賞食兼用百合京鶴高效組培再生體系的優化
2022-08-27 03:52:14祁先宇張文亮楊盼盼徐雷鋒
中國蔬菜 2022年8期
祁先宇 張文亮 楊盼盼 徐雷鋒* 明 軍*
(1 中國農業科學院蔬菜花卉研究所,北京 100081;2 沈陽農業大學生物科學技術學院,遼寧沈陽 110866)
百合(spp.)是具有較高營養價值的高檔蔬菜,也是觀賞花卉,還是我國重要的藥用植物(龍雅宜和張金政,1998)。京鶴是中國農業科學院蔬菜花卉研究所選育的具有自主產權的賞食兼用型優質百合品種,不僅抗逆性較強,而且鱗莖營養成分和生物活性物質含量也較高,具有很高的經濟價值,亟須進行大規模繁殖示范推廣(袁素霞 等,2016)。
分球繁殖和鱗片扦插是百合生產上常用的繁殖方式,但是這些繁殖方式繁殖系數低,且易積累病毒,影響百合的產量和品質(馬生軍 等,2018)。植物組織培養是一種可以保障作物品種優良特性的高效、現代化、工廠化原種生產方法。在百合組織培養中,鱗莖不定芽的分化培養是提高百合組培苗繁殖系數的關鍵步驟。常規的不定芽誘導分化是依據百合品種差異選用不同的細胞分裂素與生長素,不定芽誘導率較低,介于2.00%~4.88%之間(蘇菁華,2014;Islam et al.,2017;Sharifi et al.,2017;陳姝男,2018;張旭紅 等,2018;Gao et al.,2018;Mohebodini et al.,2018;Sahoo et al.,2018;李茂娟 等,2019;李卓憶,2019;張文婷 等,2019;Royandazagh,2019;符勇耀 等,2020;Patil et al.,2021;Yang et al.,2021)。
草甘膦作為目前使用最廣泛、安全的除草劑之一(Krimsky,2022),雖然其作用機制的研究已經比較清楚(Amrhein et al.,1980;Stephen,2019),但是目前未見草甘膦在百合及其他植物組織培養再生方面的研究報道。中國農業科學院蔬菜花卉所百合課題組前期在抗草甘膦轉基因京鶴百合的試驗研究中,發現低濃度草甘膦對京鶴鱗片不定芽誘導具有顯著的促進作用,且誘導率顯著高于常規組織培養再生分化體系;本課題組雖然建立了常規的京鶴不定芽誘導體系,但誘導系數(4.32)、生根系數(14.78)仍不高,且沒有進行鱗莖增殖方面的試驗。因此,本試驗首次使用草甘膦為誘導分化劑,以鱗片為外植體,優化京鶴高效組培再生體系,以期為百合鱗片不定芽誘導、組培苗生產提供理論參考。
1 材料與方法
1.1 試驗材料
試驗于2018 年4 月在中國農業科學院蔬菜花卉研究所進行。供試百合品種京鶴為中國農業科學院蔬菜花卉研究所自主培育,所選材料為秋季采收的生長健壯、無病蟲害的種球中層鱗片,種球直徑10~15 cm。試驗用草甘膦(Glyphosate)含量為95%(德國,Sigma,P9556-5G)。
1.2 試驗方法
采用MS 固體培養基〔MS 基本培養基(含維生素),瓊脂7 g·L,pH 值5.8〕,于121 ℃條件下滅菌20 min。培養室溫度為(25 ± 1)℃,暗培養。
1.2.1 草甘膦誘導京鶴鱗片不定芽試驗 以本課題組已有的京鶴不定芽分化培養基(MS+1.0 mg·L6-BA+0.1 mg·LNAA+30 g·L蔗糖)為基礎,設置4 個草甘膦濃度處理:CK1,0 mg·L;T1,2.5 mg·L;T2,5.0 mg·L;T3,7.5 mg·L。取京鶴健康種球鱗片,洗凈后用10%次氯酸鈉溶液浸泡滅菌15 min,再用無菌蒸餾水清洗3 遍,然后用解剖刀切割成1 cm × 1 cm 的小方塊,接種于添加不同濃度草甘膦的分化培養基上。每處理3 次生物學重復,每重復接種30 個外植體;置于室溫25℃條件下進行暗培養,40 d 后統計不定芽數,計算不定芽誘導系數。
不定芽誘導系數=不定芽數/接種的外植體數
1.2.2 6-BA、NAA 對京鶴不定芽增殖的影響 鱗片接種培養40 d 后,將上一試驗最佳條件下誘導產生的不定芽轉接到添加不同濃度6-BA 與NAA的培養基中進行增殖培養(MS+30 g·L蔗糖)。設置6 個6-BA 與NAA 濃度處理組合:J1,0.5 mg ·L6-BA+0.1 mg·LNAA;J2,0.5 mg·L6-BA+0.2 mg·LNAA;J3,1.0 mg·L6-BA+0.1 mg·LNAA;J4,1.0 mg·L6-BA+0.2 mg·LNAA;J5,2.0 mg·L6-BA+0.1 mg·LNAA;J6,2.0 mg·L6-BA+0.2 mg·LNAA。每處理3次生物學重復,每重復接種30 個外植體;置于室溫25 ℃、暗培養條件下進行增殖培養,60 d 后統計再生成的小鱗莖數,計算不定芽增殖系數。
增殖系數=再生小鱗莖數/接種外植體數
1.2.3 蔗糖對京鶴不定芽鱗莖膨大的影響 在MS培養基的基礎上,設置4 個蔗糖濃度處理:Z1,30 g·L;Z2,60 g·L;Z3,90 g·L;Z4,120 g ·L。將上一試驗最佳條件下得到的直徑1 cm 左右的小鱗莖轉接至不同蔗糖濃度的培養基中,每處理3 次生物學重復,每重復接種30 個小鱗莖;置于室溫25 ℃、暗培養條件下進行鱗莖膨大培養,30 d 后轉接1 次,60 d 后統計各鱗莖的直徑。
1.2.4 NAA 對京鶴膨大鱗莖生根的影響 在1/2MS+30 g·L蔗糖培養基的基礎上,設置4 個NAA 濃度處理:CK2,0 mg·L;G1,0.1 mg·L;G2,0.2 mg·L;G3,0.3 mg·L。將最佳條件下膨大培養60 d 后的鱗莖接種到添加不同濃度NAA的生根培養基中,每處理3 次生物學重復,每重復接種30 個膨大鱗莖;置于室溫25 ℃條件下進行暗培養,30 d 后統計所有鱗莖的總根數,計算生根系數。
生根系數=總根數/接種的鱗莖數
1.3 數據處理
利用Microsoft Excel 軟件進行試驗數據分析,使用SPSS Statistics 17.0 軟件進行顯著性檢驗。
2 結果與分析
2.1 草甘膦對京鶴鱗片不定芽誘導分化的效果
不同濃度的草甘膦對京鶴鱗片不定芽的誘導分化有顯著影響,鱗片內側密集分化產生了大量小仔球,而常規的不定芽分化培養基處理(CK1)只是在傷口處形成小仔球(圖1)。

圖1 草甘膦對京鶴單個鱗片不定芽的誘導分化
從表1 可以看出,T1 處理的誘導系數最高,為6.46,顯著高于其他處理;其次是CK1 和T2 處理,T3 處理最低。據觀察,T2 和T3 處理的部分外植體有褐化死亡的情況,尤其是T3 處理,褐化死亡率較高(圖2)。由此可見,低濃度草甘膦對京鶴不定芽分化具有顯著的促進作用,隨著濃度升高,不定芽誘導受到阻礙,甚至出現褐化死亡現象。因此,本試驗條件下2.5 mg·L草甘膦最有利于京鶴鱗片誘導不定芽。
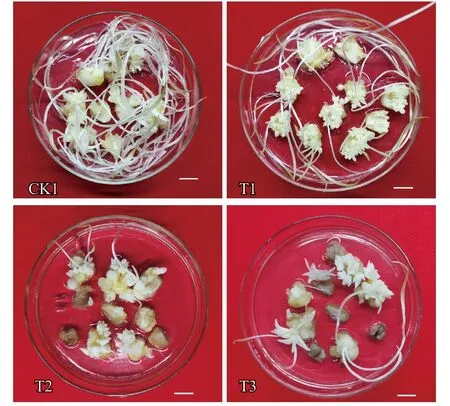
圖2 不同濃度草甘膦對京鶴鱗片不定芽的誘導情況
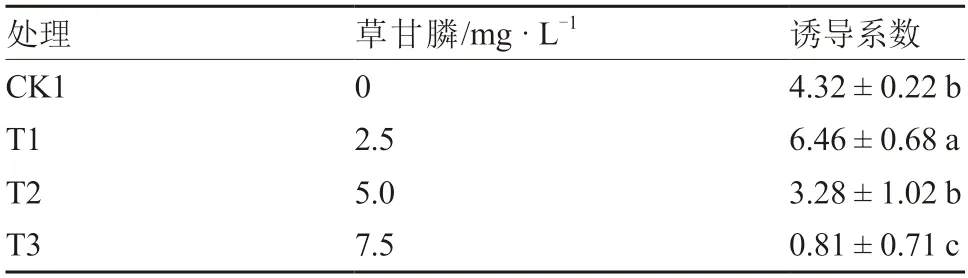
表1 不同濃度草甘膦誘導分化京鶴鱗片不定芽的效果
2.2 6-BA 和NAA 對京鶴不定芽的增殖效果
從表2 可以看出,不同濃度的6-BA 和NAA對京鶴不定芽增殖系數無顯著影響,表明京鶴在不定芽增殖期對6-BA 和NAA 不敏感(圖3)。J1 處理不定芽增殖系數最大,6-BA 與NAA 用量較低,定為本次試驗京鶴鱗片不定芽增殖的最適組合。
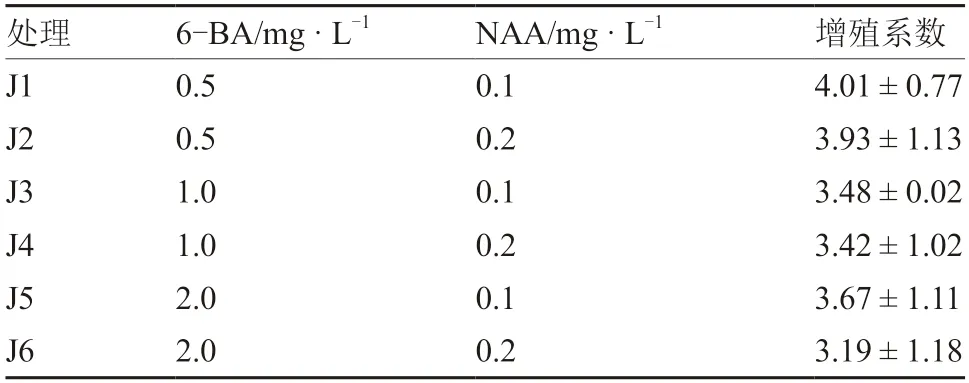
表2 不同濃度6-BA 和NAA 對京鶴不定芽增殖的影響
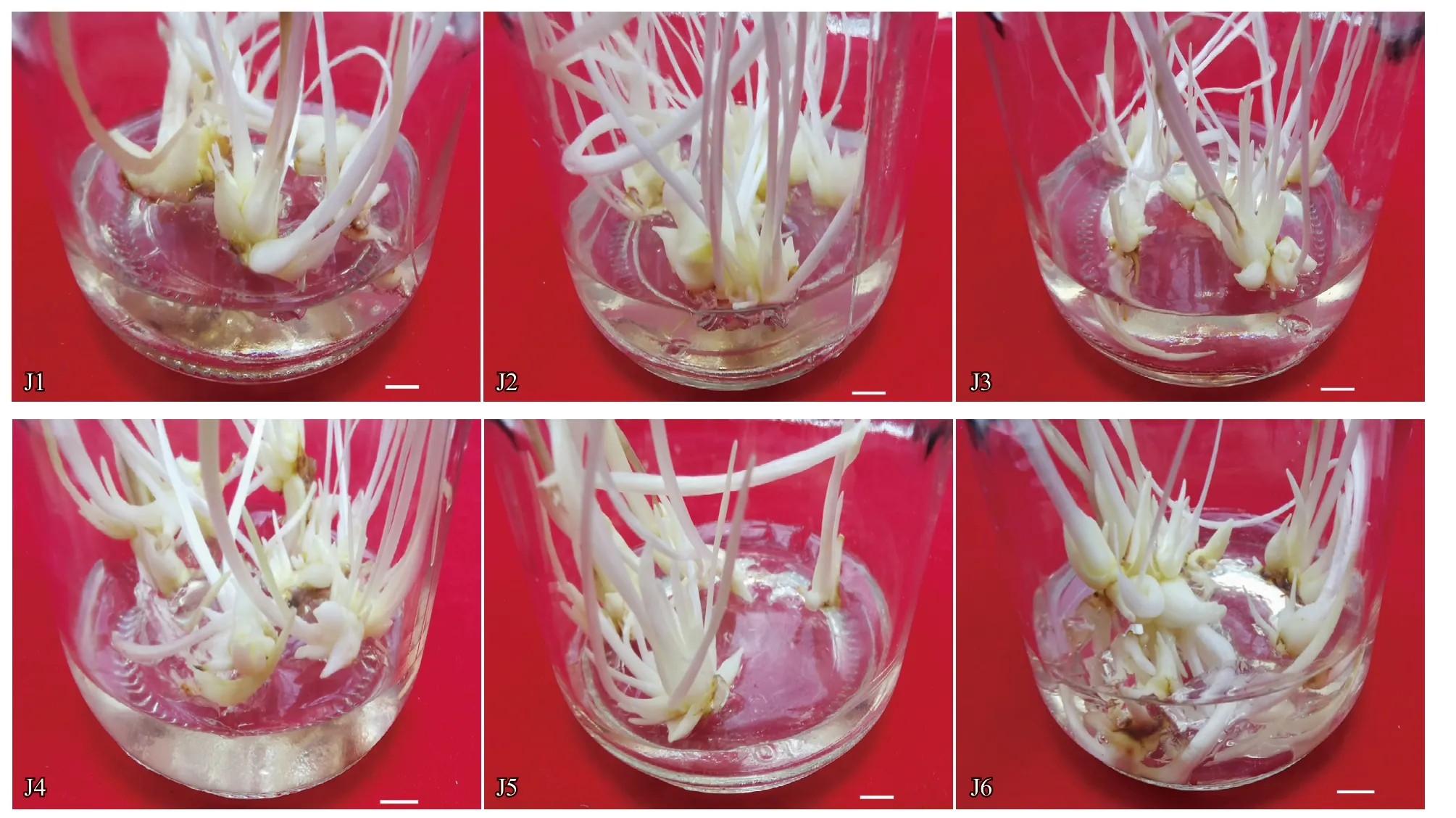
圖3 不同濃度6-BA 和NAA 對京鶴不定芽增殖的影響
2.3 蔗糖對京鶴不定芽鱗莖膨大的影響
從表3 可以看出,在蔗糖濃度為30~90 mg·L時,京鶴不定芽鱗莖直徑隨著蔗糖濃度的提高而增大;當蔗糖濃度為90 mg·L時,平均鱗莖直徑達2.64 cm;而當蔗糖濃度為120 mg·L時,鱗莖直徑不升反降。以上說明,本試驗條件下90 mg·L蔗糖最有利于京鶴不定芽鱗莖的膨大生長。

表3 不同濃度蔗糖對京鶴不定芽鱗莖膨大的影響
2.4 NAA 對京鶴膨大鱗莖生根的影響
從表4 可以看出,G2 和G3 處理的京鶴膨大鱗莖(組培苗)生根系數顯著高于CK2 和G1 處理,其中G3 處理的根也比其他處理更粗壯(圖4)。考慮到組培苗后期移栽種植時,健康、粗壯的根是必要的條件,所以本試驗選擇0.3 mg·LNAA 作為京鶴組培苗生根階段最合適的濃度。

表4 不同濃度NAA 對京鶴膨大鱗莖生根的影響

圖4 不同濃度NAA 對京鶴膨大鱗莖生根的影響
3 討論與結論
3.1 草甘膦在京鶴鱗片不定芽分化中的作用
前人已經建立了多種百合的組織培養再生體系,如金黃花滇百合(張文婷 等,2019)、歐洲百合(張旭紅 等,2018)、蘭州百合(Yang et al.,2021)、毛百合(張艷波,2013)、亞洲百合(楊薇紅 等,2004)、Caesars Palace(李茂娟 等,2019)、東方百合Starfighter(Youssef et al.,2019)、白花百合(Royandazagh,2019;Patil et al.,2021)等,并且在百合不定芽誘導研究中一般使用細胞分裂素與生長素。
本試驗在進行百合鱗片不定芽誘導、分化時,在常規培養組分6-BA、NAA 與蔗糖基礎上,首次使用草甘膦作為誘導分化劑。結果表明,低濃度(2.5 mg·L)草甘膦對京鶴不定芽的誘導分化效果顯著高于常規誘導培養基,每個外植體都能分化不定芽,誘導系數達到6.46,顯著優于前人的試驗結果(楊薇紅 等,2004;張艷波,2013;蘇菁華,2014;Bakhshaie et al.,2016;馬生軍等,2018;Mohebodini et al.,2018;張文婷 等,2019;符勇耀等,2020;Patil et al.,2021);并且,本試驗中使用不同濃度6-BA、NAA 與30 g·L蔗糖的常規誘導結果,也比有些使用細胞分裂素或生長素組合與不同濃度蔗糖的試驗誘導系數高(Sahoo et al.,2018;Lestari et al.,2019;Youssef et al.,2019;Deswiniyanti &Lestari,2020;Filippova et al.,2020)。
3.2 草甘膦在百合鱗片分化中的可能機制及應用
已有研究表明,植物體內存在一種重要的合成芳香族氨基酸的莽草酸途徑,在莽草酸途徑中,磷酸烯醇式丙酮酸(PEP)與莽草酸-3-磷酸(S3P)生成5-烯醇式丙酮酸-3-磷酸莽草酸,這步反應需要5-稀醇丙酮莽草酸-3-磷酸酯合成酶(EPSPS)的催化,才能進行之后的反應并最終形成苯丙氨酸、酪氨酸、色氨酸(Amrhein et al.,1980)。草甘膦可以通過與磷酸烯醇式丙酮酸結合并形成5-烯醇式丙酮酸莽草酸-3-磷酸合酶,競爭性的抑制EPSPS,阻斷苯丙氨酸、酪氨酸和色氨酸的合成來達到殺死雜草的效果(Jaworski,1972;Stephen,2019)。天然植物生長素活性成分IAA 的合成分為依賴色氨酸和非依賴色氨酸兩條途徑,依賴色氨酸的合成途徑又分為吲哚乙醛肟途徑、吲哚丙酮酸途徑、色胺途徑和吲哚乙酰胺途徑,在這些依賴色氨酸的途徑中,色氨酸作為合成IAA 的重要前體物質,經過一系列酶的催化合成吲哚-3-乙醛肟、色胺、吲哚-3-丙酮酸等中間產物,并最終合成IAA,色氨酸的缺失會影響植物體內生長素的合成(王家利 等,2012)。本試驗中,草甘膦的加入可能阻斷了京鶴百合鱗片內部色氨酸的合成,進而使生長素合成受阻,導致內源細胞分裂素和生長素的比值升高,促進了鱗片不定芽的分化,其相關作用機制尚需進一步試驗探究。
草甘膦常用于轉基因材料的抗性篩選,暫時未發現安全問題。草甘膦是否也可以促進其他百合品種不定芽的分化有待進一步研究。
綜上,本試驗建立了草甘膦誘導京鶴鱗片不定芽分化的高效方法,獲得最佳誘導培養基(MS+1.0 mg·L6-BA+0.1 mg·LNAA+2.5 mg·L草甘膦+30 g·L蔗糖),誘導系數達到6.46,顯著高于常規培養基;并以此為基礎,獲得了最佳繼代增殖培養基(MS+0.5 mg·L6-BA+0.1 mg·LNAA+30 g·L蔗糖)、鱗莖膨大培養基(MS+90 g·L蔗糖)以及生根培養基(1/2MS +0.3 mg·LNAA+30 g·L蔗糖),建立了優良百合品種京鶴的高效再生體系,為百合鱗片不定芽誘導、組培苗生產提供了新的思路。
