生物素標記重組豬圓環病毒2d基因型毒株感染性克隆的構建與鑒定
2022-08-06 08:50:38林小楓劉梅雪袁瑋藝張瓊歌
中國獸醫雜志 2022年5期
林小楓 , 劉梅雪 , 袁瑋藝 , 張瓊歌 , 朱 磊 , 杜 謙
(1.西北農林科技大學動物醫學院 , 陜西 楊凌 712100 ; 2.楊凌金海生物技術有限公司 , 陜西 楊凌 712100)
豬圓環病毒2型(Porcine circovirus type 2,PCV2)是目前嚴重危害養豬業的一種重要病原。PCV2感染能抑制機體免疫,導致豬圓環病毒病綜合征(Porcine circovirus associated disease,PCVAD),且PCV2在豬群中的持續存在,給養豬業造成了巨大的損失[1]。PCV2的單鏈環狀DNA基因組包含2個主要的編碼框(Open reading frame,ORF),即ORF1和ORF2。可根據ORF2基因序列的差異將PCV2分為8個不同的基因型,其中PCV2a、PCV2b和PCV2d是主要的流行基因型[2]。豬群中最早流行PCV2a基因型,然后轉變為PCV2b,目前正緩慢轉變為PCV2d[3-4]。雖然有報道稱,目前市場上銷售的基于PCV2a基因型毒株的疫苗對于PCV2d基因型毒株的保護效果與PCV2a基因型毒株相當,但臨床仍存在已接種PCV2a基因型毒株疫苗的豬發生PCVAD,因此PCV2臨床毒株可能突破疫苗免疫保護[5]。原核生物素連接酶(Prokaryotic biotin ligase,BirA)是原核生物調控生物素合成的酶,生物素受體肽(Biotin acceptor peptide,BAP)是一種廣泛使用的15 aa標簽肽,其可被插入目的蛋白,使目的蛋白被BirA高效識別并標記生物素[6]。利用BirA-BAP標記系統獲得生物素化PCV2d毒株可為后續進一步研究PCV2d感染突破疫苗免疫保護的具體機制提供材料。
本試驗從陜西咸陽某臨床發病豬場采集17頭仔豬的血清、脾臟和淋巴結等組織樣本,PCR檢測發現其中5頭仔豬為PCV2陽性,經測序發現1頭仔豬感染了PCV2d基因型毒株。將PCV2d基因組orf2序列中226~246 bp替換為BAP編碼序列后,環化并轉染穩定表達BirA的PK15細胞,經連續傳代5次后,獲得生物素標記的重組PCV2d基因型毒株,本結果將為后續開展PCV2d基因型毒株免疫逃避機制研究提供有效的工具。
1 材料與方法
1.1 病料來源 2頭臨床發病仔豬和15頭未發病仔豬的血清、脾臟和淋巴結等組織樣本均采集自陜西咸陽武功縣某養殖場。
1.2 細胞、菌株和質粒 PK15細胞、大腸埃希菌(E.coli)DH5α感受態細胞和pCI-neo-N1質粒,由本實驗室保存;pMD-18T質粒,購自TaKaRa公司。
1.3 主要試劑 2×TaqMaster Mix(Dye plus)和T4連接酶,均購自南京Vazyme公司;SacⅡ、BamH Ⅰ和XhoⅠ等內切酶,均購自TaKaRa公司;M5 DL2 000 plus DNA Marker,購自北京聚合美公司;Lipo6000,購自碧云天公司;鼠抗BirA單克隆抗體,購自NOVUS公司;HRP標記鏈霉親和素,購自Santa Cruz公司;鼠抗PCV2 Cap多克隆抗體,由本實驗室制備并保存。
1.4 試驗方法
1.4.1 引物設計 根據已報道的PCV2(GenBank:MH492006)序列,設計PCV2全長擴增引物PCV2-F和PCV2-R,以及PCV2檢測引物PCV2JC-F和PCV2JC-R;根據PCV2 Cap編碼序列的Loop CD區域(L75PPGGGSN82),設計引入BAP標簽(GLNDIFEAQKIEWHE)序列的Overlap引物BAP-F和BAP-R;根據大腸埃希菌的BirA基因序列(GenBank:M10123.1)設計引物BirA-F和BirA-R。引物由生工生物工程(上海)股份有限公司合成,序列見表1。
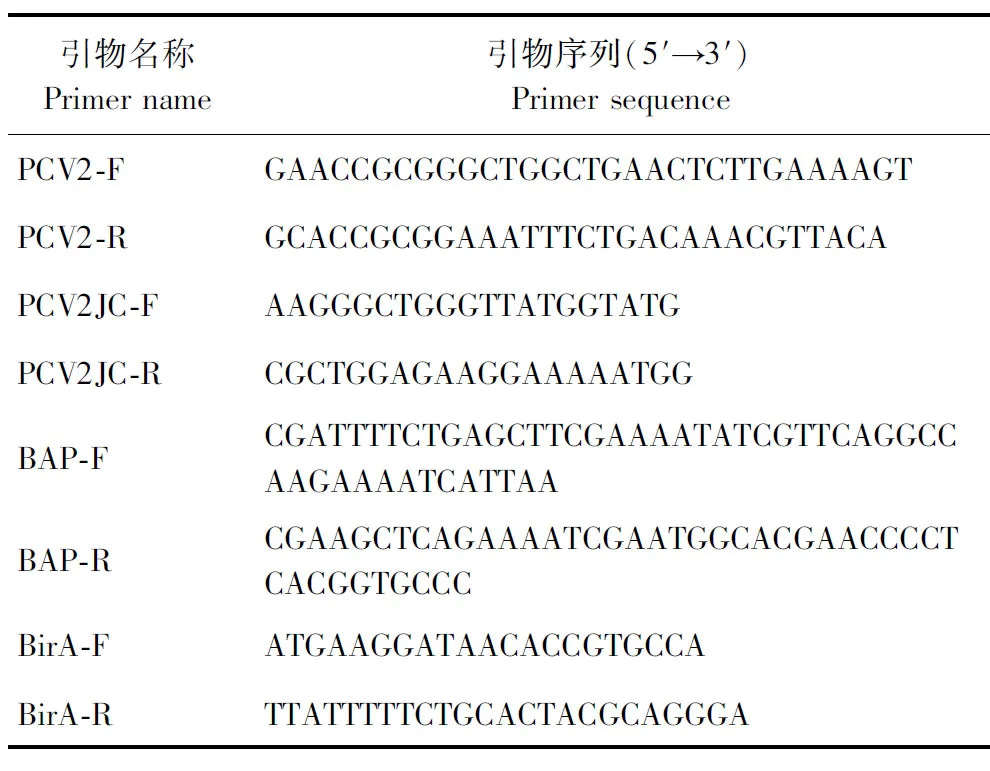
表1 引物序列Table 1 Primer sequences
1.4.2 基因組DNA的提取 將采集的組織樣本或細胞經-80 ℃反復凍融,離心取上清。在收集的上清液中加入1%終體積SDS和0.5 mg/μL終濃度蛋白酶K,顛倒混勻1 min,56 ℃水浴30 min。加入等體積DNA提取液,震蕩混勻5 min后,12 000 r/min離心5 min。吸取上清,加入等體積氯仿,混勻5 min后,室溫12 000 r/min離心5 min。吸取上清,加入1/10體積2 mol/L乙酸鈉和2倍體積無水乙醇,-80 ℃過夜。4 ℃ 12 000 r/min離心15 min,棄上清,加入1 mL 75%乙醇洗滌2次,4 ℃離心棄上清,自然風干后加入ddH2O溶解,置于-20 ℃備用。
1.4.3 PCV2基因組擴增和遺傳進化分析 利用PCV2檢測引物PCV2JC-F和PCV2JC-R對提取的17份組織DNA樣品進行PCR檢測,設置陽性對照和陰性對照。對于PCV2陽性DNA樣品,利用PCV2全長引物進行PCV2全基因組PCR擴增,回收PCR產物,送生工生物工程(上海)股份有限公司測序。為了明確所擴增PCV2的基因型,利用MEGA 5.10軟件,采用鄰接(Neighbor-joining,NJ)法,并進行1 000次自舉重復,對測序結果進行遺傳進化分析。
1.4.4 重組質粒T-PCV2d-BAP的構建 根據獲得的PCV2d DNA中orf2的226~246 bp序列,分別利用Overlap引物BAP-F和PCV2-R、PCV2-F和BAP-R進行PCR擴增,獲得產物片段1和片段2,以片段1和片段2共同為模板,利用PCV2-F和PCV2-R進行PCR擴增,獲得重組PCV2d-BAP目的片段。將其克隆入pMD19-T載體,轉化E.coliDH5α后挑取單克隆,擴大培養并提取質粒,分別進行PCR及SacII酶切鑒定,并進一步測序鑒定,將構建成功的重組質粒命名為T-PCV2d-BAP。
1.4.5 重組質粒pCI-BirA的構建 取3 mLE.coliDH5α飽和菌液,離心棄上清,STE洗滌1次,加入500 μL TE緩沖液,沸水浴10 min后,以12 000 r /min離心15 min,取上清作為PCR模板。利用BirA-F和BirA-R進行PCR擴增,條件為:95 ℃預變性10 min,94 ℃ 40 s,65 ℃ 1 min,72 ℃ 1 min,30個循環后,再于72 ℃延伸10 min。回收PCR產物,利用BamH I和XhoI雙酶切,同時對pCI-neo-N1質粒進行雙酶切,回收酶切產物,用T4連接酶于16 ℃過夜連接,轉化E.coliDH5α后挑取單克隆并擴大培養,提取質粒后經PCR、雙酶切和測序鑒定,將構建成功的重組質粒命名為pCI-BirA。
1.4.6 穩定表達BirA的PK15細胞系的構建及鑒定 將狀態良好的PK15細胞以6×104個/孔鋪制24孔細胞板,置于37 ℃、5% CO2恒溫細胞培養箱培養。待細胞密度達到70%左右時,利用Lipo6000將構建好的重組質粒pCI-BirA轉染至PK15細胞,48 h后加入G418進行篩選,獲得單細胞克隆后擴大培養。收取擴大培養的細胞,提取細胞mRNA,反轉錄為cDNA后,PCR擴增BirA基因編碼序列,同時Western blot檢測細胞中BirA蛋白的表達,PCR和Western blot陽性細胞克隆即為穩定表達BirA的PK15細胞系。
1.4.7 生物素標記PCV2d基因型毒株的獲得及鑒定 利用內切酶SacII將T-PCV2d-BAP酶切,獲得PCV2d-BAP編碼序列,將其以T4連接酶于16 ℃連接過夜,連接產物經純化后,轉染至穩定表達BirA的PK15細胞,盲傳5代后,收取細胞,裂解并收集上清,提取DNA,利用PCV2特異性引物進行PCR檢測,同時將獲得的細胞上清再次感染PK15細胞,裂解細胞后,以鼠抗PCV2 Cap多克隆抗體和HRP標記的鏈酶親和素進行Western blot檢測,陽性毒株即為生物素標記PCV2d基因型毒株。
2 結果
2.1 臨床樣本的檢測及PCV2d全基因組擴增 將采集的2頭發病仔豬和15頭未發病仔豬樣本,分別提取DNA后,以PCV2特異性檢測引物進行PCR擴增,結果顯示,2頭發病仔豬均為PCV2陽性,15頭未發病仔豬中有3頭為PCV2陽性(圖1A)。分別將5頭PCV2核酸陽性仔豬樣本中的PCV2全基因組DNA進行PCR擴增,均得到大小約為1 760 bp的目的條帶(圖1B)。對目的條帶進行測序,并進行遺傳進化分析,結果顯示,擴增的5條目的條帶中有1條為PCV2d序列,擴增自1頭發病仔豬的組織樣本,其余4條為PCV2b序列,分別來源于另一頭發病仔豬和3頭未發病仔豬(圖1C)。
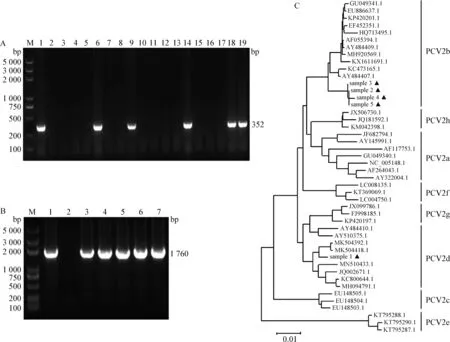
圖1 臨床樣本PCR檢測及PCV2d全基因組遺傳進化分析Fig.1 PCR detection of clinical samples and genetic evolution analysis of PCV2d whole genomeA:17份臨床樣本PCV2特異性PCR擴增(M:DNA標準DL2 000 plus; 1:陽性對照; 2:陰性對照; 3~17:臨床未發病仔豬組織樣本; 18、19:臨床發病仔豬組織樣本);B:5份陽性樣本PCV2全基因組PCR擴增(M:DNA標準DL2 000 plus; 1:陽性對照; 2:陰性對照; 3~7:PCV2陽性臨床樣本);C:PCV2全基因組遺傳進化樹(▲:本試驗擴增序列)A:PCV2 specific PCR amplification of 17 clinical samples(M:DNA Marker DL2 000 plus; 1:Positive control; 2:Negative control; 3-17:Clinical tissue samples from non-diseased pigs; 18,19:Clinical tissue samples from diseased pigs);B:PCV2 whole genome PCR amplification of 5 positive samples(M:DNA Marker DL2 000 plus; 1:Positive control;2:Negative control; 3-7:PCV2 positive clinical samples);C:Phylogenetic tree of PCV2 genomes(▲:Sequences amplified in this study)
2.2 重組質粒T-PCV2d-BAP的構建 根據獲得的1株PCV2d全基因組DNA中orf2的226~246 bp序列,設計引入BAP序列的Overlap引物,分別進行PCR擴增,獲得大小約為842 bp的PCR片段1和大小約為1 028 bp的PCR片段2(圖2A)。以片段1和片段2同時為模板,用PCV2全基因組引物進行PCR擴增,獲得大小約為1 790 bp的PCV2d-BAP目的條帶(圖2B),將其克隆入pMD19-T載體,經PCR及SacⅡ酶切鑒定,均獲得了預期大小的目的條帶(圖2C),測序鑒定正確,表明重組質粒T-PCV2d-BAP構建成功。
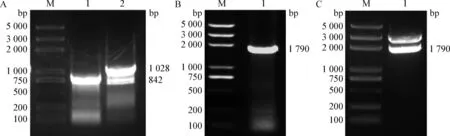
圖2 重組質粒T-PCV2d-BAP的構建Fig.2 Construction of recombinant plasmid T-PCV2d-BAPA:PCV2d片段1和片段2的PCR擴增(M:DNA標準DL2 000 plus; 1:片段1; 2:片段2); B:PCV2d-BAP的PCR擴增(M:DNA標準DL2 000 plus; 1:PCV2d-BAP);C:重組質粒T-PCV2d-BAP的Sac Ⅱ酶切鑒定(M:DNA標準DL2 000 plus; 1:Sac II酶切鑒定產物)A:PCR amplification of fragment 1 and fragment 2 of PCV2d(M:DNA Marker DL2 000 plus; 1:Fragment 1; 2:Fragment 2); B:PCR amplification of PCV2d-BAP(M:DNA Marker DL2 000 plus; 1:PCV2d-BAP);C:Identification of recombinant plasmid T-PCV2d-BAP by Sac Ⅱ enzymes digestion(M:DNA Marker DL2 000 plus; 1:Sac Ⅱ enzymes digestion products)
2.3 重組質粒pCI-BirA的構建 設計E.coliDH5αBirA基因特異性引物,經PCR擴增,獲得大小約為966 bp的目的條帶(圖3A),將其克隆入pCI-neo載體,經雙酶切鑒定,獲得大小約為966 bp的目的條帶(圖3B),測序鑒定正確,表明已成功構建pCI-BirA重組質粒。
圖3 重組質粒pCI-BirA的構建Fig.3 Construction of recombinant plasmid pCI-BirAA:BirA基因的PCR擴增(M:DNA標準DL2 000 plus; 1:BirA);B:重組質粒pCI-BirA的雙酶切鑒定(M:DNA標準DL2 000 plus; 1:BamH I和Xho I雙酶切產物)A:PCR amplification of BirA gene(M:DNA Marker DL2 000 plus; 1:BirA);B:Identification of recombinant plasmid pCI-BirA by double enzymes digestion(M:DNA Marker DL2 000 plus; 1:BamH I and Xho I double enzymes digestion products)
2.4 穩定表達BirA的PK15細胞系的構建及鑒定 將構建好的pCI-BirA重組質粒轉染PK15細胞,48 h后加入G418進行篩選,獲得單細胞克隆后擴大培養(圖4A),提取細胞mRNA并反轉錄后,PCR擴增BirA基因編碼序列,結果顯示獲得了大小約為966 bp的目的條帶(圖4B);Western blot結果顯示,獲得的細胞克隆表達BirA蛋白(圖4C),與預期大小一致,表明成功獲得了穩定表達BirA的PK15細胞系。
圖4 穩定表達BirA的PK15細胞系的構建及鑒定Fig.4 Construction and identification of PK15 cell line stably expressing BirAA:G418篩選后單克隆擴大培養的PK15細胞(100×);B:PK15細胞系BirA mRNA的PCR鑒定(M:DNA標準DL2 000 plus; 1:BirA mRNA); C:PK15細胞系BirA蛋白的Western blot鑒定(WT:野生型PK15細胞; BirA:表達BirA的PK15細胞A:Expanded PK15 cells after G418 screening (100×);B:PCR identification of BirA mRNA in PK15 cell line(M:DNA Marker DL2 000; 1:BirA mRNA);C:Western blot identification of BirA protein in PK15 cell line(WT:Wild-type PK15 cells; BirA:PK15 cells expressing BirA)
2.5 生物素標記PCV2d基因型毒株的獲得及鑒定 將環化的PCV2d-BAP轉染穩定表達BirA的PK15細胞,盲傳5代,經檢測,成功獲得大小約為352 bp的目的片段(圖5A)。同時將獲得的上清感染PK15細胞,裂解細胞后進行Western blot檢測,結果顯示,以鼠抗PCV2 Cap多克隆抗體和HRP標記的鏈酶親和素均能檢測到大小約為29 kDa的目的條帶(圖5B),表明成功獲得了生物素標記PCV2d基因型毒株。
圖5 生物素標記PCV2d基因型毒株的獲得及鑒定Fig.5 Obtain and identification of biotin-labeled PCV2d genotype strainA:細胞裂解上清PCV2特異性PCR鑒定(1:陽性對照; 2:細胞裂解上清; 3:陰性對照);B:生物素標記PCV2d基因型毒株的Western blot鑒定(1:PCV2陽性對照; 2:生物素標記PCV2d基因型毒株; 3:陰性對照)A:PCV2 specific PCR identification of cell lysis supernatant(1:Positive control; 2:Cell lysis supernatant; 3:Negative control); B:Western blot identification of biotin-labeled PCV2d genotype strain(1:PCV2 positive control; 2:Biotin-labeled PCV2d genotype strain; 3:Negative control)
3 討論
PCV2是目前已知最小的動物病毒,其基因組大小約為1 766~1 768 nt的單鏈環狀DNA。在單鏈DNA病毒中,PCV2基因組具有最高的突變概率,每年能達到約1.2×10-3突變/位點[7]。PCV2基因組的這些突變使其可以被分為不同基因型,分別為PCV2a、PCV2b、PCV2c、PCV2d、PCV2e、PCV2f、PCV2g和PCV2h[2,5]。在目前已報道的PCV2序列中,PCV2b基因型最多,其次是PCV2d基因型,第三是PCV2a基因型,這3種PCV2基因型均呈世界性分布[2]。從PCV2序列報道的時間上來看,最早報道的是PCV2a基因型,但大約從2002年開始,PCV2b基因型的報道增多,特別是2005年以后,PCV2b基因型開始成為主要的流行基因型[2]。隨著PCV2基因組的持續突變,2010年以后PCV2d基因型的報道量急劇增加,并呈現逐漸取代PCV2b基因型成為主要流行基因型的趨勢[2]。更重要的是,有研究顯示,雖然目前廣泛使用的基于PCV2a基因型毒株的疫苗能使免疫后的仔豬在被PCV2a、PCV2b或PCV2d基因型毒株感染時具有相似的保護作用,但在PCV2b和PCV2d基因型毒株感染的仔豬體內中和抗體的產生量相對較少[8]。目前,世界主要養豬國家已廣泛使用PCV2疫苗對豬群進行免疫,但與此同時PCV2d基因型毒株報道增多,這可能與疫苗的選擇壓力有關[5]。事實上,早前已有報道顯示,PCV2在疫苗選擇壓力下已經發生了免疫逃避進化趨向[9]。因此,對豬群中PCV2流行情況的持續監控,以及對流行PCV2基因型的進一步深入研究,將有助于更好地防控豬圓環病毒病。在本試驗中,對臨床發病的2頭仔豬和同豬群未發病的15頭仔豬的組織樣本進行了PCV2核酸檢測,結果顯示,2頭發病仔豬均為PCV2陽性,15頭未發病仔豬中有3頭為PCV2陽性,經進一步測序發現,兩頭發病仔豬中一頭感染了PCV2d基因型毒株,另一頭感染了PCV2b基因型毒株,3頭未發病仔豬均為PCV2b基因型毒株感染,推測本試驗檢測豬群中感染的PCV2基因型仍以PCV2b為主,但出現了PCV2d基因型毒株導致的感染和發病。
PCV2基因組有2個主要的ORF,分別編碼復制酶Rep/Rep′和衣殼蛋白Cap。前期研究發現,PCV2 Cap編碼序列的Loop CD區域(L75PPGGGSN82)可在不改變病毒衣殼結構和入胞能力的情況下插入外源基因[10]。因此,本試驗基于臨床獲得的PCV2d基因型毒株基因組,將Cap的Loop CD區域替換為生物素受體肽BAP編碼序列,構建了能融合表達BAP標簽的PCV2d基因型毒株全基因組。BAP是生物素連接酶BirA的底物,BirA-BAP系統是一種原核生物蛋白質翻譯后生物素化修飾系統[11]。BirA可特異性地識別BAP,并將生物素分子共價結合至BAP的賴氨酸殘基上。這種利用BirA-BAP系統的生物素化標記,已被用于純化蛋白、標記抗體、蛋白功能研究和構建生物素標記重組病毒等[11-12]。本試驗將構建的能融合表達BAP標簽的PCV2d全基因組DNA經環化后,轉染穩定表達BirA的PK15細胞,利用BirA-BAP系統獲得了生物素標記的PCV2d基因型毒株。這種生物素標記的PCV2d基因型毒株將有助于進一步研究PCV2d基因型毒株的感染與免疫逃避機制。但是,這種PCV2d基因型毒株衣殼上標記的生物素能否持續穩定的存在,還需要多代次的擴增后進一步檢測。
綜上所述,本試驗從臨床發病豬群中獲得了1株PCV2d基因型毒株全基因組序列,以此毒株序列為基礎,利用BirA-BAP系統構建了生物素標記PCV2d基因型毒株感染性克隆,獲得了生物素標記的PCV2d毒株,為后續開展PCV2d基因型毒株免疫逃避機制研究提供了基礎。
