長清茶新梢中細胞色素P450 基因CYP94B3 的季節表達響應
2022-08-30 10:29:18宋來超劉芳彬王晨羽丁兆堂孫海偉馬青平
山東農業科學 2022年7期
宋來超,劉芳彬,王晨羽,丁兆堂,孫海偉,馬青平
(1. 聊城大學農學院,山東 聊城 252000;2. 山東省農業科學院茶葉研究所,山東 日照 276800;3. 泰安市農業科學研究院茶葉研究所,山東 泰安 271001)
細胞色素P450 (cytochrome P450,簡稱CYP450)為一種B 族細胞色素,其輔基為血紅素,屬于強大的超基因家族蛋白酶之一,內質網與線粒體內膜是真核生物中CYP450 存在的主要場所。 CYP450 最早發現于大鼠肝微粒體中,1969年Frear 等[1]從棉花(Gossypium hirsutumL.)中發現了CYP450,這是首次發現植物細胞中存在CYP450。 隨后,人們分別從擬南芥(Arabidopsis thalianaL.)[2]、水稻(Oryza sativaL.)[3]、小麥(Triticum aestivumL.)[4]等植物中發現了CYP450的存在。
不同家族CYP450 的氨基酸序列具有一定的保守性,即CYP450 在不同物種之間具有同源性,其三級結構均由N 端的β 折疊和C 端的α 螺旋構成。 在所有CYP450 的典型保守結構域中,C末端血紅素的結合域含有FXXGXRXCXG 保守序列,這是鑒定CYP450 的主要特征結構,其中的半胱氨酸(Cys)具有完全保守性;K 螺旋區域中具有保守序列EXXR,其中谷氨酸(Glu)和精氨酸(Arg)完全保守;PDR 結構域中具有保守序列PXRX。
CYP450在植物不同組織、不同發育時期及不同脅迫條件下的表達存在較為明顯的差異。F3′H基因屬于CYP75B2 家族基因,茶樹F3′H基因在第一葉、第二葉、第三葉、第四葉、嫩莖、根、老葉中的表達水平依次降低,在根和老葉中幾乎不表達[5]。 崔會婷[6]研究發現蒺藜苜蓿葉中的Mt-CYP450基因表達量最高,且從嫩苗期到結果期呈先上升后下降的趨勢。 茶樹CYP71A26與CYP71B34基因受蟲害脅迫時分別表現為上調和下調表達,而受冷熱脅迫時則分別表現為下調和上調表達[7];受干旱、高溫、氮脅迫、鹽脅迫的影響,高羊茅P450 基因的表達表現為上調趨勢[8]。
CYP450 在植物體內起重要作用,主要分為兩類:一是參與多種植物內源性物質的代謝與合成,在苯丙烷代謝產物木質素、黃酮、異黃酮等物質的合成過程中發揮重要作用,且其在黃酮生物合成中的作用底物廣泛,對同一位點結構相似的化合物能夠同時進行催化[9]。 研究表明,鹽和植物根際促生菌(PGPR) 接種聯合處理誘導的CYP94B3基因可參與茉莉酸(JA)代謝[10];煙草香氣物質西柏三烯二醇的合成受CYP71D16基因的催化[11];CYP71AU87可催化萜類的合成與代謝過程[12];肉桂酸羧化酶(C4H)屬于CYP450 家族酶系,滇水金鳳(Impatiens uliginosaFranch)的C4H1、C4H2基因可催化反式肉桂酸轉化為香豆酸[13];紫蘇和蝴蝶草的CYP93B1、CYP93B2基因可參與黃酮合成酶Ⅱ的合成并發揮最重要的作用,其中黃酮合成酶Ⅱ屬于CYP93 亞家族,是黃酮類化合物合成過程中的關鍵酶;花青素、縮合單寧化合物的合成離不開CYP75 的參與,CYP75 對黃酮化合物的C3′位、C5′位上的羥基化具有催化作用[14];CYP93B 亞家族能夠催化黃酮類化合物C2 位羥基化并使其脫水,形成C =C[15],如CYP93B10 和CYP93B11 可將黃酮類物質轉化為2-羥基黃酮。 二是參與生物體代謝解毒。 植物生長過程中常會受到多種病蟲的侵害或化學藥品的毒害,細胞色素P450 作為解毒過程中的關鍵酶能將除草劑、殺蟲劑等外源化學物質轉化為無毒物質。 研究發現,棉鈴蟲CYP9A 亞家族6 個成員及CYP6B 亞家族四個成員能代謝殺蟲劑菊戊酸酯[16],多個細胞色素P450 的過表達可使致倦庫蚊(Culex quinquefasciatus)種群對氯菊酯類殺蟲劑的抗性增強[17]。
茶樹[Camellia sinensis(L.) O. Kuntze]是山茶科山茶屬中種植最為廣泛的植物,也是三大飲品之一。 黃酮類化合物是它的次生代謝產物之一,其骨架為C6-C3-C6,主要包括黃酮、異黃酮、花青素等天然化合物。 黃酮類化合物在茶葉中含量豐富,可影響茶葉的品質、滋味以及色澤等,其中黃酮醇類物質可增強咖啡堿的苦味,同時使茶湯干燥柔和,有更好的收斂感[18],其含量決定著綠茶的湯色。 CYP450 作為“萬能生物催化劑”,可參與黃酮類化合物的生物合成,間接影響茶葉的品質。
本試驗以不同季節的長清茶葉片為材料提取RNA,反轉錄合成cDNA,通過轉錄組分析(登錄號PRJNA830980)篩選得到CsCYP94B3基因,將茶樹及與其同源性較高的26 個物種的CYP94B3進行蛋白序列比對并構建系統進化樹,運用實時熒光定量PCR 技術測定不同季節CsCYP94B3的相對表達量,以期為長清茶采摘及品質提高提供一定的理論依據。
1 材料與方法
1.1 植物材料
長清茶鮮葉采自濟南市長清區立泰山、圣虎山、南湖玉露3 個茶園中,采摘時間均為2020年春季(5月)、夏季(7月)和秋季(9月)晴朗無風天氣的9 時,采摘的葉片均為一芽二葉、長勢良好、生長均勻、無病蟲害。 葉片采摘后用2 mL 離心管封存,立即于液氮中速凍,然后置于-80℃冰箱中保存備用,樣品設置3 次生物學重復。
1.2 試劑與設備
華越洋快速通用植物RNA 提取試劑盒、Thermo Scientific RevertAid First Strand cDNA Synthesis Kit、2000 DNA Marker、2×TaqPlus Master Mix (Dye Plus)、5×DNA Loading buffer、Biomarker 2×SYBR Green Fast qPCR Mix、瓊脂糖,PCR 儀、離心機、高通量組織研磨儀、紫外透射儀、金屬浴、渦旋振蕩器、超微量紫外分光光度計。
1.3 總RNA 提取與第一鏈cDNA 合成
使用高通量組織研磨儀將經過液氮速凍的長清茶葉片研磨成粉末,按照華越洋快速通用植物RNA 提取試劑盒說明書的操作步驟提取總RNA。總RNA 在1%瓊脂糖凝膠電泳上檢測完整度,并使用超微量紫外分光光度計測其濃度和純度。 吸取1 μg RNA,用Thermo Fermentas 反轉錄試劑盒K1622 合成第一鏈cDNA。 逆轉錄的反應條件為42℃溫育60 min,70℃反應5 min(此步反應目的為使逆轉錄酶失活),反應結束后將制備好的cDNA 置于-20℃冰箱保存備用。
1.4 CYP94B3 生物信息學分析
將通過轉錄組分析(登錄號PRJNA830980)篩選得到的CsCYP94B3基因,通過NCBI 中的BLAST 進行CsCYP94B3基因的同源性比對,獲得與該基因同源性較高的榴蓮(Durio zibethinus)、葡萄(Vitis vinifera)、銀白楊(Populus alba)、柑橘(Citrus sinensis)、歐洲木犀欖(Olea europaeasubsp.europaea)、小果咖啡(Coffea arabica)、番茄(Solanum lycopersicum)、野生煙草(Nicotiana attenuata)、可可樹(Theobroma cacao)等26 個物種的蛋白序列,并將CsCYP94B3基因序列通過DNAMAN 軟件翻譯成蛋白序列。 使用ExPASy 網站分析CYP94B3 氨基酸的組成、理化性質及疏水性。 選取葡萄、榴蓮、可可樹3 個代表性物種與茶樹的CYP94B3 蛋白序列使用DANMAN 軟件進行多重序列比對。 將茶樹與其他26 個物種的CYP94B3 蛋白序列使用MEGA-X 中的鄰位相連法(Neighbor-joining)-泊松模型構建系統進化樹。
1.5 實時熒光定量PCR
利用實時熒光定量PCR 測定CsCYP94B3基因在春、夏、秋季茶葉中的表達量。 PCR 總體系20 μL:cDNA 2 μL,10 μmol/L 上下游引物各0.5 μL (F:5′-AGCACATTCTCAAGACCAACTTC-3′,R:5′-ACTCGTGACTCGCTAACTTCC-3′),無菌水7 μL,Biomarker 2×SYBR Green Fast qPCR Mix 10 μL。 反應程序為:95℃3 min;95℃5 s,60℃30 ~40 s,共45 個循環。CsGAPDH基因為內參基因,CsCYP94B3基因的相對表達量用2-ΔΔCt法計算,統計分析使用SPSS 18.0 和Microsoft Excel 2010 軟件進行,差異顯著性分析基于Duncan’s 多重比較的單因素方差分析進行,P<0.05 表示兩兩之間差異顯著。
2 結果與分析
2.1 茶樹CYP94B3 蛋白理化性質及疏水性分析
根據ExPASy 網站分析結果,CsCYP94B3 的蛋白質分子量為58.79 kDa,理論等電點為8.76;氨基酸殘基數為518 個,堿性氨基酸的平均含量為15%,酸性氨基酸的平均含量為10%,芳香族氨基酸約占10%,雜環族氨基酸約占9%,脂肪族氨基酸約占81%;不穩定指數為41.51,屬于不穩定蛋白;總平均疏水性-0.052,為可溶性蛋白。 疏水性分析(圖1)發現,CsCYP94B3 蛋白疏水性在第16 位亮氨酸(Ile)具有最大值2.900,在第32位組氨酸(His)具有最小值-3.411,為親水蛋白。
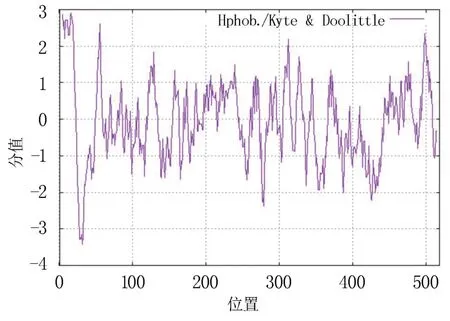
圖1 茶樹CYP94B3 蛋白疏水性分析
2.2 不同物種CYP94B3 蛋白多重序列比對分析
多重序列比對(圖2)發現,茶樹CYP94B3 蛋白序 列 與 葡 萄、 榴 蓮、 可 可 樹3 個 物 種 的CYP94B3 蛋白序列一致性為80. 41%,且其CYP94B3 蛋白序列都含有PDR 結構域(PXRX)、K 螺旋(EXXR)以及C 末端的血紅素結合域(FXXGXXXCXG),其中血紅素結合域是鑒別細胞色素P450 基因的主要特征結構,因此,茶樹、葡萄、榴蓮、可可樹的CYP94B3基因均為細胞色素P450 亞家族基因。
2.3 茶樹與其他物種的CYP94B3 進化樹構建
將茶樹與歐洲木犀欖、小果咖啡、煙草等26個物種的CYP94B3 蛋白序列使用MEGA-X 軟件中的鄰位相連法(Neighbor-joining)-泊松模型構建系統進化樹,結果(圖3)顯示,茶樹與中華獼猴桃、大桉、歐洲木犀欖、小果咖啡、番茄、野生煙草6 個物種的親緣關系較近,與其他20 個物種的親緣關系較遠。

圖3 茶樹及其他物種CYP94B3 的系統進化樹分析
2.4 CsCYP94B3 在不同季節長清茶新梢中的表達變化
CsCYP94B3基因在3 個茶園長清茶樹的春、夏、秋新梢中均可表達,但不同季節、不同地理條件間存在差異(圖4)。 在立泰山、圣虎山、南湖玉露3 處茶園茶樹中,CsCYP94B3基因在夏季茶樹新梢中的相對表達量明顯高于春秋兩季,且與圣虎山茶園春季、南湖玉露茶園春秋兩季差異達顯著水平。 圣虎山和南湖玉露茶園的茶樹新梢中CsCYP94B3基因相對表達量秋季略高于春季,但差異不顯著;立泰山茶園茶樹新梢中CsCYP94B3基因相對表達量春秋兩季相當。 表明CsCYP94B3基因的表達與環境條件有關,可響應季節變化,表現為夏季上調表達。
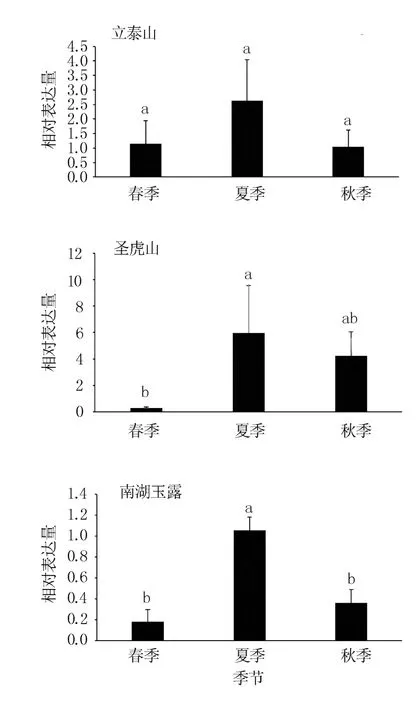
圖4 不同茶園不同季節長清茶中CYP94B3基因的相對表達量
3 討論與結論
P450 廣泛存在于動植物等細胞中,同一種植物的不同組織中都有分布但含量各不相同,其基因的表達量受到不同外界因素的調控[19]。 本研究結果表明,茶樹與中華獼猴桃、大桉、歐洲木犀欖、小果咖啡、番茄、野生煙草的CYP94B3 親緣關系較近,而與其他20 個物種親緣關系較遠,這種現象可能與細胞色素P450 堿基易突變有關。 前人研究發現,CYP704B2基因在第4 個外顯子處常發生多對堿基的替換或缺失,從而造成氨基酸的轉變或缺失[20]。 因此,不同物種CYP94B3 常在系統進化樹上發生分離。
本試驗通過實時熒光定量PCR 分析了不同茶園不同季節長清茶CYP94B3基因的相對表達量,結果發現,3個茶園的長清茶CYP94B3基因均在夏季表達量最高,推測夏季溫度高、光照強,長清茶生長代謝最旺盛,需要更多的細胞色素P450參與多種內源物質的合成與代謝反應。
綜上所述,茶樹CYP94B3基因的表達受季節因素調控,夏季上調表達,且南湖玉露和圣虎山的CsCYP94B3基因表達量夏季明顯高于春秋季。 這將為長清茶采摘及品質提高提供一定的參考,并為通過人工干預給茶樹創造適宜的生長環境以提高茶葉品質和產量奠定基礎。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
四川蠶業(2021年3期)2021-02-12 02:38:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
中成藥(2018年11期)2018-11-24 02:57:00
汽車觀察(2018年10期)2018-11-06 07:05:26
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
中成藥(2017年4期)2017-05-17 06:09:50
