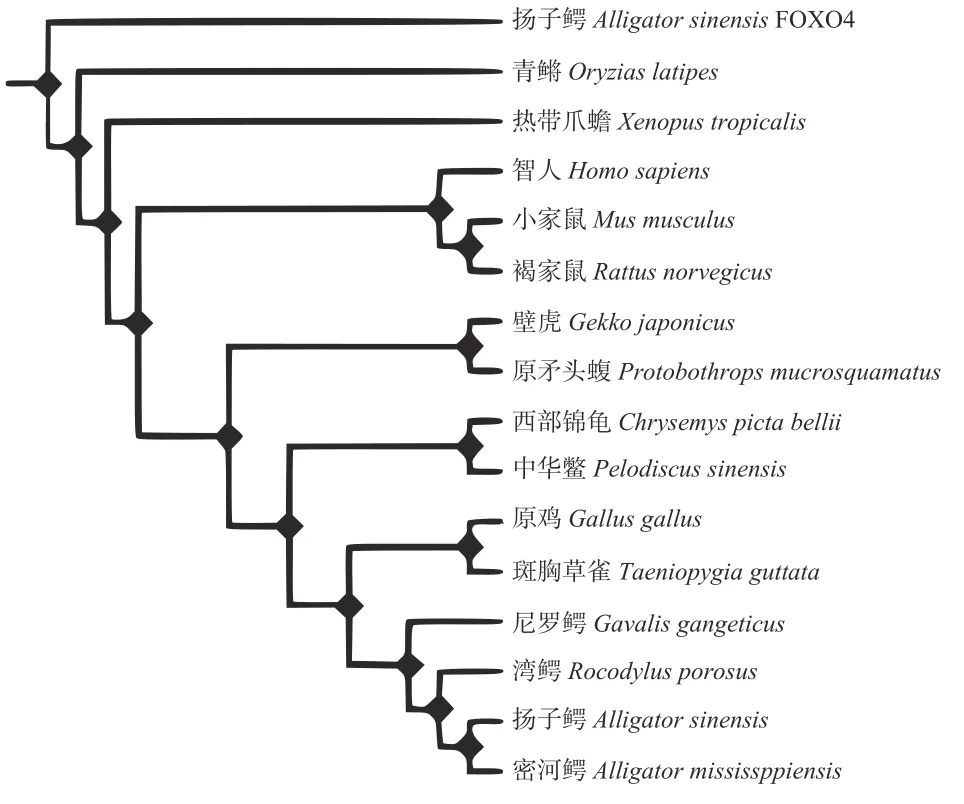
圖1 揚子鱷FoxO1基因氨基酸序列系統進化樹Fig. 1 Phylogenetic tree of amino acid sequence of FoxO1 gene in Alligator sinensis
2.3 免疫組織化學結果
FoxO1mRNA在初孵揚子鱷性腺復合體和外周組織的表達譜結果如圖 2所示, 在初孵揚子鱷外周組織中FoxO1mRNA表達情況為在大腸、肺臟和胃組織中有強或較強的表達, 在性腺復合體中有強表達。FoxO1蛋白在初孵揚子鱷性腺復合體和外周組織中的免疫組織化學結果如圖 3所示, 在光鏡下, 腎上腺小體由類固醇生成細胞群和嗜鉻細胞群組成。類固醇生成細胞群位于腎上腺小體周圍,細胞排列緊密、成索團狀分布、細胞呈立方形或柱狀, 胞質染色淺、呈網絡狀, 細胞核呈圓形或卵圓形、位于細胞中央、染色深。結果表明FoxO1蛋白在性腺復合體的皮質部、髓質部和腎上腺部位均有表達, 在性腺復合體中, FoxO1蛋白在腎上腺類固醇合成細胞中強表達, 在性腺皮質部表達量顯著高于髓質部(髓質與皮質部的間隔用虛線表示), 且其在性腺皮質部中的表達主要位于前體顆粒細胞(顆粒細胞核型呈卵圓形、前體顆粒細胞核型呈立方或不規則形)。其中腎上腺類固醇合成細胞和嗜鉻細胞中均呈強表達(圖 2B), 性腺皮質部的表達量顯著高于髓質部且其在性腺皮質部中的表達主要位于前體顆粒細胞和卵母細胞(圖 2C);FoxO1蛋白在肺間質中有較強表達, 肺泡外圍細胞有強表達(圖 2D); FoxO1蛋白在大腸內縱肌層及外縱肌層部分細胞有表達(圖 2E); 黏膜肌層部分細胞團及大腸絨毛上皮有分布均勻的較強表達(圖 2F);FoxO1蛋白在胃體的漿膜層、黏膜下層和環肌層均有較強表達(圖 2G)。

圖2 FoxO1在揚子鱷外周組織中的時空表達規律Fig. 2 Temporal and spatial expression of FoxO1 in the peripheral tissues of Alligator sinensis
2.4 FoxO1蛋白在不同出殼后日齡雌性揚子鱷性腺復合體中的時空變化規律
分別以17、63和96日齡雌性揚子鱷性腺復合體組織為對象, 對FoxO1蛋白的免疫熒光信號定位情況進行檢測。如圖 3所示, FoxO1蛋白在不同時期性腺組織中的免疫陽性信號在性腺皮質部、髓質部中、中腎和腎上腺等處均有不同程度表達, 但其表達模式存在時期差異。17日齡性腺組織中的FoxO1主要表達于腎上腺和中腎區, 且顯著高于63日齡和96日齡的性腺組織相應區域。FoxO1在17日齡性腺皮質部表達量極微弱, 隨著性腺發育過程表達逐漸上調, 在63日齡性腺皮質部呈無規則隨機分布, 在96日齡性腺皮質部中, 其熒光信號分布趨向于卵母細胞發育程度更高的位置。熒光定量結果表明17、63和96日齡性腺組織中的FoxO1基因mRNA豐度差異不顯著, 與此不同Western blot結果表明: 17、63和96日齡性腺組織中的FoxO1表達量呈先上升后下降的變化模式, 表現為63日齡FoxO1基因表達量顯著高于17日齡和96日齡。

圖3 FoxO1在揚子鱷性腺組織中的時空表達規律Fig. 3 Temporal and spatial expression of FoxO1 in the ovarian tissues of Chinese Alligator
3 討論
3.1 Forkhead box (Fox)蛋白家族的發現及其生物學功能
Forkhead box (Fox)蛋白家族因能引起果蠅“叉頭”突變而得名, 是一類DNA結合域具有翼狀螺旋結構的轉錄因子, 通過與靶基因啟動子中的叉頭轉錄因子識別位點結合, 進一步抑制或激活相關靶基因的轉錄從而發揮生物學功能。Fox含19個亞家族, 其中作為研究最為透徹的FoxO亞家族中成員中高度同源的FoxO1、FoxO3、FoxO4和FoxO6四個亞型, 已被證實在哺乳動物[14]和秀麗線蟲[15]等低等生物中廣泛存在。FoxO1是進化上保守的叉頭轉錄因子FoxO亞家族的成員, 有研究發現其在胰島素和生長因子對葡萄糖穩態和細胞分化等生命活動的影響過程中發揮重要作用[16], 且對葡萄糖代謝、細胞周期調控、細胞凋亡和氧化應激抵抗等過程也發揮重要影響。近年來對FoxO1的研究大多集中于小鼠、豬、牛、羊等哺乳動物, 尚未見關于該基因的序列和功能在鱷目動物中的相關報道。本研究通過揚子鱷FoxO1編碼區序列的克隆和序列特征分析, 檢測其在外周組織中的表達分布,結合其在性腺早期發育過程中的時空變化規律研究進一步探究其在揚子鱷卵子發生進程中的功能。
FoxO1是FoxO亞家族中發現最早的成員, 其與成肌細胞分化及脂肪細胞代謝有關, 并作為潛在因子參與骨骼肌的分化及Ⅰ型肌纖維基因的表達, 因此是研究肌肉及脂肪組織發育和代謝的重要候選因子[17]。FoxO1對肌細胞的分化及脂肪細胞的代謝有著一定的作用, 其能促進脂細胞的分化[18]、影響骨骼肌的生長發育[19]和Ⅰ型肌纖維基因的表達[20]。大量研究表明FoxO轉錄因子也是控制胚胎發生的重要物質, 對所有的胚原基層和器官的發育都是必需的[21]。在哺乳動物中, FoxO亞家族中的FoxO1、FoxO3和FoxO4廣泛表達于全身組織, 而FoxO6主要表達在腦的特定區域[22]。由此可推斷其在動物的生長發育方面也發揮了重要作用。楊燕軍[17]研究發現FoxO1基因在初生豬(1日齡)和成年豬(9月齡)的肝、肺、腎、脾、心、胃、皮下脂肪、肌肉等組織中均有表達, 只是表達豐度隨發育階段和組織的不同而具有差異。王玲[23]研究發現FoxO1基因在新生犢牛各個組織中如腹內脂肪、小腸淋巴結、肺、脾、胸腺、胃、腎、肝和心均有表達, 但在不同組織中的表達量有差異, 這可能與FoxO1基因在有機體各組織器官中的生物學效應有關。本實驗發現FoxO1蛋白在初孵鱷肺、胃和大腸中均有較強或強表達, 說明其在初孵鱷的生長、代謝中可能有著某些特定的功能, 如促進組織的生長發育與成熟、直接或間接介導各組織的協調工作等。但是對揚子鱷的相關研究尚未取得足夠證據, 相關問題仍需更多的研究去發現。
3.2 Forkhead Box (Fox)蛋白家族在雌性生殖系統中的功能報道
FoxO1在許多細胞類型中表達并參與了包括細胞增殖、凋亡、自噬和氧化應激抵抗在內的諸多生物學過程[24], 其中FoxO1已被證實能夠通過影響卵巢卵泡的啟動、發育、成熟及閉鎖過程影響動物的繁殖性能。有研究指出FoxO1在多種哺乳動物的卵泡中均有表達, 也有研究發現,FoxO1mRNA在卵巢中表達豐富[25], 此外, 對香豬[26]的卵巢卵泡細胞進行免疫組化時發現FoxO1在香豬卵泡的多個發育階段的顆粒細胞細胞質中都有表達;在原始卵泡、次級卵泡和三級卵泡中顆粒細胞的表達弱; 在大腔卵泡的顆粒細胞細胞質中有較強的表達; 在閉鎖卵泡的顆粒細胞中的表達強度減弱;凋亡顆粒細胞中沒有檢測到FoxO1的表達且有隨卵泡發育逐漸增強的趨勢。Tarnawa等[27]對多種哺乳動物使用免疫組化方法進行實驗, 發現FoxO1在不同動物卵泡的不同發育階段的顆粒細胞中表達情況均不同, 表現為原始卵泡前體顆粒細胞不表達,而在發育中及更高級發育卵泡顆粒細胞中特異表達。還有研究表明,FoxO1對小鼠[15]、牛[23]的卵泡細胞的凋亡起到促進作用, 過表達FoxO1后發現小鼠卵泡顆粒細胞的凋亡水平顯著增加。以上研究結果表明FoxO1對目前已知的部分哺乳動物的卵泡發育具有一定的調控作用。卵巢實質是由外周的皮質和中央的髓質構成, 皮質部中含有不同發育階段的卵泡。本研究發現FoxO1蛋白在初孵揚子鱷的性腺組織的皮質部及髓質部均有表達且皮質部表達量明顯高于髓質, 皮質部中主要表達于前體顆粒細胞, 這與毛文智等[28]在小尾寒羊的研究中指出卵泡閉鎖進程易發生于如次級卵泡和三級卵泡這兩類發育程度較高的卵泡中, 申明[15]以小鼠為研究對象得出FoxO1的高表達就會導致顆粒細胞的凋亡與卵泡閉鎖的結論相呼應, 皮質部中含有顆粒細胞以及發育程度較高的卵泡, 故FoxO1在皮質部有較高表達。在本研究中還發現FoxO1蛋白在性腺組織中的表達模式存在著時期差異, 在揚子鱷性腺發育進程中, FoxO1的表達逐漸趨向于卵母細胞發育程度更高的位置, 這與上文中所提到的多種哺乳動物不同發育階段卵泡顆粒細胞中表達情況的結果相接近[27]。由此猜想其對卵泡及其周圍組織的發育成熟具有一定影響, 可以推測FoxO1在爬行動物揚子鱷和其他哺乳動物的卵母細胞發育、顆粒細胞的增殖和凋亡以及卵泡閉鎖過程中扮演著重要的角色, 說明FoxO1在揚子鱷和其他哺乳動物的生殖生理調控方面發揮了一定的作用, 但是其調控機理還有待進一步研究。后續繼續追蹤FoxO1蛋白在雌性揚子鱷性腺發育各個階段的定位、遷移及定量表達信息并結合FoxO1蛋白編碼基因層面的相關分析, 可對FoxO1蛋白對揚子鱷性腺發育的影響有更深層的發現。
3.3 Forkhead Box (Fox)蛋白家族在雌性卵子發生過程中的功能報道
哺乳動物卵巢內卵子發生過程受到多種因素的影響, 如內分泌、旁分泌、自身調節因子及環境等的協同作用。Richards等[12]最早發現, 大鼠卵巢中FoxO轉錄因子的表達和活性受IGF-1、雌激素及促性腺激素調控, 表明這些內分泌因素會影響FoxO功能的發揮, 國內的研究者騰云以小鼠為研究對象發現FSH對閉鎖卵泡中FoxO1的表達有明顯的抑制作用[11], 進一步證實了Richards等[12]的結論。FoxO1磷酸化的抑制能增強離體培養小鼠顆粒細胞周期調節因子PCNA和Ccnd-2基因的表達,進而阻滯細胞發生凋亡[29], 而這一過程也是FSH通過AKT/FoxO1信號通路調控相關下游靶基因的轉錄水平完成的。可以說明,FoxO1基因的表達水平與體內激素、信號通路等密切相關。本研究對17、63和96日齡雌性揚子鱷性腺復合體FoxO1mRNA進行了熒光定量PCR檢測, 實驗結果表明17、63和96日齡性腺組織中的FoxO1基因mRNA豐度差異不顯著。同時對這3個時期的雌性揚子鱷性腺復合體FoxO1蛋白進行Western blot半定量檢測, 實驗結果表明FoxO1蛋白在初孵揚子鱷性腺組織中的表達量存在著時期差異。17、63和96日齡性腺組織中的FoxO1表達量呈先上升后下降的變化模式, 表現為63日齡FoxO1基因表達量顯著高于17日齡和96日齡。熒光定量PCR和 Western Blot結果存在差異, 可能原因分析如下兩點: (1)所研究的基因可能存在著轉錄后修飾; (2)所取實驗材料均為實驗室前期所培養孵化的揚子鱷, 在不同的卵子發生時期過程中的性腺復合體組織, 染色質結構會在配子發生過程中發生顯著的變化, 其相關基因的轉錄活性也會發生相應的改變; 而在染色質重構和基因轉錄調節中表觀遺傳修飾發揮了不可或缺的重要的作用,據此可以推測, 在揚子鱷的配子發生發育過程中,表觀遺傳信息有可能發生了較大的變化。FoxO1與顆粒細胞上FSH 受體密切相關, 鑒于FoxO1作為信號通路中的關鍵一員, 其表達量變化及與上下游基因的相互作用對卵子發生過程中有重要影響。在個體發育過程中, 其表達量水平會發生一定變化,呈現一定的時期差異, 但是其變化原因還有待進一步研究。推測隨著揚子鱷的發育, 體內有關雌性激素發生變化, 從而導致FoxO1轉錄因子的表達量發生變化, 進而調控雌性生殖生理過程。然而, 參與卵子發生過程中的一些信號通路, 尤其是一些新的信號轉導通路及其調節機制仍不明確, 探索這些信號通路在卵子發生, 以及FoxO1在揚子鱷生殖生理調控中的分子機制仍將是今后研究的熱點。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
