2株分泌型鐵載體真菌對番茄青枯病的防效
2022-08-08 03:41:40趙江源鄒雪峰楊濟達朱紅業楊佩文李銘剛
植物保護 2022年4期
趙江源, 鄒雪峰, 何 翔, 張 慶, 楊濟達,朱紅業, 楊佩文*, 李銘剛
(1. 云南大學云南省微生物研究所, 昆明 650091; 2. 云南農業大學植物保護學院, 昆明 650201;3. 云南省農業科學院農業環境資源研究所, 昆明 650205)
云南省哀牢山國家級自然保護區地處云南橫斷山脈縱谷地帶。東亞季風氣候、青藏高原季風氣候和南亞東南亞熱帶季風氣候均在此匯集,形成特殊的地形-氣候-水文特征耦合。該地區土壤有機質含量豐富,蘊藏著豐富的種質資源[1]。從前期我們對土壤微生物分離、篩選和拮抗試驗結果看,該區域原始森林土壤環境中存在豐富的鐵載體微生物資源,值得進行深入發掘與利用[2]。
鐵元素對微生物生長、發育及代謝過程有重要影響[3-4]。自然界中鐵元素主要以不溶解三價態穩定形式存在,為獲取鐵元素,微生物通常會分泌對鐵離子(Fe3+)有螯合能力的鐵載體(siderophore)來實現這一過程[5]。微生物鐵載體主要類型包括異羥肟酸鹽型(hydroxamates)、兒茶酚鹽型(catecholates)及羧酸鹽型(carboxylates)[6]。
目前,植物病害如番茄青枯病[7]、馬鈴薯青枯病[8]、煙草青枯病[9]的暴發、主要是青枯雷爾氏菌Ralstoniasolanacearum大量繁殖擴增所導致[10-11],而這其中環境鐵元素的分配可能是一個重要原因[12]。正因為如此,如何利用自然界中非病原菌所產鐵載體,通過干擾病原菌對鐵元素的正常攝取來抑制植物病原微生物生長已成為近年來的研究熱點[13]。比如,Solanki等[14]發現,具有鐵載體活性的根瘤菌株對水稻紋枯病菌Rhizoctoniasolani抑制活性較強,而不產鐵載體菌株的拮抗活性較弱。Dutta等[15]對生防菌進行篩選時,發現防效較強的菌株普遍具有分泌兒茶酚鹽型鐵載體的能力。Ambrosi等[16]發現,熒光假單胞菌Pseudomonasfluorescens代謝產物中存在大量的羧酸鹽型鐵載體化合物;其通過分泌一系列鐵載體物質來抑制土壤環境中植物病原菌的生長。Fgaier 等[17]基于微生物鐵載體鐵螯合原理,建立鐵載體介入下的微生物群落模型,得出產生鐵載體的菌種比其他菌種更具競爭優勢的結論,并以此來調控植物病原微生物群落結構和組成[18]。
目前有關鐵載體對植物病原菌抑菌活性的研究還相對較少,本研究以分離自云南省哀牢山原始森林土壤中高活性鐵載體菌株為材料,分析其鐵載體結構類型及其對特定植物病原微生物的拮抗效果和對植物病害的防治效果,可擴充植物病害拮抗菌(劑)范圍,揭示鐵載體在植物病害防控中的應用前景,為鐵載體拮抗菌劑(制劑)的研發、相關作用機制的深入研究奠定基礎。
1 材料與方法
1.1 供試菌株
云南木霉Trichodermayunnanense(編號2-14F2)與擬球孢白僵菌Beauveriapseudobassiana(編號2-8F2)分離自云南省哀牢山國家自然保護區原始森林土壤,菌株保藏于中國普通微生物菌種保藏管理中心,保藏編號為CGMCC No.17926與CGMCC No. 21047。青枯雷爾氏菌Ralstoniasolanacearum分離自番茄青枯病病株。供試真菌均在20%甘油中保存于-80℃。試驗前先將相關菌株接種在PDA平板上活化至少2代,使其恢復旺盛生長。
1.2 供試溶液及培養基的制備
CAS顯色液配制:取0.060 5 g鉻天青S (chromeazurol S, CAS)溶于50 mL去離子水中,加入0.002 7 g FeCl3攪拌混勻,標記為A液;再稱取0.072 9 g十六烷基三甲基溴化銨溶于40 mL去離子水,標記為B液;最后將A液緩慢倒入B液中,攪拌均勻即可。
指示菌懸液配制:青枯雷爾氏菌斜面試管中加入2 mL無菌水,刮取菌苔,之后用無菌水進行梯度稀釋(用酶標儀或分光光度計檢測OD625讀數),取OD625=0.1菌液配制菌懸液(濃度為1×108cfu/mL)。
PDA培養基:馬鈴薯200 g/L、葡萄糖20 g/L、瓊脂15~20 g、去離子水1.0 L、pH(7.0±0.1);無鐵查氏液體培養基:葡萄糖30.0 g/L、硝酸鈉2.0 g/L、三水合磷酸鉀1.0 g/L、氯化鉀0.5 g/L、七水合硫酸鎂0.5 g/L、8-羥基喹啉0.75 g/L、去離子水1.0 L、pH(7.0±0.1);NA培養基:蛋白胨10.0 g/L、牛肉粉3.0 g/L、氯化鈉5.0 g/L、瓊脂15.0 g/L、pH(7.3±0.1);雙層顯色培養基:底層為5 mL的水瓊脂培養基(加2 mL CAS液),上層為5 mL的PDA培養基。
1.3 菌株產鐵載體活性測定
用打孔器取菌餅(直徑0.5 cm),接種在雙層顯色培養基的中心上層,每菌株3個重復。將完成接種的培養基放置在(27±1)℃恒溫培養箱中培養7 d,觀察顯色層顏色變化。
挑選培養基底層顏色變紅的菌株,加入到無鐵查氏液體培養基中,在28℃ 150 r/min的恒溫搖床(型號:HS-200B)上培養 48 h,培養結束后吸取2~5 mL培養液用0.22 μm無菌濾膜過濾后加入等體積的CAS檢測液,靜置1 h后用全波長酶標儀(型號:Multiska GO)測定其 OD630(記作“As”),用相同方法測定未接菌的液體培養基的 OD630作為參比值(記作“Ar”)。鐵載體的濃度用鐵載體活性單位(siderophore unit, SU)表示,SU=[(Ar-As)/Ar]×100%,測定重復3次,取平均值進行比較分析。
1.4 鐵載體活性物質分離
將25 g蒸熟大米裝入到500 mL組培瓶中,121℃高壓蒸汽滅菌30 min,然后加入1 mL 2-8F2或 2-14F2菌懸液,充分攪拌,放入(37±1)℃恒溫培養箱內靜置培養30 d。完成發酵后在發酵瓶中加入95%甲醇充分浸泡24 h,用5 mm孔徑濾紙過濾。旋轉蒸發儀(R-210 BUCHI)溫度設置45℃,轉數700 r/min,將濾液中有機溶劑完全蒸發。
通過柱層析色譜法分離活性物質。先用正相硅膠 G(200~300目,青島海洋化學工業制品廠)進行初步劃段分離,采用Pan等[20]的方法利用流動相梯度恒速洗脫。經正相硅膠分離的餾分選用Sephadex-LH20凝膠(GE,美國)繼續純化,流動相為甲醇,流速為10 mL/30 min,用自動收集儀收集餾分,每個餾分10 mL,采用CAS液與2% FeCl3檢測各餾分顯色情況,其中與CAS液反應顯紅色,同時與2% FeCl3反應顯褐色或棕褐色的餾分為活性餾分。根據顯色情況合并濃縮活性餾分備用。
獲得的鐵載體活性餾分減壓蒸干后置于冰箱4℃保藏,使用時用去離子水溶解,配制成不同濃度的鐵載體溶液。
1.5 鐵載體類型判定
1 mL鐵載體活性物質中加入1 mL 2% FeCl3溶液,出現紅色或紫色表明測試樣品中含有鐵載體。用紫外分光光度計檢測(EU-2200R),在 420~450 nm 之間出現吸收峰則說明螯合物質為異羥肟酸鹽型鐵載體。
1.6 鐵載體活性物質抑菌活性測定
用雙層平板培養基測定鐵載體活性物質對青枯雷爾氏菌的抑制效果。將圓形濾紙片(直徑0.5 cm)在抑菌待測液(0.25 mg/mL的鐵載體溶液)中浸泡5 min,備用。在融化的NA培養基(約55℃)中加入400 μL青枯雷爾氏菌菌懸液(1×109cfu/mL),充分搖勻后倒在水瓊脂平板上層,待培養基凝固后接入浸泡待測液的濾紙片和未處理的濾紙片(空白對照),濾紙片以培養皿圓心為中心對稱放置。每處理3次重復(3個平板)。25℃培養72 h后記錄抑菌圈面積。
使用96孔板測定鐵載體活性物質對青枯雷爾氏菌的抑制率。板孔中加入100 μL青枯雷爾氏菌菌懸液(濃度為1×108cfu/mL)與100 μL NA液體培養基,之后在96孔板A/B 兩排的第一孔中,分別加入100 μL的缺鐵(未加FeCl3溶液)與富鐵(含1 mol/L FeCl3)的鐵載體活性物質溶液,從第一孔開始進行倍半稀釋。每孔對應鐵載體活性提取物的濃度分別為0.60、0.30、0.15、0.075、0.038、0.0187 5、0.009 mg/mL……以此類推。每處理3個重復(96孔板中3行)。將96孔板放入37℃恒溫培養箱16~20 h后,用酶標儀測定 625 nm 處的吸光度,換算成麥氏比濁濃度或微生物的近似濃度,計算殺菌(抑菌)率。計算公式為:抑制率=[對照組微生物濃度(或麥氏比濁濃度)-抑菌待測組微生物濃度(或麥氏比濁濃度)]/對照組微生物濃度(或麥氏比濁濃度)×100%。對照組未添加任何抑菌物質。
1.7 鐵載體活性物質抑制番茄青枯病盆栽驗證
設置對照組1(CKA):蒸餾水405 mL;對照組2(CKB):蒸餾水400 mL+青枯雷爾氏菌懸液5 mL;處理組1:0.45、0.15 mg/mL和0.05 mg/mL 2-8F2菌株鐵載體活性物質處理, 記為2-8F2A、2-8F2B和2-8F2C;處理組2:0.45、0.15 mg/mL和0.05 mg/mL 2-14F2菌株鐵載體活性物質處理,記為2-14F2A、2-14F2B和2-14F2C。處理組1、2均為蒸餾水350 mL+各濃度的鐵載體活性物質50 mL+青枯雷爾氏菌懸液5 mL。另設置不同濃度的活性物質中添加50 mL FeCl3溶液使其終濃度為1 mol/L 的處理。每處理3次重復。將不同處理的溶液對5株幼苗期的番茄植株進行灌根,然后置于溫度(24±1)℃、濕度(85±5)%的恒溫恒濕光照人工氣候培養箱(MGC-450HP,L∥D=12 h∥12 h),培養30 d,統計植株發病情況。
病害調查采用番茄青枯病分級標準[19]。0級,葉片無癥狀;1級,植株1/4以下的葉片表現萎蔫癥狀;2級,植株1/4~1/2葉片表現萎蔫癥狀;3級,植株1/2以上葉片表現萎蔫癥狀;4級,全株萎蔫死亡。病情指數=∑(各級病株數×相應級值)/(調查總株數×最大級值)×100。防治效果=(空白對照病情指數—處理病情指數)/空白對照病情指數×100%。數據采用SPSS進行方差分析,R語言作圖。
2 結果與分析
2.1 產鐵載體活性菌株菌落形態及所產鐵載體體外活性
本研究中兩株具有高鐵載體活性的真菌編號分別為2-14F2與2-8F2,其菌落形態見圖1(a及b)。兩菌株在CAS雙層顯色培養基上培養后,均可產生明顯的紅色暈圈(圖1c和1d),表示2個菌株均能產生螯合鐵離子的鐵載體。通過對兩菌株產鐵載體活力值定量分析,得出2-14F2與2-8F2菌株產鐵載體活性單位(SU)分別為62.02%和52.06%。
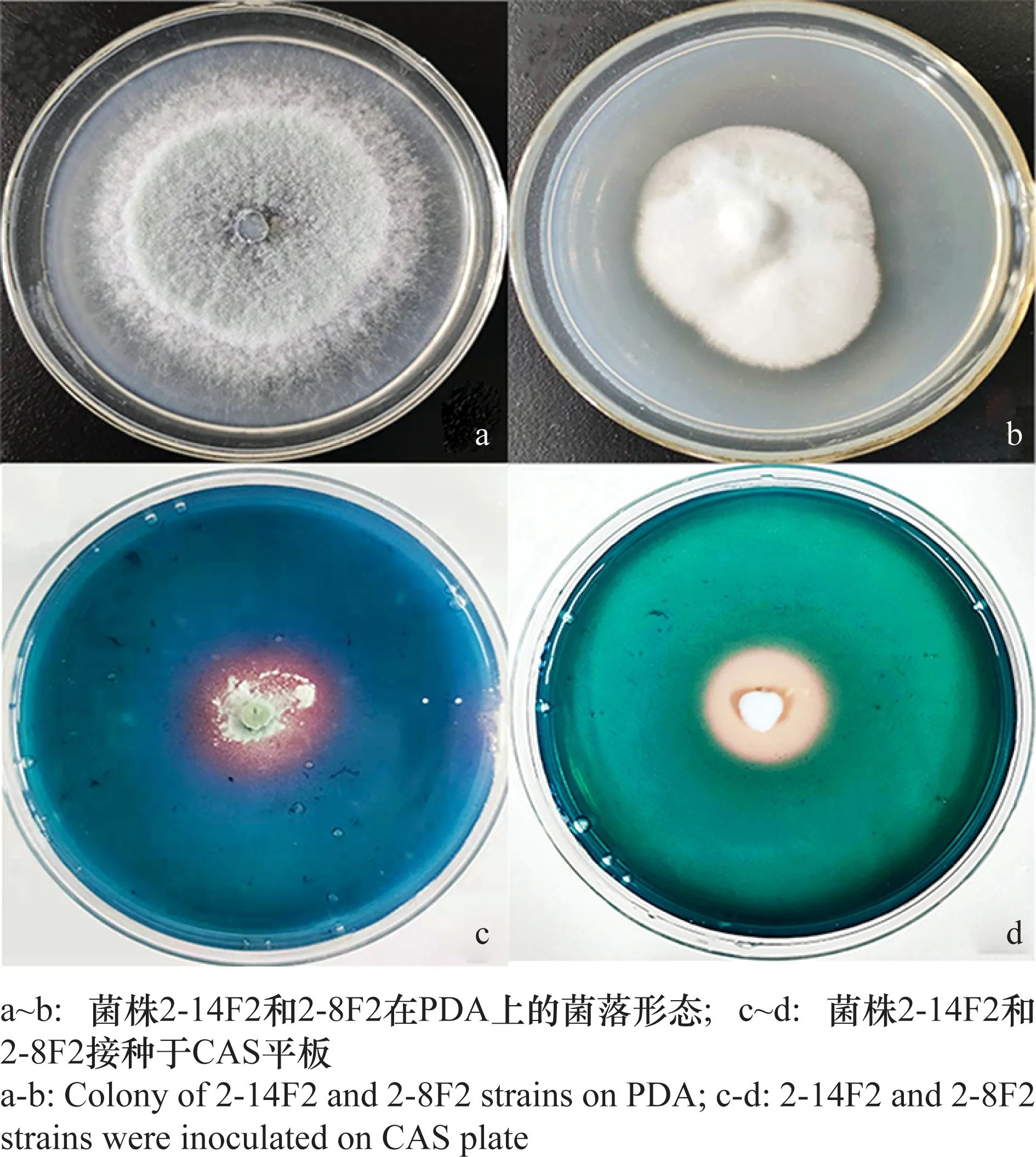
圖1 菌株2-14F2與2-8F2 在PDA和CAS平板上的培養形態Fig.1 The culture morphology of 2-14F2 and 2-8F2 strains on PDA and CAS plates
2.2 鐵載體活性餾分分離
對2-14F2和2-8F2真菌固體發酵粗提物反復經正相硅膠、C18反相硅膠分離及凝膠層析系統純化后(圖2),真菌2-14F2鐵載體活性物質(CAS 顯色為紅色,同時與 2% FeCl3反應顯褐色或棕褐色的餾分)集中在3、5餾分(圖2c),2-8F2鐵載體活性物質集中在2、3餾分(圖2d)。

圖2 菌株 2-14F2和2-8F2分泌的鐵載體活性餾分分離Fig.2 Separation of siderophore active fractions from 2-14F2 and 2-8F2 strains
2.3 活性部位鐵載體類型判定
通過紫外分光光度計(EU-2200R)全波段掃描后發現,2-8F2與2-14F2菌株鐵載體活性物質分別在423 nm和421 nm處吸光度值最高(圖3),故判定兩菌株所產鐵載體類型為異羥肟酸鹽型(hydroxamates)。
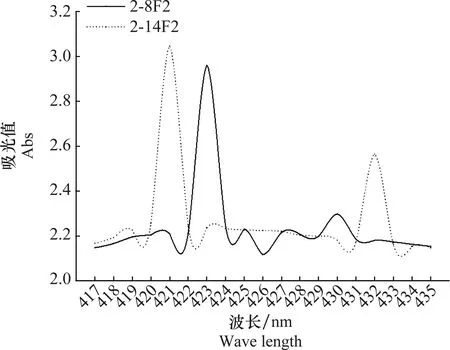
圖3 2-8F2和2-14F2菌株產鐵載體在全波段下吸光度值Fig.3 Full-band absorbance values of 2-8F2 and 2-14F2 siderophores
2.4 鐵載體活性物質抑菌活性測定
紙片擴散可快速判斷鐵載體活性物質對青枯雷爾氏菌的抑制效果。與空白對照(CK)相比,2-14F2與2-8F2菌株鐵載體活性物質表現出明顯的抑菌效果(圖4)。
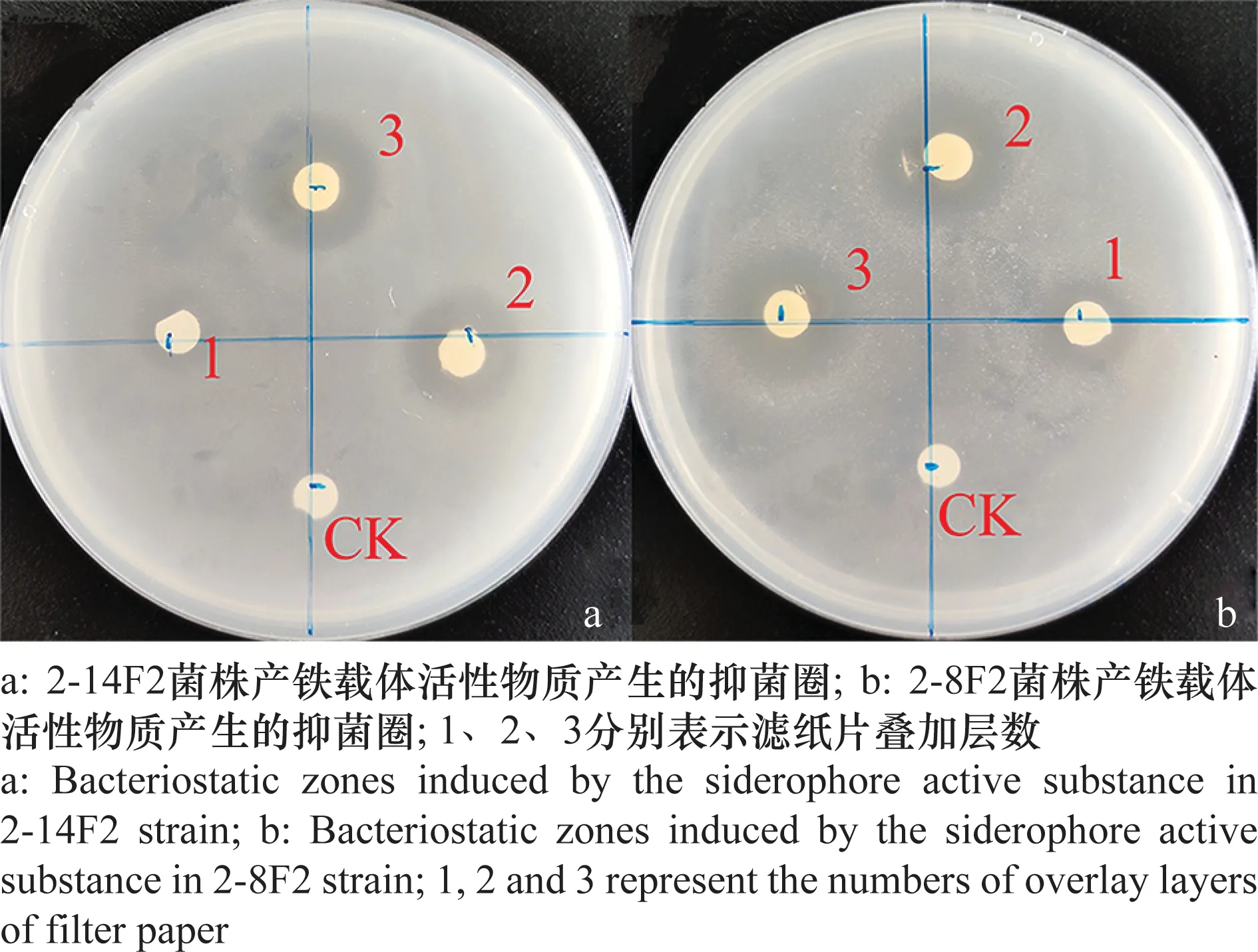
圖4 2-8F2與2-14F2菌株產鐵載體活性物質抑菌定性測定Fig.4 Qualitative determination of the antibacterial activity of siderophores in strains 2-8F2 and 2-14F2
2-8F2與2-14F2的抑菌率隨著鐵載體活性物質濃度下降而降低。此外,當2-8F2菌株與2-14F2菌株鐵載體活性物質中添加FeCl3后,其抑菌率與未加FeCl3樣品相比,抑菌率在高濃度范圍內明顯下降(圖5)。表明抑菌效果與鐵載體所處的環境鐵離子濃度相關。
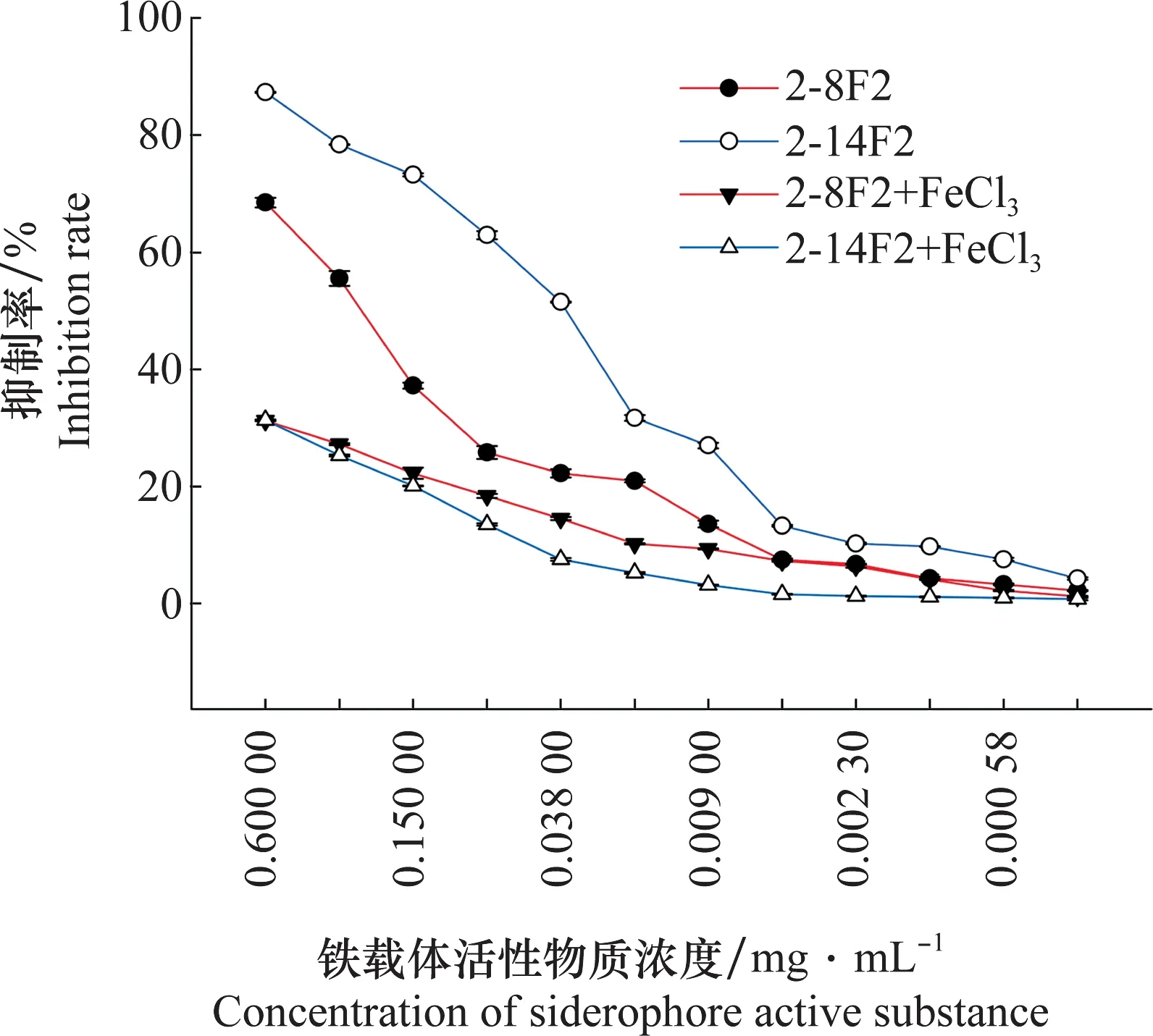
圖5 2-8F2與2-14F2菌株鐵載體活性物質對青枯雷爾氏菌的抑菌率Fig.5 Detection of the antibacterial rate of siderophore active substances from strains 2-8F2 and 2-14F2 against Ralstonia solanacearum
2.5 盆栽試驗驗證鐵載體活性物質對番茄青枯病的防治效果
番茄植株接種青枯雷爾氏菌15 d后,未接種青枯菌的CKA對照組植株未出現萎蔫的癥狀,發病率為0。接種青枯菌的CKB對照組,植株均出現莖基部萎蔫的癥狀,病情指數可達97.31。與CKB處理組相比,2-14F2與2-8F2(A、B和C)處理組的發病率和病情指數明顯降低。表1所示,當鐵載體活性物質濃度為0.15 mg/mL時,防治效果為51.36%(2-14F2B)與50.27%(2-8F2B),此外當添加等體積的1 mol/mL FeCl3溶液后防治效果下降40.55百分點(2-8F2)和41.09百分點(2-14F2)。該結果再次表明:2-14F2和2-8F2菌株的鐵載體活性物質對番茄青枯病具有較好的防治效果(圖6),且抑菌效應與鐵載體所處的環境鐵離子濃度相關。
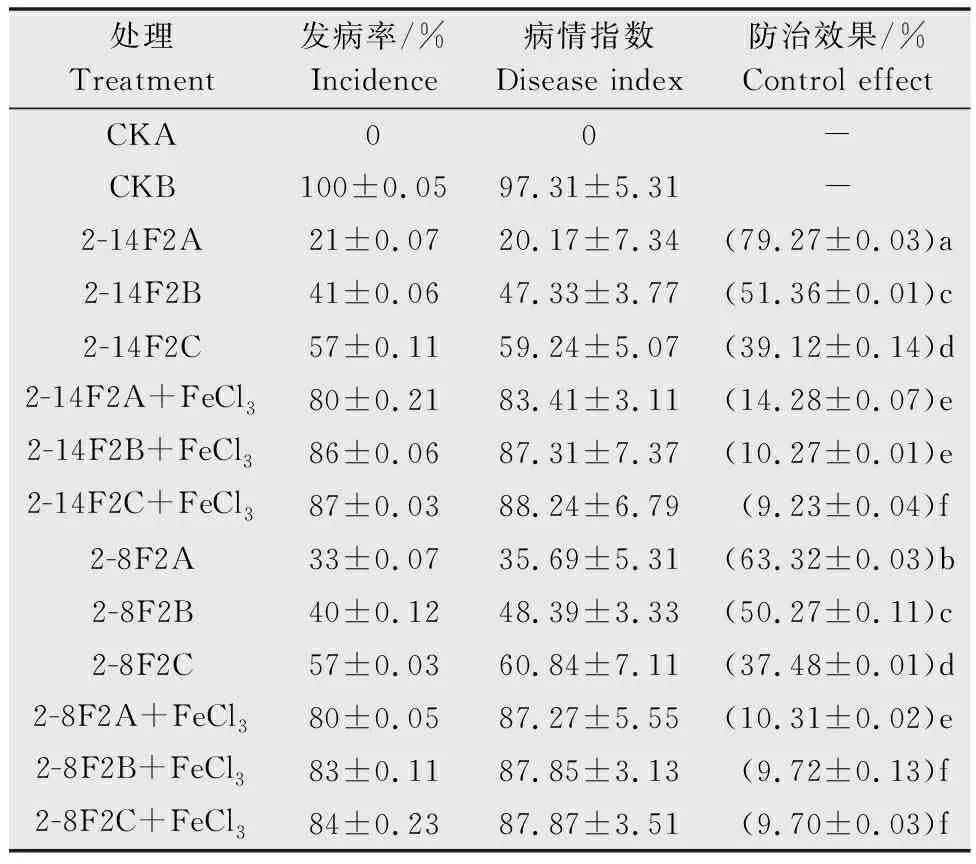
表1 2-8F2與2-14F2菌株鐵載體活性物質對番茄青枯病的防治結果1)Table 1 Control results of 2-8F2 and 2-14F2 siderophore active substances against tomato bacterial wilt

圖6 2-8F2與2-14F2鐵載體活性物質對番茄青枯病的抑制效果Fig.6 Inhibitory effect of siderophore active substances from strains 2-8F2 and 2-14F2 against tomato bacterial wilt
3 結論與討論
許多真菌均可產生鐵載體,已報到的有曲霉屬Aspergillus[21]、青霉屬Penicillium[22]、鏈格孢屬Alternaria[23]、木霉屬Trichoderma[24]及白僵菌屬Beauveria[25]等。本研究獲得的2個高鐵載體活性菌株2-8F2及2-14F2分別屬于白僵菌屬和木霉屬微生物(擬球孢白僵菌及云南木霉)。通過文獻檢索,我們發現:布氏白僵菌Beauveriabrongniartii[26]、球孢白僵菌B.bassiana[27]與木霉Trichodermaharzianum[28]、棘孢木霉T.asperellum[29]、綠木霉T.virens[30]、T.peudokoningii[31]均具有鐵載體活性報道,但活性均低于本研究菌株,因此,2-8F2與2-14F2均為同屬微生物中的高鐵載體活性菌株。根據文獻報道,白僵菌屬微生物對交鏈格孢Alternariaalternata[32]、青枯雷爾氏菌[33]、枯斑擬盤多毛孢Pestalotiopsisfunera[34]、烏黑皮殼菌ValsasordidaNits.[35]、紅曲霉Aspergillusruber、松球殼孢菌Sphaeropsissapinea[36]等植物病原菌具有拮抗活性。木霉屬微生物對大麗輪枝菌Verticilliumdahliae、大雄疫霉Phytophthoramegasperma[37]、立枯絲核菌Rhizoctoniasolani[38]、青枯雷爾氏菌[39]、褐根病菌Phellinusnoxius和尖孢鐮刀菌Fusariumoxysporum[40]等植物病原菌具有拮抗活性。本文首次開展了相關屬微生物產鐵載體活性物質對青枯雷爾氏菌抑菌活性研究,有效驗證了分泌型鐵載體真菌對植物病原菌的防控,具有廣譜性。
本研究離體及活體盆栽試驗均發現,在鐵充足條件下,2-8F2與2-14F2兩株菌鐵載體活性物質的活性均顯著降低,表明環境鐵離子是2-8F2與2-14F2菌株發揮抑菌作用的重要影響因子。出現這一現象的可能原因是:鐵載體抑菌效應與其較強的Fe3+競爭螯合能力有關;當環境中鐵離子豐富時,靶標菌株有充足的鐵資源進行生長,從而鈍化了鐵載體物質的鐵競爭性抑菌作用。
本研究從云南省哀牢山國家級自然保護區森林土壤中分離到2株產鐵載體菌株,分別是云南木霉及擬球孢白僵菌。其鐵載體化學結構類型均為異羥肟酸型。兩菌株的鐵載體活性物質對番茄青枯病防治效果高于50%,具有開發成新型植物病原生物拮抗劑的潛力。
