重慶喀斯特生境中楨楠種群結構與動態特征
2022-08-03 02:53:06何雪娜王佳敏劉俊婷譚興旺劉錦春
生態學報 2022年12期
雷 穎,何雪娜,王佳敏,劉俊婷,譚興旺,趙 彬,劉錦春,*
1 西南大學生命科學學院,三峽庫區生態環境教育部重點實驗室,重慶市三峽庫區植物生態與資源重點實驗室,重慶 400715 2 重慶市永川區國有林場,重慶 402100
種群結構與動態特征是種群生態學研究的兩大核心[1—2],是植物種群的生物學特性對環境長期適應和選擇的結果[3]。珍稀漸危樹種在植物的系統演化和遺傳育種中占據著重要地位,且在經濟、科研和文化等方面有著較高的價值,其數量稀少,生境特殊[4]。珍稀保護樹種在特定生境下的種群結構和動態特征以及繁殖更新情況,可以反映珍稀樹種與環境之間的相互關系,對分析種群的現實情況、預測種群未來的走向以及實施合理的保護措施有重要意義。
楨楠(PhoebezhennanS. Lee)為樟科(Lauraceae)楠屬(PhoebeNees)的高大喬木,是珍稀漸危樹種[5—6]。由于歷年來對楨楠的濫砍濫伐,其原生林幾乎被毀,加上楨楠后期生長緩慢[7]和遺傳多樣性較低[8]等原因,導致現存資源有限且分布零散[9]。研究表明,楨楠喜濕耐蔭,適宜在酸性土壤的山腰山谷中生存[10],野生多散生于海拔1500 m以下的亞熱帶常綠闊葉林中[5,11]。然而,在野外調查中發現重慶市大足區及永川區的喀斯特生境中也存在長勢良好的楨楠林。
喀斯特生境是一種整體資源量低的脆弱生境,其巖石裸露率高、土壤淺薄且呈斑塊化分布、土壤營養貧瘠且持水性能較差[12]。由于生境特點和人類活動的影響,喀斯特地區的植被常呈退化之勢,喬木樹種生存困難。受喀斯特生境獨特地貌形成的地理屏障限制,植物種子很難通過水流、風力和重力等非生物因子進行長距離傳播而到達到適宜其萌發的微生境中;且喀斯特的特殊生境也不利于后期種子的萌發和幼苗的生長,從而對后期植物種群的空間分布產生重要影響[13]。因此我們推測喀斯特地區的楨楠種群可能存在結構不穩定、更新困難等問題。
目前對于楨楠的研究大多集中在抗性生理[14—15]、營養生理[16—17]和群落特征[18—20]等方面,也有少數研究關注了部分楠木種群的結構和動態特征。如吳大榮等[21]研究了福建羅卜巖閩楠種群,發現該種群空間分布格局總體上具有由集群向隨機分布的趨勢,具有較強的自然更新的能力。徐振東[18]和李賢碧等[22]研究表明,貴州思南縣楠木種群和湖北利川楠木種群所在群落物種組成均較豐富,更新能力強。然而,曾凡勇和馮邦賢[23]的研究卻發現,德江沙溪楠木群落物種多樣性整體偏低,更新能力較弱。可見,楠木種群結構和動態特征與其所在的群落環境及群落物種組成有一定的關系。
因此,本研究通過對重慶喀斯特生境中的兩個楨楠種群的年齡結構、數量動態特征與空間分布格局的定量研究,明確其在喀斯特生境中的生存現狀,揭示其動態規律和未來發展趨勢,以期為楨楠種群的保護、群落更新和植被恢復提供理論依據。
1 研究區概況
研究區域選擇重慶市大足區高坪鎮紙槽溝和永川區太嵐坳,是楨楠的天然群落分布區。該區四季分明,春季回溫早,夏季氣溫高,降雨分布不均勻,光照充足,雨熱同季,屬于亞熱帶季風氣候。地貌以丘陵、山地為主,坡地面積較大。土壤為黃壤土,中性至微酸性[7]。
重慶市大足區高坪鎮紙槽溝(29°50′N,105°39′E),海拔480 m,境內分布的野生楠木占地約200 hm2,集中成片達107 hm2,林內百年樹齡以上2萬余棵,主要以黃絨潤楠(Machilusgrijsii)為主。該楠木林已有兩三百年歷史,一直由該地村民守護。該區年均無霜期323 d,全年均降水量727 mm,集中在5—9月。該區域內有高等植物24科28屬31種。
重慶市永川區太嵐坳(29°33′N,105°53′E)大碑寺公路附近,海拔628 m,境內分布的天然楨楠占地約64 hm2,林內最高樹齡有上百年。當地村民從1957年起對此林進行嚴格管理,現林場為楨楠建立了一套完整的保護體系,實現全方位立體化的保護。該區年均無霜期317 d,年均降水量1015 mm,集中在5—8月。該區域內有高等植物26科36屬38種。
2 研究方法
2.1 樣地設置與調查
由于兩個區域的楨楠林林分均比較簡單,因此設計的樣方面積均為10 m×10 m。根據地形特點,大足區的楨楠林中共設置7個樣方,呈“一”字形排列,永川區共設置10個樣方,呈環狀分布。調查并記錄所有樣方內楨楠的個體數、樹高、胸徑、高度和冠幅等指標。同時測定和記錄各樣地的海拔、坡度、坡向、郁閉度、草本覆蓋度和凋落物厚度(表1)。
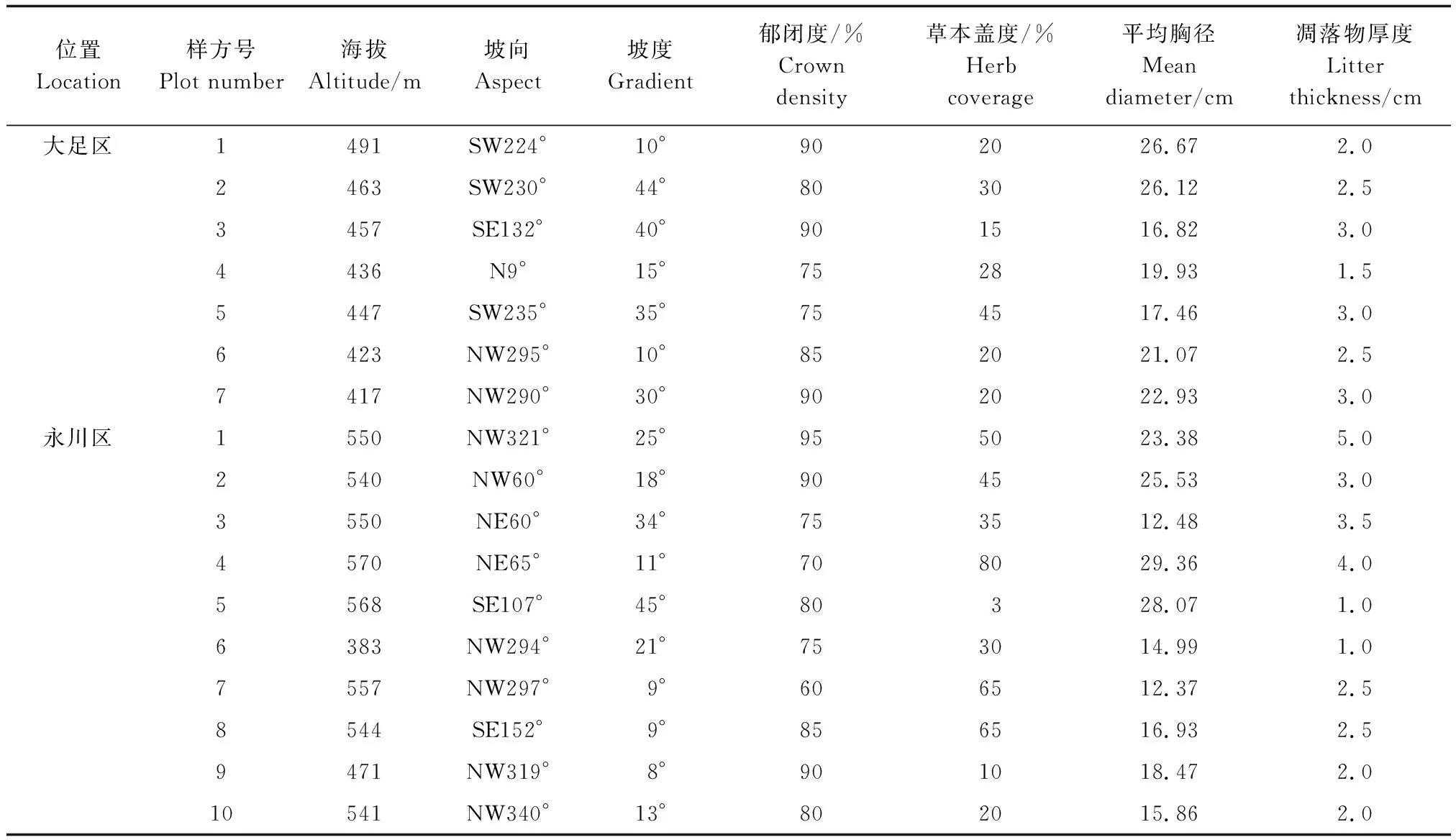
表1 楨楠種群環境條件Table 1 The environmental conditions of P. zhennan population
2.2 種群徑級劃分
由于同一樹種的年齡結構(齡級)和徑級對相同的環境所呈現出的規律具有一致性[24],再加上樹木年齡獲取比較困難,因此采用徑級結構代替年齡結構的方法。根據所得調查數據和已有研究[7,25—26],將楨楠劃分為8個齡級:第Ⅰ齡級,幼苗,H<2.5 m;第Ⅱ齡級,幼樹,H≥2.5 m,胸徑<10 cm;第Ⅲ齡級,中樹,10 cm≤胸徑<15 cm;第Ⅳ齡級,中樹,15 cm≤胸徑<20 cm;第Ⅴ齡級,成樹,20 cm≤胸徑<25 cm;第Ⅵ齡級,成樹,25 cm≤胸徑<30 cm;第Ⅶ齡級,成樹,30 cm≤胸徑<35 cm;第Ⅷ齡級,老樹,胸徑≥35cm。
2.3 靜態生命表及存活曲線
靜態生命表是用特定時間點上調查種群內各個年齡組上存活狀況的數據編制而成的[27]。
在調查中系統誤差不可避免,有可能會出現生命表中的死亡率為負值的情況,這是由于靜態生命表反映的是世代重疊的年齡動態歷程中的一個特定時間,而不是該種群的全部生活史[28]。因此在編制生命表之前對實測數據進行勻滑處理[29]得到ax(實際存活個體數經勻滑處理后所得存活數),編制出楨楠種群的靜態生命表。
此外,采用Hett和Loucks[30]的方法檢驗楨楠種群的存活曲線。指數方程Nx=N0e-bx描述Deevey-Ⅱ型存活曲線,冪函數Nx=N0x-b描述Deevey-Ⅲ型存活曲線。
2.4 種群年齡結構及動態分析
運用陳曉德[31]的種群結構動態量化分析方法,得到楨楠種群年齡結構的動態指數,從而對種群動態(V)進行定量的描述。在方法中,Vn取正值、負值、零值時分別反映種群兩相鄰齡級個體數量的增長、衰退、穩定的結構動態關系;Vpi、V′pi取正值、負值、零值分別反映種群個體數量的增長、衰退、穩定。計算公式如下:
式中,Sn與Sn+1分別為第n和第n+1齡級的個體數;k為種群大小數量級。
2.5 種群空間結構分析
采用擴散系數(C),并進行t檢驗,從而判斷楨楠的空間分布格局,同時使用負二項指數(K)、叢生指數(I)、平均擁擠度(M*)和聚塊性指數(PAI)、以及聚集度指數(Ca)等多個指標擬合楨楠種群的分布格局[32]。具體計算公式如下:
(1)擴散系數C
C=V/m
當C>1、C=1 和C<1時,種群表現為聚集、隨機和均勻分布。
(2)負二項指數K
當K<0時為均勻分布;K>0為聚集分布,低K值表示叢生比較顯著,即聚集強度大,高K值表示叢生程度低,聚集強度弱;當K值趨于無窮大時(一般為K<8),種群逼近泊松分布。
(3)叢生指數I
叢生指數I>0、I=0和I<0時,種群分別表現為為聚集、隨機和均勻分布。
(4)平均擁擠度和聚塊性指標
PAI=M*/m
M*為平均擁擠度,代表的是樣方內生物個體的擁擠程度;PAI為聚塊性指標,當PAI>1時,為聚集分布;PAI=1時,為隨機分布;PAI<1時,為均勻分布。
(5)聚集指數
當Ca<0、Ca=0、Ca>0時,種群分別表現為均勻分布、隨機分布、聚集分布。
以上所有公式中V和m均分別表示種群方差和均值。
3 結果與分析
3.1 種群年齡結構
大足區與永川區兩個楨楠種群年齡結構均為幼苗、幼樹株數多,中、成、老樹個體比例大致相同,整體為增長型(圖1)。在楨楠種群實際調查中,樣方內的幼苗由于生存空間的限制不可能全部存活,且楨楠幼苗20 cm以下的個體成活率較低。大足區的第Ⅰ齡級幼苗平均高度0.42 m,0.2 m以上的幼苗占總體幼苗的60%(16棵),由此來看,大足區楨楠種群各齡級數目相當,幼苗、幼樹、中樹、老樹比例基本相同,大足楨楠種群的實際年齡結構應該為穩定型。永川區的0.2 m以上的幼苗占總體幼苗的74%(248棵),第Ⅰ齡級幼苗遠大于其他齡級,但永川區楨楠種群依然呈幼樹多的增長型。
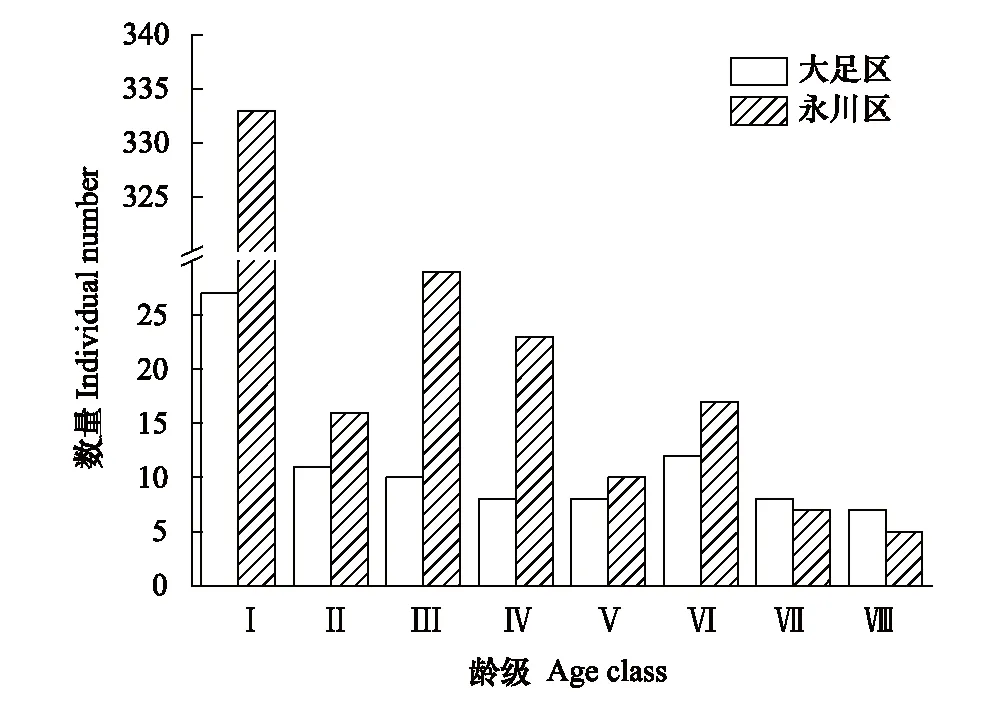
圖1 楨楠種群齡級結構圖Fig.1 Age class structure diagram of P. zhennan populationⅠ—Ⅷ表示楨楠的年齡結構劃分為第1—8齡級
3.2 靜態生命表與存活曲線
隨著徑級的增加,兩個種群的標準化存活個體數(lx)、消失率(Kx)和死亡率(qx)逐漸減小,說明楨楠幼苗、幼樹的死亡率較高,種群易受外界環境的干擾(表2)。大足區楨楠種群的生命期望(ex)在幼苗、幼樹和中樹(Ⅰ—Ⅳ齡級)階段較高,并且隨著個體的生長而下降,符合種群生態學特征。永川區種群中樹的生命期望(ex)達到頂峰,說明該種群正處于活力旺盛的時期,此后開始逐漸降低。
大足區楨楠種群前期死亡率較高,后期較低(表2、圖2);永川區楨楠種群幼苗幼樹死亡率較高(表2、圖2)。兩個楨楠種群存活曲線均與Deevey-Ⅲ型擬合程度較高(表3)。
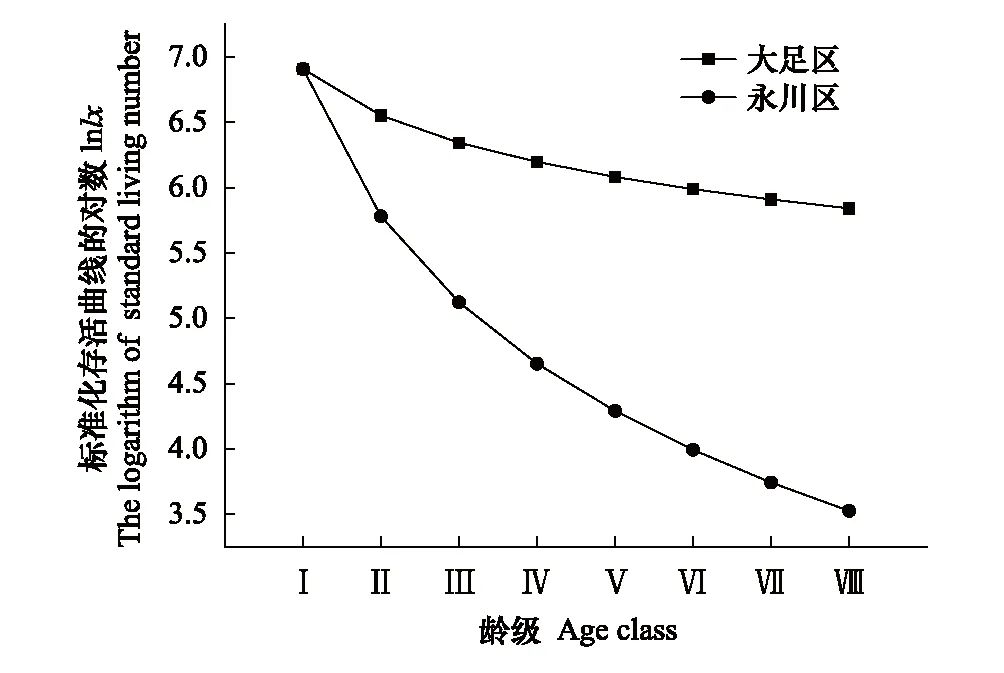
圖2 楨楠種群存活曲線Fig.2 Survival curve of P. zhennan population
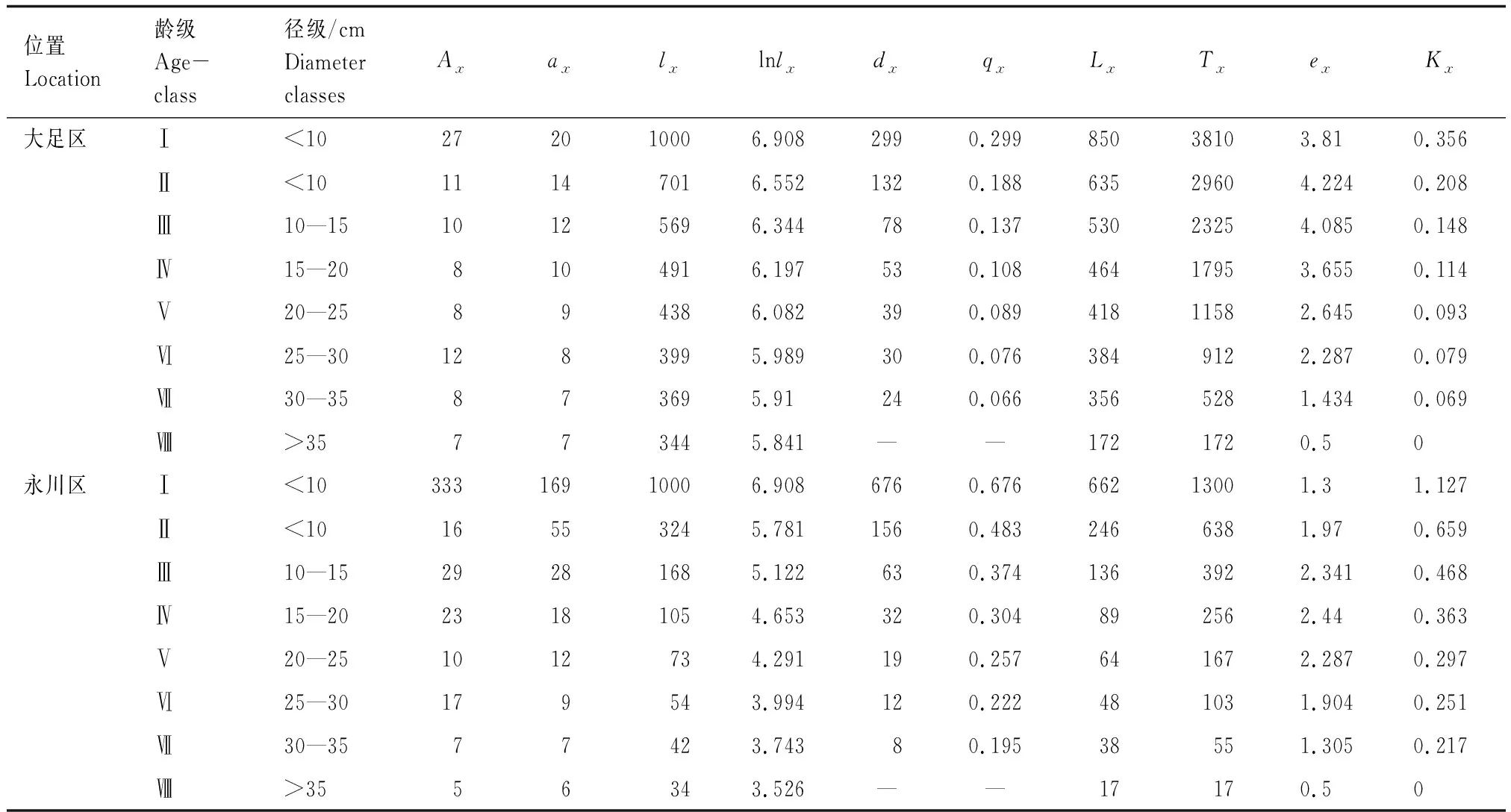
表2 楨楠種群靜態生命表Table 2 Static life table of P. zhennan population
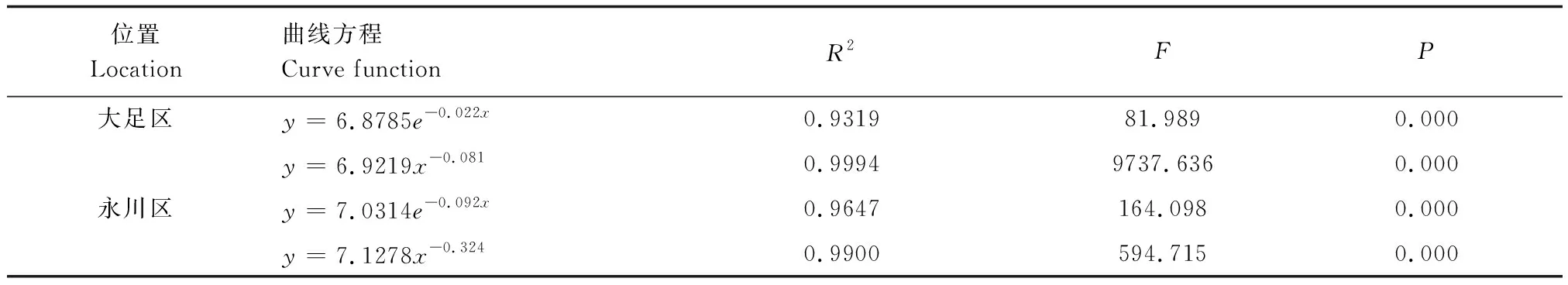
表3 楨楠種群存活曲線的檢驗模型Table 3 Test models of survival curves of P. zhennan population
3.3 種群年齡結構的動態變化分析
大足區楨楠種群在V1、V2、V3、V6和V7均大于0,種群呈現增長的結構動態,V4等于0,種群呈穩定狀態,V5小于0,表明這個齡級的楨楠種群呈衰退狀態;永川區的V2和V5均小于0,種群呈衰退狀態(表4)。

表4 楨楠種群年齡結構動態指數/%Table 4 Dvnamic indices of P. zhennan population age structure
種群總體動態變化指數分析結果顯示:大足區(Vpi=25.40%)和永川區(Vpi=77.40%)種群動態指數均大于0,種群整體結構屬于增長型(表4)。當不忽略外部干擾時,種群齡級結構動態指數V′pi=0.45%和V′pi=1.94%均趨于0,且種群結構對隨機干擾的敏感指數,即隨機干擾風險的極大值Pmax=1.79%(大足)、Pmax=2.50%(永川)(表4),說明永川的楨楠種群結構對隨機干擾的敏感性更大。因此,當兩個種群在受到外部干擾的情況下,大體上仍然是增長型的種群,只不過增長速度較慢,并且對外界干擾敏感度高。
3.4 楨楠種群空間分布格局
大足區楨楠種群呈隨機分布,聚集指數為0.07;永川區楨楠種群呈聚集分布,聚集指數為5.35(表5)。兩個種群在不同齡級上均呈聚集分布,從聚集強度來看,大足區和永川區楨楠種群均為幼苗聚集程度最大(表6)。
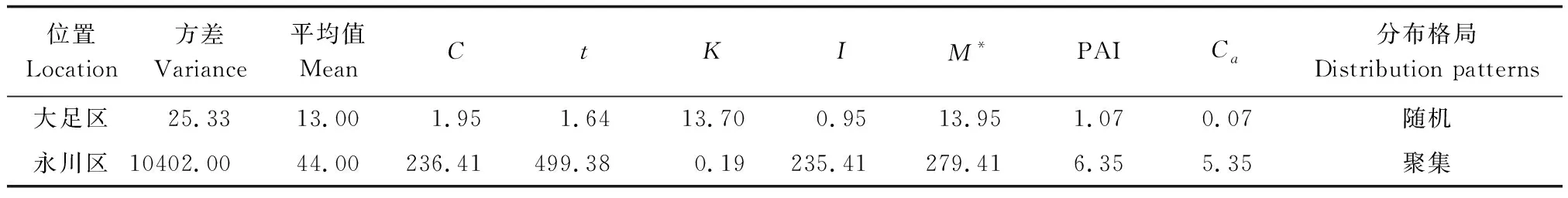
表5 楨楠種群分布格局Table 5 Distribution pattern of P. zhennan population
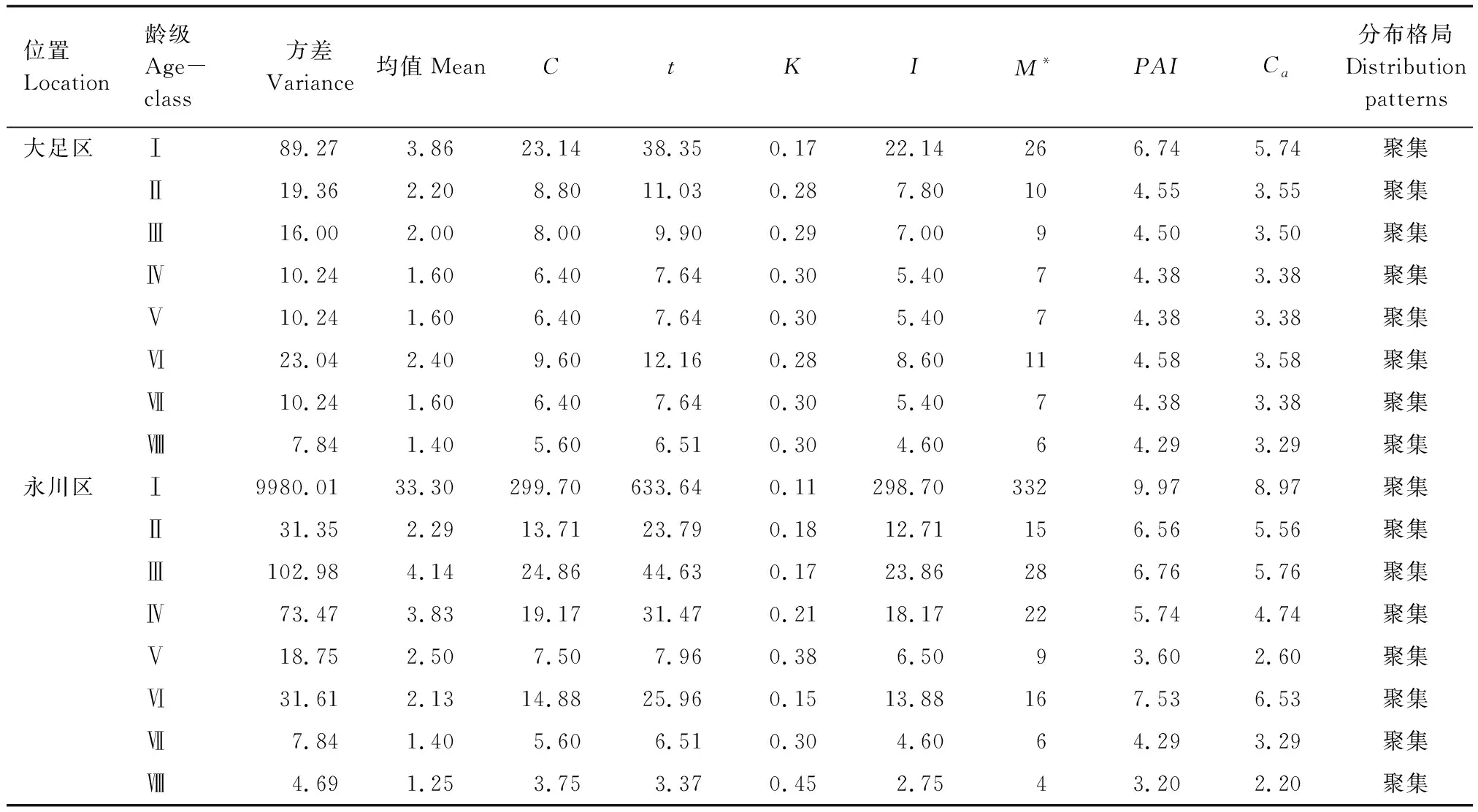
表6 楨楠不同徑級分布格局Table 6 Distribution pattern of different diameter classes in P. zhennan
4 討論
多數研究者認為楨楠適合生長在土壤深厚、溫潤潮濕的地方,而喀斯特地區土層淺薄,且容易出現干濕交替的現象,不宜楨楠生長。然而,大足區楨楠種群年齡結構屬于穩定型,永川區種群年齡結構屬于增長型,這與種群動態分析的結果一致,說明楨楠也能在喀斯特地區生長,但是兩個區域的種群動態量化分析中仍存在局部齡級間衰退的現象。
大足區種群成樹階段呈衰退狀態,永川區的幼樹和成樹也出現類似的情況,即在幼樹、成樹階段呈衰退型。由于兩地區的楨楠一直受到當地村民的保護,因此推測該階段的生存壓力主要是環境和種群自我調節導致的。楨楠幼樹有一定的耐陰性[33],而成樹對光的需求逐漸增強[34],且楨楠林郁閉度均在70%以上。隨著齡級的增長,所需養分和光照逐漸增加,導致部分競爭力弱的成樹無法進入林冠層。Janzen[35]提出動物取食、幼苗競爭的強度會受種子/幼苗與母株的距離有關,越近發生的頻率越高。由于永川區20 cm以下的幼苗較多且呈聚集型分布(5號、6號樣地凋落物厚度較小,種子萌發率遠高于其他樣地),推測其密度效應較強,種內競爭過于激烈且幼苗數量很可能遠遠超過了當地的環境容納量,大部分幼苗無法成功轉向幼樹階段,所以在一定范圍內出現大量幼苗,且幼苗都在母株周圍,大大提高了楨楠幼苗的死亡率。這與Comita和Hubbell等[36]研究的幼苗密度與耐陰性對幼苗存活影響的結果一致。再加上喀斯特地區土壤資源匱乏,水土易從巖石裂隙中流失,雨后即處于干旱缺水的狀態[37],這對抵抗能力差的幼苗、幼樹來說是致命,因此呈衰退狀態。
從生命表、存活曲線和種群動態分析來看,大足區和永川區楨楠種群前期幼苗和幼樹死亡率較高,此后死亡率逐漸下降。從種群年齡結構動態變化指數來看,在不忽略外部干擾時,兩個楨楠種群整體結構呈增長型,這與湖北來鳳縣楨楠種群[9]的調查結果相反。其主要原因是大足區和永川區的楨楠種群長期以來受到人為干擾的強度較小,而來鳳縣的楨楠受人為砍伐嚴重。但這兩個區域的楨楠種群也存在動態指數小于0的階段,說明兩個種群增長緩慢且對外界干擾敏感度高。
楨楠種群的空間分布格局受自身繁殖方式和環境的影響較大。整體而言,大足區楨楠種群的空間分布格局為隨機分布,永川區楨楠種群為集群分布。研究表明,集群分布是楠木種群常見的分布方式[21]。原因是楨楠的種子傳播能力不強,主要是靠自由掉落和鳥類傳播,因此常聚集在母樹附近,呈現聚集分布。永川區楨楠種群集群分布,一是與楨楠的種子傳播力有關,另一方面也與其地形原因不易擴散有關。這也可能是對喀斯特環境的一種積極響應策略,因為聚集分布可以增強種群抵抗外界干擾的能力。而大足區的楨楠種群呈現隨機分布的狀況,其原因可能是種群內部之間的競爭導致的,幼苗充足,且幼苗到成樹之間的競爭較為激烈,因而對資源(光照、水分和養分等)的需求加大,競爭加劇,從而形成均勻分布的格局。同時,大足區與永川區的森林小氣候的不同,可能也是導致楨楠種群空間分布格局差異的另一原因,永川區的年平均降雨量比大足區多288 mm,土壤微酸性,海拔較高,因此永川區更適合楨楠的生存,且在一定范圍內可利用的資源相對豐富,能容納的楨楠就更多。此外,大足區楠木林的范圍是永川區楨楠林的3倍左右,可能是由于兩者的尺度不同導致調查結果有差異,這也能看出楨楠種群發育過程中,分布格局由集群分布向隨機分布發展。
可見,重慶大足區與永川區兩個楨楠種群生存現狀良好,整體呈現穩定趨勢,但局部齡級間仍存衰退現象。與我們的推測相反,喀斯特生境中楨楠種群仍有較強的更新能力,但因種群生長周期長[7],遺傳多樣性較低[8],對外部干擾敏感,適應能力較差。因此,對楨楠種群的保護仍不能掉以輕心,需要制定適宜的保護措施,進行科學合理的管理。同時建議在幼苗幼樹死亡率高的情況下可以采用人工輔助措施,將幼苗從聚集的地方移植一部分出去,減少內部的競爭壓力,待幼苗長成小樹時再將其移植到合適的野外分布區,在現有條件下不斷擴大楨楠資源,創造更多的生態價值和經濟價值。
