女貞種子休眠機制和萌發特性研究
2022-08-11 06:34:50燕麗萍趙嘉豪程天利
江蘇農業科學 2022年15期
吳 紅,燕麗萍,夏 群,趙嘉豪,程天利
(1.江蘇農牧科技職業學院園林園藝學院,江蘇泰州 225300;2.山東省林業科學研究院/山東省林木遺傳改良重點實驗室,山東濟南 250000)
具有生活力的成熟種子在適宜的環境條件下也不能立即萌發,必須經歷一個相對靜止的階段,這種現象叫種子的休眠,種子休眠是種子在長期系統發育過程中獲得的一種抵抗外界不適宜環境的適應性,而在農業生產上,種子的休眠嚴重限制了種子生產以及種子的擴繁應用。因此,種子休眠機制以及打破種子休眠方法的研究是種子開發利用的重要保障。近年來,國內外學者已廣泛開展了多種作物種子休眠機制研究,劉洋等對烏蘇里鼠李()種子休眠的特性進行研究發現,該種子為綜合休眠型,是由種皮障礙和內源抑制物質共同導致休眠;張云等通過對膜苞鳶尾()種子的研究發現,膜苞鳶尾種子胚無休眠特性,種皮和胚乳束縛是造成種子休眠的主要原因。另外,國內外學者關于破除種子休眠的方法也開展了大量的研究,如金銀花()種子低溫層積75 d能夠有效解除種子休眠;用刀刺破種皮或用96%濃硫酸浸種能打破西藏砂生槐()種子的休眠,提高發芽率。
女貞()為木樨科(Oleaceae)女貞屬()植物,原產于我國,現廣泛栽植于長江流域及華南、西南地區,向西延伸至陜西、甘肅等地。女貞四季婆娑,樹形整齊,適應性強,是園林中常用的觀賞樹種。從女貞葉中提取的冬青油具清香味,可用作甜食和牙膏的香料;冬青油還具有鎮痛、消炎、抗風濕和祛風等作用,可用來治療肌肉疼痛。林文群等的研究表明,女貞種子含有女貞子酸(ligustrin)、甘露醇(mannitlo)、齊墩果酸(oleanolic acid)、熊果酸(ursolic acid)、樺木醇(betulin)等,有滋養肝腎、強腰膝、烏須明目的功效。國內學者對女貞的栽培技術、抗逆生理和病蟲害防治等方面進行了研究。對種子的研究多集中在播種技術方面,如種子播種品質性狀、種子種苗地理變異及苗期抗寒性的研究、種子化學成分的研究、女貞種子和幼苗對模擬酸雨的反應,而對其種子萌發特性特別是種子體眠機制以及破除種子休眠的方法研究還未見報道。本研究以女貞種子為研究對象,通過對其種子吸水特性、種皮(果皮)的機械阻力等進行研究,探討女貞種子的休眠機制和萌發特性,并通過HSO溶液、NaOH溶液、浸種和低溫層積等處理來研究快速破除女貞種子休眠的方法,以期為女貞的種質資源利用,大規模育苗提供技術參考。
1 材料與方法
1.1 試驗材料
于2020年8月采集江蘇農牧科技職業學院(32°01′57″~33°10′59″E,119°38′24″~120°32′20″N)校內實訓基地內果皮呈現紫色的女貞果實,用手搓洗去除果皮和果肉,自然干燥后挑選飽滿的種子置于牛皮紙袋內存于0~4 ℃冷庫。內源性物質試驗所用青雜三號白菜()種子購自江蘇中江種業股份有限公司。
1.2 試驗方法
1.2.1 種子形態特征和千粒質量測定 采用放大鏡觀察種子外部狀況(形狀、顏色以及種皮和果皮特性);采用精度為0.05 mm的游標卡尺測定種子的縱徑和橫徑;采用感量為0.001的電子天平測定種子的千粒質量,重復8次取其平均值。
1.2.2 女貞種子吸水率的測定 種子吸水特性的測定參照GB 2772—1999《林木種子檢驗規程》中介紹的方法,取上述自然風干的去殼和未去殼的種子各100粒,置于200 mL燒杯內,加入水量沒過種子表面,于25 ℃的恒溫箱浸種。重復3次。定期取出(白天每2 h 1次、夜間每4 h 1次)種子,用濾紙吸干種子表面水分,用感量為0.001 g的電子天平測定種子質量,依次重復至種子質量不再變化為止。
種子吸水率計算公式:=(-)×100%。
式中:為女貞種子的吸水率;為種子浸種后的質量;為種子浸種前的質量。
1.2.3 種子發芽試驗 采用喻梅等介紹的紙上發芽法,在25 ℃光照培養箱中進行,光/暗周期為 12 h/12 h,每皿50粒,3次重復;種子發芽期間逐日記錄,種子露白視為發芽,記錄種子發芽數,當供試種子連續5 d不萌發時視為發芽結束,根據以下公式計算女貞種子發芽率和發芽勢。
發芽率()=()×100%;
發芽勢()=(/)×100%。
式中:為發芽種子數;為供試種子數;為發芽最高峰時發芽種子數。
1.2.4 離體胚培養 隨機取女貞種子200粒,用蒸餾水浸泡24 h后去殼,分4組每組50粒進行離體胚試驗,處理如下:(1)去除全部胚乳,只保留胚;(2)去除子葉端胚乳,露出1/2子葉;(3)去除胚根端胚乳,露出1/2胚根;(4)保留全部的胚乳。每組重復3次,上述材料分別置于潔凈培養皿中,皿中鋪2層潔凈濾紙并用去離子水浸濕,然后將培養皿置于25 ℃光照培養箱內培養,統計各組萌芽情況。
1.2.5 內源性物質提取及白菜籽發芽的測定 隨機取新鮮女貞種子,剝取木質化的內果皮和胚乳,用感量為0.001 g的電子天平稱取上述材料各5 g,置于預冷的研缽中研碎,用80%甲醇溶液轉移于 25 mL 試管中,后置于4 ℃冰箱中浸提48 h,期間不斷振蕩以保證提取充分,然后離心取上清液,重復以上操作并將上清液混合置于50 ℃烘箱內濃縮蒸干并用蒸餾水定容至50 mL備用。分別取定容后的浸提液1、2、3、4、5 mL用蒸餾水定容至5 mL配制成20%、40%、60%、80%、100% 的溶液,浸泡白菜種子,將浸泡種子置于25 ℃光照培養箱中培養,測定白菜種子的發芽率,每個處理重復3次,以蒸餾水浸泡為對照。
1.2.6 種子解除休眠方法研究
1.2.6.1 HSO、NaOH溶液處理對女貞種子萌發特性的影響 女貞種子用體積分數為98% 的HSO溶液浸種處理10、20、30、40、50 min;或者用0.3 mol/L的NaOH溶液浸種6、12、18、24、36 h,浸種后的種子用蒸餾水沖洗3次,以未浸種的種子為對照,參照“1.2.3”節中的方法測定種子的發芽率和發芽勢,每個處理50粒種子,3次重復。
1.2.6.2 浸種水溫和時間對女貞種子萌發特性的影響 用20、35、50、65 ℃的自來水分別處理女貞種子4、6、8、10、12、16 h,以未浸泡的種子為對照,處理結束后參照“1.2.3”節中的方法測定種子的發芽率,每個處理50粒種子,3次重復。
1.2.6.3 低溫層積處理對女貞種子萌發特性的影響 將沙(最大持水量50%)與女貞種子以3∶1的比例(體積比)埋藏于塑料花盆中,后置于4 ℃條件下保存。每隔3 d噴水并翻動以保證塑料花盆內的濕度和通氣環境。層積30、50、70、90 d,以未層積處理的種子為對照。處理結束后參照“1.2.3”節中的方法測定種子的發芽率,每個處理50粒種子,3次重復。
1.3 數據處理
試驗數據采用Microsoft Excel及DPS v7.05進行數據分析和作圖,各處理采用最小顯著性差異()法對數據進行差異性分析。
2 結果與分析
2.1 種子形態特征和千粒質量測定結果
女貞種子為腎形,兩端尖細,成熟時為黃褐色至棕褐色,千粒質量為32.17 g,種子縱橫徑比為2.19;種子由種皮、胚乳和胚3個部分組成,成熟的種子外被一層木質化的內果皮,種皮褐色膜質化,種皮內為厚而堅硬的胚乳,是種子的重要組成部分,胚包埋于胚乳內,體積小,子葉發達且下胚軸長(圖1)。
2.2 女貞種子的吸水率變化
女貞種子吸水率變化情況如圖2所示。隨著女貞種子吸水時間的延長,吸水速率逐漸增加,去殼和未去殼種子的吸水率曲線變化一致,但去殼種子的吸水率大于未去殼種子。由圖2可知,女貞種子物理吸水過程包括3個時期:0~8 h為種子快速吸水期,吸水量大而迅速,8 h時去殼和未去殼種子吸水率分別達到63.89%、37.90%;8~28 h為緩慢吸水期,種子吸水緩慢但歷時長,在28 h時去殼和未去殼種子的吸水率分別為77.46%、57.30%;28~48 h為飽和吸水期,吸水量逐漸趨于平衡,在飽和吸水末期,去殼種子已完成種子總吸水量的84.09%,幾近飽和,未去殼種子吸水率已達到63.40%,表明女貞種子果皮具有一定的透水性。


2.3 種殼對女貞種子萌發的影響
造成種子休眠的原因之一是機械壓迫,即種子外堅硬的種皮(果皮)或胚乳緊緊包裹種胚,以至于胚在萌發時胚芽或胚根生長力不足以穿透其束縛而造成休眠,因此種子萌發與否取決于種胚的穿透力和機械壓迫力之間的比值。
女貞種子離體培養結果表明,剝去全部胚乳的離體胚置床培養1周后胚根端先變綠生長,2周后子葉開始生長,萌發率達到99%;露出1/2胚根的處理在前期胚根開始膨大,第2周胚根彎曲生長,而包被在胚乳中的子葉卻無生長現象,后期胚根停止生長,種子霉爛;露出1/2子葉的離體胚培養1周后子葉開始變綠,2周后子葉展開,但直至第4周胚根仍無生長跡象;保留完整胚乳的種胚置床1周后均無胚根或子葉露出,后期陸續發霉腐爛。這說明女貞種胚不具備休眠性,去掉種殼后的種子不能順利萌發,堅硬的胚乳影響了水和氧氣的透性,因此,胚乳可能是限制女貞種子萌發的原因之一。
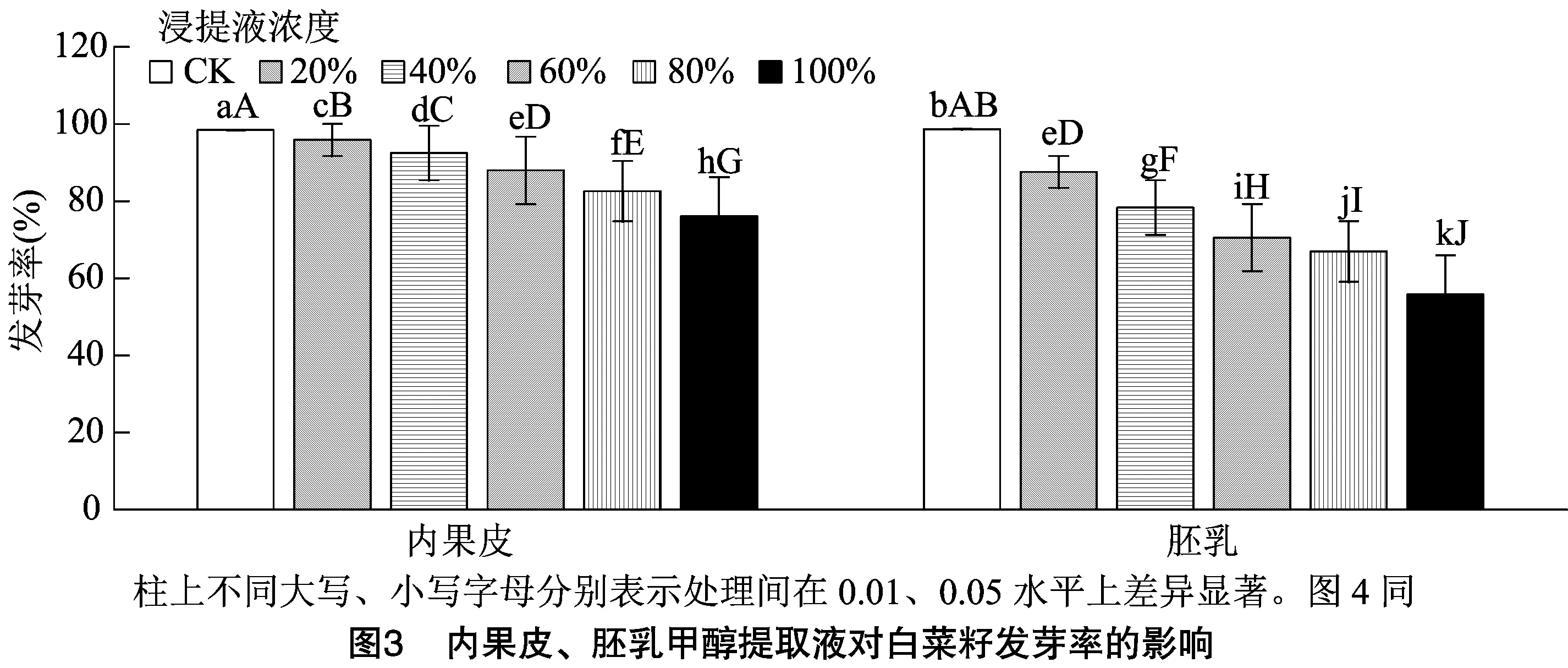
2.4 女貞種子內果皮、胚乳浸提液對白菜籽萌發率的影響
種子內含有內源抑制物質是導致種子休眠的一個重要原因,種子不同部位的浸提液對白菜籽發芽率影響的研究是測定內源抑制物質的快速而行之有效的方法。女貞種子內果皮、胚乳甲醇浸提液對白菜籽發芽率的影響如圖3所示,女貞種子內果皮和胚乳浸提液在一定程度上抑制了白菜種子的萌發,且不同部位浸提液處理白菜種子的發芽率隨著浸提液濃度的增大而降低,不同濃度的內果皮甲醇提取液處理的白菜籽發芽率分別比對照降低了2.75%、6.05%、9.83%、16.19%、21.97%;不同濃度的胚乳甲醇提取液處理的白菜籽發芽率分別比對照降低了9.64%、19.07%、26.69%、30.43%、42.65%,各處理與對照之間均差異極顯著(<0.01)。從不同部位浸提液對白菜籽發芽率的影響來看胚乳的抑制效果大于內果皮,方差分析結果表明,除60%內果皮浸提液和20%胚乳浸提液差異不顯著外,其他各處理間差異均極顯著(<0.01)。
2.5 不同處理對女貞種子萌發的影響
2.5.1 HSO、NaOH溶液處理 由表1可以看出,經體積分數為98%的HSO溶液浸種處理10、20、30、40、50 min 的發芽率分別為35.30%、60.49%、88.59%、71.19%、67.48%,均高于對照,且各處理間差異極顯著(<0.01);在10~30 min范圍內隨著處理時間的延長,發芽率呈上升趨勢;從30~50 min 范圍內隨著處理時間的延長,發芽率呈下降趨勢。由表1還可以看出,女貞種子的發芽勢與發芽率變化趨勢一致,表明在30 min時女貞種子的發芽率高而且發芽集中(發芽勢最高),因此,女貞種子用濃硫酸浸種處理的最佳時間為 30 min 左右。

表1 H2SO4溶液處理對女貞種子發芽特性的影響
NaOH溶液處理結果(表2)顯示,0.3 mol/L NaOH溶液處理女貞種子,在6~24 h范圍內種子的發芽率和發芽勢均隨處理時間的增加而不斷增加,各處理間差異達極顯著水平(<0.01);處理24 h后,種子的發芽率和發芽勢分別達85.07%、82.48%,處理效果最好;處理36 h后種子的發芽率和發芽勢分別達到了84.10%、74.27%,處理效果僅次于處理24 h,且二者之間的發芽率差異不顯著,但二者之間的發芽勢差異極顯著(<0.01)。究其原因在于,短時間的NaOH溶液處理時,其腐蝕性可軟化內果皮,提高種子的透水性和透氣性,促進種子萌發;而長時間處理時種子內的蛋白質活性降低,種子萌發受到抑制。

表2 NaOH溶液處理對女貞種子發芽特性的影響
2.5.2 浸種處理 浸種對女貞種子發芽率的影響結果(圖4)表明,在浸泡4~10 h種子的發芽率隨水溫的增加呈先升后降的趨勢,大體表現為在20~50 ℃范圍內,隨著浸種水溫的增加,種子的發芽率逐漸上升,在50 ℃時達到最高值;12~16 h隨著水溫的增加種子的發芽率逐漸降低,且各溫度處理間差異極顯著(<0.01)。女貞果實為核果,內果皮堅硬致密,未浸泡處理的女貞種子的發芽率僅為32.33%,經浸種處理的種子的發芽率均高于CK。因此,水溫在20~50 ℃范圍內浸種女貞種子的發芽率最高。不同浸種時間種子發芽率亦有不同,20 ℃浸泡12 h時發芽率最高,達84.67%;其次是50 ℃浸泡 8 h 和35 ℃浸泡10 h處理,其發芽率分別為83.33%、82.67%,二者無顯著差異。65 ℃浸泡12、16 h處理的發芽率較低,均低于50%,分別為48.67%和45.44%;65 ℃浸泡10 h和50 ℃浸泡 16 h 處理的種子發芽率均低于60%,分別為51.33%、53.72%。
女貞種子的發芽率受水溫和浸泡時間的雙重影響,在適宜的浸種溫度下,隨著水溫的增加,浸種時間會相應縮短,可見浸種處理可以軟化內果皮,去除種子表面蠟質、提高透性并浸出種子內的抑制物質,從而提高種子萌發率。因此本試驗的結果表明,女貞密實的內果皮是阻礙種子萌發的重要因素。

2.5.3 低溫層積對種子萌發的影響 由圖5可知,低溫層積時間的長短對女貞種子發芽率有很大的影響。在4 ℃處理條件下,種子累計發芽率隨著層積天數的增加先升高后下降,層積50 d種子的發芽率最高,種子累計發芽率達85.88%,比對照提高20.66%,層積效果最好,其后依次為層積30 d、層積70 d、CK、層積90 d,各溫度處理間差異極顯著(<0.01)。層積處理90 d時累計發芽率僅為64.91%,發芽率極顯著(<0.01)低于對照,比對照提高8.56%。這一變化規律與蔡丹紅等的研究結果一致。
3 討論與結論
種子休眠指具有生活力的種子停留在不能發芽的狀態的一種現象,種子休眠是植物形成的抵御不良環境的一種適應能力,種胚引起的體眠可分為胚的影響、種皮(果皮)影響和抑制物質影響。
有些種子外部形態發育成熟且已具有成熟種子的特征,但生理上必須經過一定階段的發育,才能完成其內部的生理生化過程。如李鐵華等發現引起鐘萼木()種子休眠的主要原因是種子內缺乏萌發促進物質和酶活性低,用赤霉素溶液浸種或層積可解除種子休眠;劉繼生等的研究表明,人參()種子成熟時其胚未發育完全,層積初期需提高溫度促進胚的生長與分化,層積后期需降低溫度促進種胚的生理生化過程。本試驗中女貞種子種胚形態結構發育完全,離體胚培養結果表明,女貞種子種胚在去除種皮和胚乳后能在25 ℃恒溫箱內順利萌發,可見女貞種胚不存在休眠。

種胚外面包裹堅硬致密的包裹物質會導致種子的物理體眠,這些包裹物質可能是種皮(果皮)或胚乳,有些種子種皮(果皮)表面含有蠟質、黏質或革質物質,影響了種胚的透水性和透氣性,進而影響種子內有氧代謝和種胚的營養供給,并對種胚胚根和胚芽的生長造成機械約束,有些硬實種子則是堅硬的種皮(果皮)對種胚胚根和胚芽的生長造成機械約束。如馮毓琴等發現,天藍苜蓿()種子成熟時胚已發育成熟,硬實種子存在種皮抑制現象,導致種子很難吸水膨脹且種胚伸長受阻;堅硬致密的胚乳也是影響種胚生長的因素之一,王宏迪研究發現,黃精()屬于綜合休眠,其密實堅硬的胚乳細胞阻礙水分和空氣進入種胚內部而導致休眠。通過女貞透水性試驗和離體胚試驗可知,女貞未去殼種子吸水率低于去殼種子,但在飽和吸水期未去殼種子的吸水量達63.40%,表明女貞內果皮雖相對堅硬致密,可能對種子萌發具有一定的影響但這不是女貞種子休眠的主要原因,本研究發現20 ℃浸泡12 h、50 ℃浸泡8 h和35 ℃浸泡10 h均可打破種子休眠,提高女貞種子的發芽率。有研究表明,HSO和NaOH溶液處理可腐蝕內果皮,打破胚乳的屏障,提高種子的透氣性,如杜小芳等發現,甘草()、苦豆子()和苦馬豆()種子屬種皮障礙引起的物理性休眠,磨破、切割種皮以及利用濃硫酸破壞種皮致密組織等措施均能減弱機械障礙,促進種子萌發。本研究用體積分數為98%的HSO溶液浸種處理10、20、30、40、50 min的發芽率與對照相比均顯著提高,其中30 min處理的發芽率最高;0.3 mol/L NaOH溶液處理也可顯著提高女貞種子的發芽率,24 h處理效果最好,表明女貞種子堅硬的內果皮和胚乳的機械束縛是阻礙種子萌發的根本性限制因子。
種子或果實內存在的內源抑制物也會導致種子休眠,如禾本科(Gramineae)、樟科(Lauraceae)、蘭科(Orchidaceae)、木蘭科(Magnoliaceae)和瑞香科(Thymelaeace)植物等都屬這一類。本研究中女貞種子不同部位內源性抑制物的提取液對白菜籽發芽抑制測定結果表明,女貞種子內果皮和胚乳中存在發芽抑制物質,說明種子內含有抑制物質可能是導致女貞種子休眠的另一個主要原因。關于這一猜測目前尚未見報道,有必要深入展開相關研究。
