懷玉山三葉青兩個栽培種塊根內生菌、根際微生物群落分布及其化感物質的比較分析
2022-08-30 10:29:32夏瑾華劉彩艷劉佳儀劉金玲劉樂彭雅婷尹明華
山東農業科學 2022年7期
夏瑾華劉彩艷劉佳儀劉金玲劉樂彭雅婷尹明華
(1. 上饒師范學院生命科學學院,江西 上饒 334001;2. 上饒農業技術創新研究院,江西 上饒 334001;3. 上饒市三葉青保育與利用技術創新中心,江西 上饒 334001;4. 上饒市藥食同源植物資源保護與利用重點實驗室,江西 上饒 334001)
三葉青為葡萄科崖爬藤屬多年生蔓生藤本植物,學名三葉崖爬藤,俗名金線吊葫蘆、蛇附子、石抱子、石猴子、石老鼠等,主要分布于浙江、江西、福建、湖南、湖北、廣東、廣西等省(自治區),為我國特有珍稀藥用植物[1]。 三葉青毒副作用小,是西藥無法代替的天然“植物抗生素”,一般以塊根或全草入藥,可抗病毒、抗炎、鎮痛、解熱、保肝[2]。 在臨床上,三葉青還可用來治療各類炎癥,如病毒性腦膜炎、乙型腦炎、病毒性肺炎、黃疸性肝炎等[3];三葉青塊根對小兒高燒和癌癥也有顯著療效,對各類腫瘤如肺癌、肝癌、胃癌、腸癌、宮頸癌等具有抑制癌細胞增殖和促進癌細胞凋亡的作用[4]。
研究表明,植物根系和根際微生物之間的相互作用對于植物的生長和品質形成至關重要[5]。植物根系通過分泌酚酸等化感物質差異性地調節根際土壤微生物群落,促進致病菌的增殖同時使有益菌衰減[6]。 三葉青在石質土壤和非石質土壤中種植兩年,發現兩種土壤的細菌群落顯著不同,且與三葉青塊根尺寸相關。 與非石質土壤中的微生物群相比,石質土壤具有更大的細菌多樣性、共生網絡復雜性,以及更多的Actinobacteria(放線菌門)、Rokubacteria(己科河菌門)、Rhizobiales(根瘤菌)、Desulfarculaceae(脫硫藻科)和Chthonobacteriae(紅細菌)細菌類群。 此外,塊莖表層土壤中細菌類群的潛在功能與宿主的基因調控途徑密切相關,如植物激素生物合成、光合作用和生物脅迫抗性,對塊莖的起始和發育至關重要[7]。 周武等[8]從三葉青塊根的氯仿抽提物中分離出4-羥基-3-甲氧基苯甲酸、對羥基苯甲酸及十六烯酸甲酯和十八烯酸乙酯的混合物,生物活性測定表明三葉青塊根的主要化感物質為4-羥基-3-甲氧基苯甲酸和對羥基苯甲酸。 還有研究表明,內生菌可通過物質循環、能量轉換與宿主相互作用進而影響藥用植物內環境氧化還原狀態、酸堿度等特性,促進氨基酸、核酸、碳水化合物等養分的轉換與儲存,加快宿主藥用植物對養分的吸收及代謝,從而提高中藥材品質[9]。 王蕾臻[10]從三葉青塊根中分離獲得了多個內生菌,其中內生真菌TH15 可顯著促進三葉青塊根的生長和發育。 目前關于三葉青栽培種之間塊根內生菌、根際微生物群落分布及其化感物質的比較分析未見報道。 本試驗以懷玉山三葉青兩個栽培種‘懷玉1 號’和‘懷玉2 號’為研究對象,利用Illumina 高通量測序和LC-MS 技術分析其根際微生物和塊根內生菌的群落分布和化感物質的差異,整體把握兩者根際微生物和塊根內生菌資源狀況,尋找可以提高懷玉山三葉青有效成分含量的根際微生物和塊根內生菌資源,對提高懷玉山三葉青藥材品質和緩解化感自毒作用具有重要的現實意義。
1 材料與方法
1.1 試驗材料
懷玉山三葉青兩個栽培種懷玉1 號和懷玉2號大棚高地栽培的3年生塊根。
1.2 試驗方法
1.2.1 兩個栽培種塊根內生菌和根際微生物的測序 DNA 提取:兩個栽培種塊根和根際土壤均各取100 g,3 次重復。 采用PowerSoilTMDNA Isolation Kit 試劑盒提取DNA,并用1%瓊脂糖凝膠電泳進行檢測。
PCR 擴增:使用正向引物341F(5′-CCTAYGGGRBGCASCAG-3′) 和 反 向 引 物806R (5′-GGACTACNNGGGTATCTAAT-3′)[11,12]擴增細菌16S rRNA 基因的V3-V4 高變區。 擴增體系(20 μL):5×FastPfu緩沖液4 μL,dNTPs(2.5 mmol/L)2 μL,引物(5 μmol/L)0.8 μL,FastPfu聚合酶0.4 μL,DNA 模板2.5 μL,加ddH2O 至20 μL。 PCR反應條件:95℃3 min;95℃30 s,55℃30 s,72℃30 s,27 個循環;72℃10 min,4℃保存。 每個樣本3 個重復,將同一樣本的PCR 產物混合后用2%瓊脂糖凝膠電泳檢測,使用AxyPrepDNA 凝膠回收試劑盒(AXYGEN 公司)回收PCR 產物,并用2%瓊脂糖凝膠電泳進行檢測。
熒光定量:將PCR 產物用QuantiFluorTM-ST藍色熒光定量系統(Promega 公司)進行定量檢測。
Miseq 文庫構建:通過PCR 將Illumina 官方接頭序列添加至目標區域外端;使用凝膠回收試劑盒切膠回收PCR 產物;Tris-HCl 緩沖液洗脫后采用2%瓊脂糖凝膠電泳進行檢測;氫氧化鈉變性,產生單鏈DNA 片段。 文庫構建試劑盒為TruSeqTMDNA Sample Prep Kit。
Miseq 測序:DNA 片段的一端與引物堿基互補,固定在芯片上;以DNA 片段為模板,芯片上固定的堿基序列為引物,在芯片上PCR 合成目標待測DNA 片段;變性、退火后,芯片上DNA 片段的另一端隨機與附近的另外一個引物互補,也被固定住,形成“橋”(bridge); PCR 擴增,產生DNA簇;DNA 擴增子線性化成為單鏈;加入改造過的DNA 聚合酶和帶有4 種熒光標記的dNTP,每次循環只合成一個堿基;用激光掃描反應板表面,讀取每條模板序列第一輪反應所聚合上去的核苷酸種類;將熒光基團和終止基團化學切割,恢復3′端粘性,繼續聚合第二個核苷酸;統計每輪收集到的熒光信號結果,獲知模板DNA 片段的序列。
生物信息分析:Miseq 測序得到的PE reads首先根據overlap 關系進行拼接,同時對序列質量進行質控和過濾,區分樣本后進行OTU 聚類分析和物種分類學分析,基于OTU 聚類分析結果,對OTU 進行多樣性指數分析以及測序深度檢測;基于分類學信息,在各個分類水平上進行群落結構的統計分析。 在上述分析的基礎上,對多樣本的群落組成和系統發育信息進行多元分析和差異顯著性檢驗等。
1.2.2 兩個栽培種塊根化感物質的測定 樣品處理:精確稱取1000 mg 樣品(兩個栽培種塊根的根際土壤)至裝有一顆直徑6 mm 的研磨珠的2 mL 離心管中;加入1000 μL 含0.02 mg/mL 內標(L-2-氯苯丙氨酸) 的提取液[甲醇∶水=4∶1(V∶V)];冷凍組織研磨儀研磨6 min(-10℃,50 Hz);低溫超聲提取30 min(5℃,40 kHz);樣品于-20℃靜置30 min;13000× g 離心15 min(4℃),移取上清液,氮氣吹干;加入100 μL 復溶液(乙腈∶水=1∶1)復溶;渦旋混勻30 s,低溫超聲萃取5 min(5℃,40 kHz);13000× g 離心10 min(4℃),移取上清液至帶內插管的進樣小瓶中上機分析;另外,每個樣本分別移取20 μL 上清液,混合后作為質控樣本。
LC-MS 檢測:本次LC-MS 分析的儀器平臺為賽默飛公司的超高效液相色譜串聯傅里葉變換質譜UHPLC -Q Exactive HF-X 系統。 色譜條件:色譜柱為ACQUITY UPLC HSS T3(100 mm ×2.1 mm i.d.,1.8 μm; Waters,Milford,USA);流動相A 為95%水+5%乙腈(含0.1%甲酸),流動相B 為47.5%乙腈+47.5%異丙醇+5%水(含0.1%甲酸),進樣量為2 μL,柱溫為40℃。 質譜條件:樣品經電噴霧電離,分別采用正、負離子掃描模式采集質譜信號。 掃描范圍:70~1050 m/z;鞘氣流速:50 arb;輔助氣流速:1350 arb;加熱溫度:425℃;毛細管溫度:325℃;噴霧電壓(正模式):3500 V;噴霧電壓(負模式):-3500 V;SLens 電壓:50 V;碰撞能:20、40、60 eV;分辨率(Full MS):60000;分辨率(MS2):7500。
代謝產物鑒定:將原始數據導入代謝組學處理軟件Progenesis QI(Waters Corporation,Milford,USA)進行基線過濾、峰識別、積分、保留時間校正、峰對齊等,最終得到含保留時間、質荷比和峰強度等信息的數據矩陣。 其后采用該軟件進行特征峰搜庫鑒定,將MS 和MS/MS 質譜信息與代謝數據庫進行匹配,MS 質量誤差設置為小于10 mg/kg,同時根據二級質譜匹配得分鑒定代謝物。主要數據庫為http:/ /www.hmdb.ca/、https:/ /metlin.scripps.edu/等主流公共數據庫以及自建的數據庫。
2 結果與分析
2.1 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的測序信息
由表1 可知,兩個樣品有效序列數為287798,有效堿基數為65272 031,序列平均長度為226 bp。

表1 懷玉山三葉青兩個栽培種塊根內生菌、根際微生物的測序信息統計
2.2 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的OTU 分析
2.2.1 分類學分析 由圖1 可知,懷玉2 號塊根(LM_G)的物種數量為111,懷玉1 號塊根(WT_G)的物種數量為91;懷玉1 號根際土壤(WT)的物種數量為402,懷玉2 號根際土壤(LM)的物種數量為492。 表明懷玉1 號塊根內生菌和根際微生物的物種數量均少于懷玉2 號。
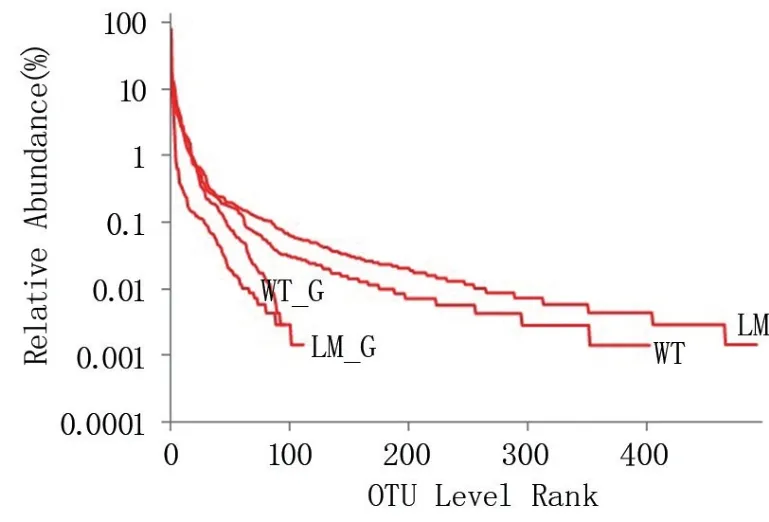
圖1 懷玉山三葉青兩個栽培種塊根內生菌和 根際微生物OTU 分析的Rank-Abundance 曲線
2.2.2 Pan/Core 物種分析 由圖2 可知,懷玉山三葉青兩個栽培種塊根內生菌和根際微生物序列Core OTU 隨著樣本數目的增加逐漸減少;Pan OTU 隨著樣本數目的增加逐漸增加。
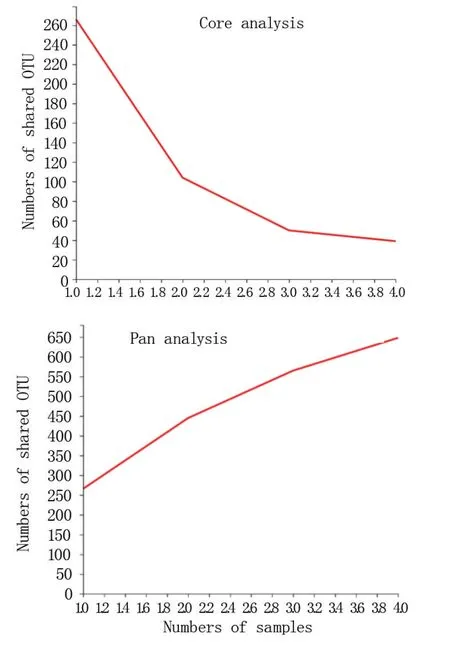
圖2 懷玉山三葉青兩個栽培種塊根內生菌和 根際微生物的Pan/Core OTU 分析
2.3 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的Alpha 多樣性分析
2.3.1 Alpha 多樣性指數 從表2 可知,懷玉1號塊根(WT_G)的Ace 指數和Chao 1 指數最高,其次是懷玉2 號根際土壤(LM),懷玉2 號塊根(LM_G)和懷玉1 號根際土壤(WT)較低,表明懷玉1 號塊根(WT_G)和懷玉2 號根際土壤(LM)的物種數較多。 懷玉1 號根際土壤(WT)的覆蓋度最高。 對于Shannon 指數,懷玉1 號塊根(WT_G)和懷玉2 號根際土壤(LM)較高,而懷玉2 號塊根(LM_G)和懷玉1 號根際土壤(WT)較低,表明懷玉1 號塊根(WT_G)和懷玉2 號根際土壤(LM)的物種多樣性較高。 對于Simpson 指數,懷玉2 號塊根(LM_G)最高,懷玉1 號塊根(WT_G)最低,表明懷玉1 號塊根(WT_G)的物種多樣性最高。
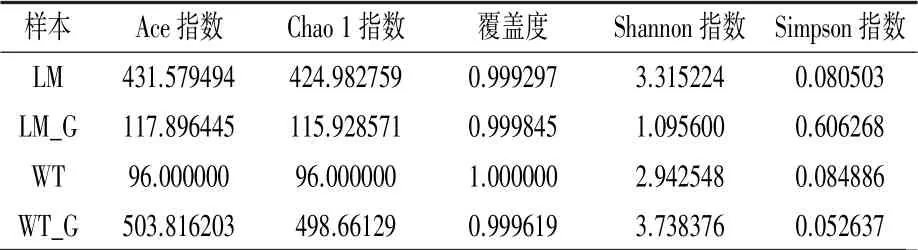
表2 懷玉山三葉青兩個栽培種塊根內生菌、根際微生物的Alpha 多樣性指數
2.3.2 稀釋曲線 由圖3 可知,懷玉1 號塊根(WT_G)、懷玉2 號根際土壤(LM)、懷玉2 號塊根(LM_G)和懷玉1 號根際土壤(WT)的稀釋曲線隨取樣量的增加均趨向平坦,說明測序數據量合理。
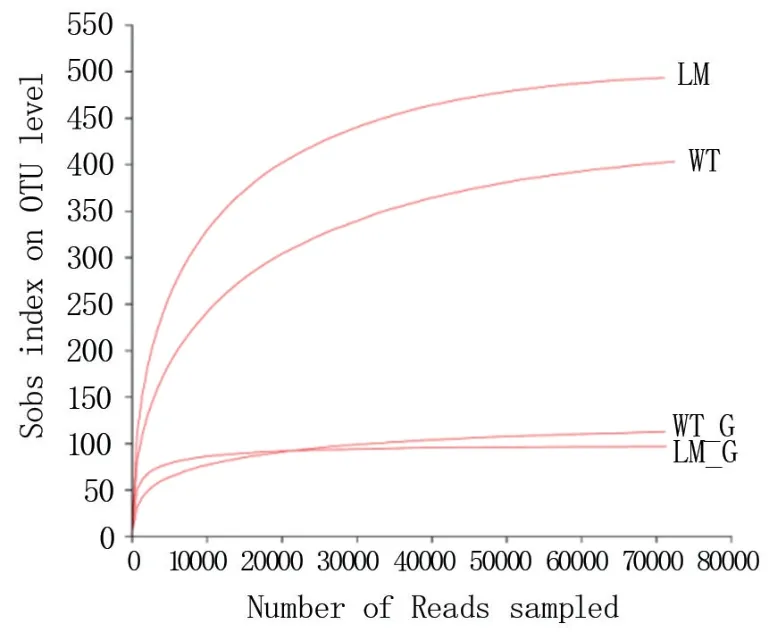
圖3 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的稀釋曲線
2.4 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的群落組成
懷玉山三葉青兩個栽培種塊根內生菌和根際微生物群落主要為子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)、羅茲菌門(Rozellomycota)、被孢霉門(Mortierellomycota,圖4)。 在懷玉2 號根際土壤(LM)中,子囊菌門(Ascomycota)占比64.87%,unclassified_k_Fungi 占比18.66%,擔子菌門(Basidiomycota)占比7.54%,羅茲菌門(Rozellomycota)占比0.93%,被孢霉門(Mortierellomycota)占比7.94%,others 占比0.06%;在懷玉1 號根際土壤(WT)中,子囊菌門(Ascomycota)占比63.18%,unclassified_k_Fungi 占比2.32%,擔子菌門(Basidiomycota)占比22.20%,羅茲菌門(Rozellomycota)占比2.83%,被孢霉門(Mortierellomycota)占比9.41%,others 占比0.05%;在懷玉2 號塊根(LM_G) 中,子囊菌門(Ascomycota) 占比96.88%,擔子菌門(Basidiomycota)占比3.08%,羅茲菌門(Rozellomycota) 占比0.01%,被孢霉門(Mortierellomycota) 占 比 0. 03%,others 占 比0.01%;在懷玉1 號塊根(WT_G)中,子囊菌門(Ascomycota)占比83.36%,unclassified_k_Fungi占比0. 07%,擔 子菌 門(Basidiomycota) 占 比15.43%,羅茲菌門(Rozellomycota)占比0.02%,被孢霉門(Mortierellomycota)占比7.94%,others 占比0.04%。表明懷玉2 號根際土壤(LM)和懷玉1號根際土壤(WT)的優勢群落均為子囊菌門(Ascomycota),且兩者的子囊菌門(Ascomycota)數量相差不大,懷玉1 號根際土壤(WT)的擔子菌門(Basidiomycota) 數量高于懷玉2 號根際土壤(LM);懷玉2 號塊根(LM_G)和懷玉1 號塊根(WT_G)的優勢群落也均為子囊菌門(Ascomycota),其次為擔子菌門(Basidiomycota),懷玉1 號塊根(WT_G)的擔子菌門(Basidiomycota)數量高于懷玉2 號塊根(LM_G),子囊菌門(Ascomycota)數量低于懷玉2 號塊根(LM_G)。
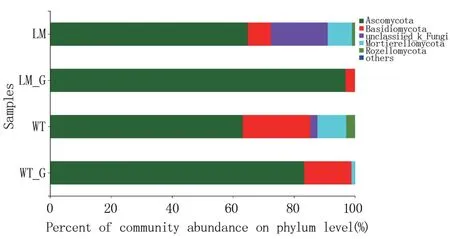
圖4 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的群落組成分析
2.5 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的樣本層級聚類分析
由圖5 可知,懷玉2 號根際土壤(LM)和懷玉1 號根際土壤(WT)聚為一個層級;懷玉2 號塊根(LM_G)和懷玉1 號塊根(WT_G)聚為另一個層級。
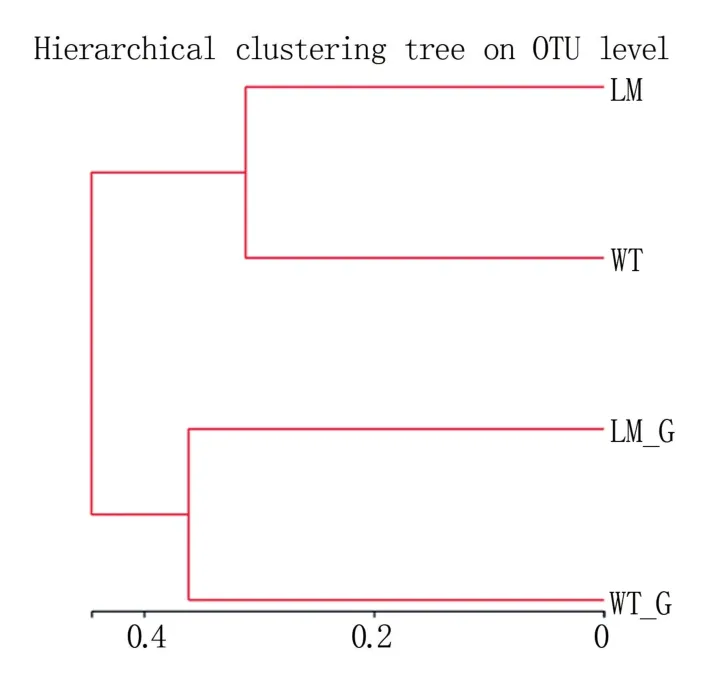
圖5 懷玉山三葉青兩個栽培種塊根內生菌和 根際微生物的樣本層級聚類分析
2.6 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的樣本距離熱圖
由圖6 可知,懷玉2 號根際土壤(LM)和懷玉2 號塊根(LM_G),懷玉1 號根際土壤(WT)和懷玉1 號塊根(WT_G)的距離最遠,顏色梯度代表的數值為1。 懷玉2 號根際土壤(LM)和懷玉1號根際土壤(WT),懷玉2 號塊根(LM_G)和懷玉1 號塊根(WT_G)的距離最近,顏色梯度代表的數值為0.6。
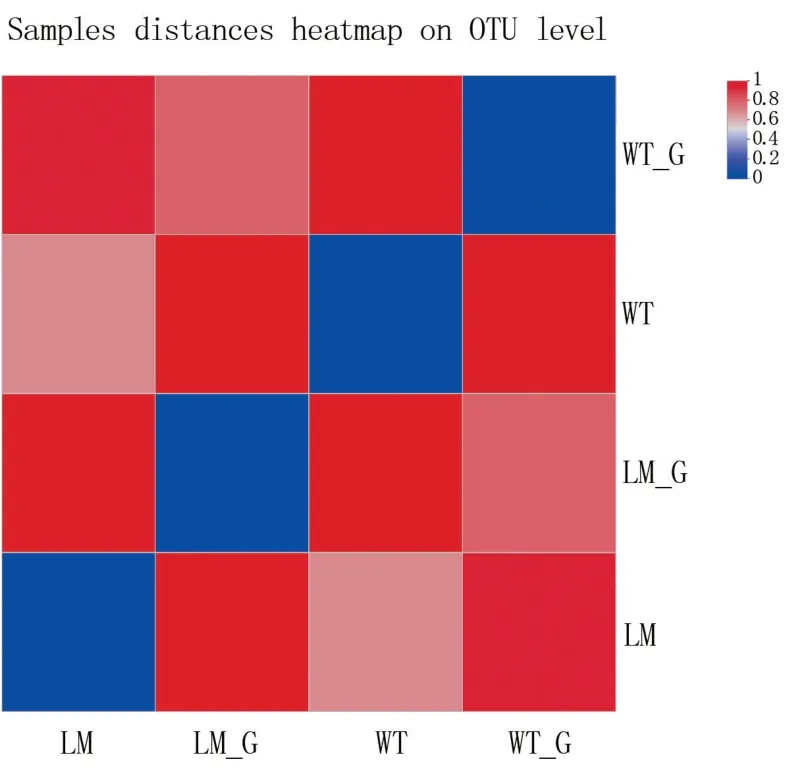
圖6 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的樣本距離熱圖
2.7 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的PCA 分析
由圖7 可知,懷玉2 號根際土壤(LM)和懷玉1 號根際土壤(WT)兩樣本點接近,表明兩樣本物種組成相似。 懷玉2 號塊根(LM_G)和懷玉1 號塊根(WT_G)兩樣本點較遠,表明兩樣本物種組成不同。
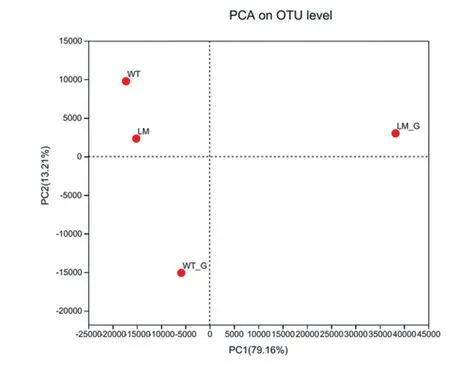
圖7 懷玉山三葉青兩個栽培種塊根內生菌和 根際微生物的PCA 分析
2.8 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的多物種差異檢驗
懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的多物種差異檢驗見圖8。 懷玉2 號根際土壤(LM)和懷玉1 號根際土壤(WT)子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)、unclassified_k_Fungi、被孢霉門(Mortierellomycota)、羅茲菌門(Rozellomycota)、球囊菌門(Glomeromycota)、 壺 菌 門( Chytridiomycota)、 捕 蟲 霉 門(Zoopagomycota)均具有顯著性差異,而Monoblepharomycota 的數量無顯著性差異。 懷玉2 號塊根(LM_G)和懷玉1 號塊根(WT_G)子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)、被孢霉門(Mortierellomycota)、unclassified_k_Fungi、球囊菌門(Glomeromycota)、羅茲菌門(Rozellomycota)、壺菌門(Chytridiomycota)的數量均具有顯著性差異。
2.9 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的KEGG 功能豐度
由圖9 可知,懷玉2 號塊根(LM_G)和懷玉1號塊根(WT_G)內生菌、懷玉2 號根際土壤(LM)和懷玉1 號根際土壤(WT)微生物KEGG 功能豐度較高的酶類主要有:EC3.6.1.3(腺苷三磷酸酶,adenosinetriphosphatase)、EC5.3.1.4(L-阿拉伯糖異構酶,L-arabinose isomerase)、EC3.2.1.3(葡聚糖1,4-α-葡萄糖苷酶,glucan 1,4-alpha-glucosidase)、EC2.7.7.6(DNA 定向RNA 聚合酶,DNAdirected RNA polymerase)、EC3.2.1.18(外顯α 唾液酸酶,exo-alpha-sialidase)、EC5.2.1.8(肽基脯氨酰異構酶,peptidylprolyl isomerase)、EC2.7.7.7(DNA 定向DNA 聚合酶,DNA-directed DNA polymerase)、EC2.3.1.48(組蛋白乙酰轉移酶,histone acetyltransferase)、EC3.6.3.14[H(+)-傳輸雙扇區ATP 酶,H(+)-transporting two-sector ATPase]、EC3.4.25.1(蛋白酶體內肽酶復合物,proteasome endopeptidase complex)、EC3.1.3.16(蛋白質絲氨酸/蘇氨酸磷酸酶,protein-serine/threonine phosphatase)、EC1.14.14.1(非特異性單加氧酶,unspecific monooxygenase)等。
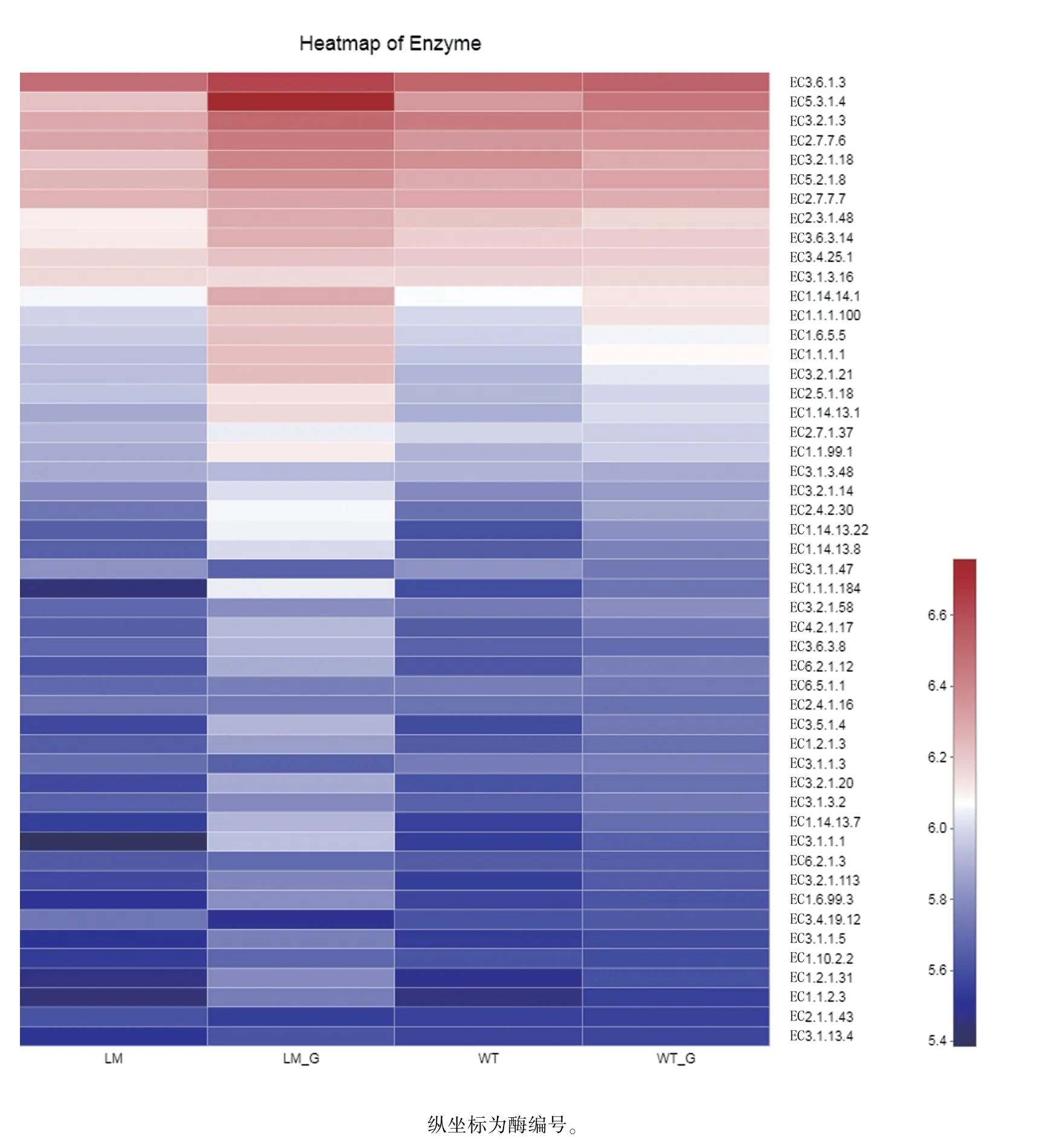
圖9 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的KEGG 功能豐度統計
2.10 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的FUNGuild 功能分類
由圖10 可知,懷玉2 號塊根(LM_G)和懷玉1 號塊根(WT_G)內生菌、懷玉2 號根際土壤(LM)和懷玉1 號根際土壤(WT)微生物的分類主要有:未定義腐生真菌(Undefined Saprotroph)、內生凋落物腐生菌土壤腐生菌未定義腐生真菌(Endophyte-Litter Saprotroph-Soil Saprotroph-Undefined Saprotroph)、動物病原內生真菌寄生蟲植物病原木腐生物(Animal Pathogen-Endophyte-Fungal Parasite-Plant Pathogen-Wood Saprotroph)、動物病原未定義腐生真菌(Animal Pathogen-Undefined Saprotroph)、動物病原體內生真菌地衣寄生蟲植物病原體土壤腐生真菌木材腐生真菌(Animal Pathogen-Endophyte-Lichen Parasite-Plant Pathogen-Soil Saprotroph-Wood Saprotroph)、真菌寄生蟲未定義腐生物(Fungal Parasite-Undefined Saprotroph)、動物病原體糞腐菌內生真菌植物腐生木(Animal Pathogen-Dung Saprotroph-Endophyte-Epiphyte -Plant Saprotroph-Wood)、動物病原體內生真菌地衣寄生蟲植物病原體木材腐生真菌(Animal Pathogen-Endophyte-Lichen Parasite-Plant Pathogen-Wood Saprotroph)、 動 物 病 原(Animal Pathogen)、植物病原(Plant Pathogen)、內生菌根植物病原菌未定義腐生真菌(Endomycorrhizal-Plant Pathogen-Undefined Saprotroph)、土壤腐生真菌(Soil Saprotroph)、內生凋落物腐生真菌木材腐生真菌(Endophyte-Litter Saprotroph-Wood Saprotroph)等。

圖10 懷玉山三葉青兩個栽培種塊根內生菌和根際微生物的FUNGuild 功能分類統計
2.11 懷玉山三葉青兩個栽培種根際土壤化感物質分析
圖11顯示,在該檢測條件下,峰形良好,分布相對均勻。

圖11 懷玉山三葉青兩個栽培種根際土壤 代謝物LC-MS 總離子色譜圖
通過LC-MS 技術對懷玉山三葉青兩個栽培種根際土壤進行檢測,共鑒定出6 種酚酸類化感物質(表3)。 懷玉1 號根際土壤(WT)的龍膽酸、香草醛酸和水楊酸含量高于懷玉2 號根際土壤(LM),而丁香酸、沒食子酸和阿魏酸含量低于懷玉2 號根際土壤(LM)。
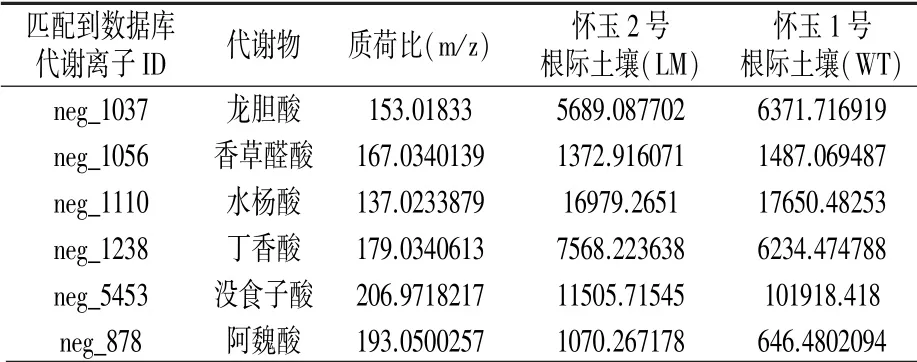
表3 懷玉山三葉青兩個栽培種根際土壤化感物質的比較分析
3 討論與結論
研究表明,根際微生物-植物-環境之間互作關系較為敏感,根際土壤微生物的多樣性、生理活性及微生物間的相互作用決定植物的品質和產量[13]。 高通量測序具有成本低、通量高等優點,在微生物生態學研究中得到了廣泛應用。 本試驗通過Miseq 測序平臺研究了懷玉山三葉青兩個栽培種根際土壤微生物的群落結構組成,為后續懷玉山三葉青化感效應研究奠定了基礎。 唐彬彬等[14]發現,在真菌群落中,子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)、 壺菌門(Chytridiomycota)和接合菌門(Zygomycota)在三七根際土壤群落中屬于優勢菌群。 蘇小惠等[11]發現,5 個苧麻品種根際土壤真菌中子囊菌門、接合菌門和擔子菌門為優勢門。 楊瀟湘等[12]發現大豆和油菜根際土壤在門水平的優勢類群相同,如子囊菌門、接合菌門、擔子菌門和壺菌門等的豐度都較高,但在屬水平上兩種樣品之間具有顯著差異。本試驗中,懷玉2 號根際土壤(LM)和懷玉1 號根際土壤(WT)的優勢群落均為子囊菌門(Ascomycota),其次為擔子菌門(Basidiomycota),與上述結果一致。 Tan[15]、吳照祥[16]等發現,三七根際有益真菌被孢霉門(Mortierellomycota)的豐富度隨三七種植年限的增加而減少,可能使得真菌屬水平上種群失衡引起一些病原真菌如子囊菌門和擔子菌門的種群豐富度增加,從而導致三七發生連作障礙。 本試驗中,懷玉2 號塊根(LM_G)子囊菌門的數量顯著多于懷玉1 號塊根(WT_G),懷玉2 號塊根(LM_G)擔子菌門(Basidiomycota)的數量顯著低于懷玉1 號塊根(WT_G),懷玉2號根際土壤(LM)有益真菌被孢霉門(Mortierellomycota) 的數量顯著低于懷玉1 號根際土壤(WT)。 懷玉山三葉青兩個栽培種的塊根內生菌和根際土壤微生物的病原真菌和有益真菌群落在同一栽培種內分布較為一致。
植物內生菌菌群結構組成隨物種及其基因型、生長環境不同而異[17]。 內生菌以植物為宿主載體,二者相互作用形成微生態系統,間接影響植物的品質和產量[18]。 蔡媛等[19]利用MiSeq 高通量技術,發現湖南、安徽、江西、湖北、貴州5 個產地15 組多花黃精內生菌數量和種類豐富,其內生真菌的優勢菌群為子囊菌門下一未知鑒定菌屬,其次為赤殼屬、炭疽屬、間座殼屬、鐮刀菌屬等。 楊立昌等[20]基于高通量測序,探明金釵石斛種子的內生真菌序列歸為14 個OTUs,隸屬于子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)和接合菌門(Zygomycota ),其中子囊菌門為優勢門。 雷鋒杰等[21]發現,人參根部相對豐度最高的內生真菌類群為子囊菌門(Ascomycota),占各人參樣品的80%以上,是人參內生真菌中最主要的真菌類群,其次是擔子菌門(Basidiomycota),其相對豐度比例較高。 本試驗結果與上述結果一致。
三葉青已被證實具有化感自毒作用,主要化感物質為4-羥基-3-甲氧基苯甲酸和對羥基苯甲酸[8]。 本試驗用GC-MS 技術檢測出6 種化感物質,其中懷玉1 號根際土壤(WT)的龍膽酸、香草醛酸和水楊酸含量高于懷玉2 號根際土壤(LM),而丁香酸、沒食子酸和阿魏酸含量低于懷玉2 號根際土壤(LM)。 說明懷玉山三葉青兩個栽培種懷玉1 號和懷玉2 號造成化感自毒和連作障礙的物質有所差異。
