微生物亞硝化抑制劑對風干腸風味物質形成的影響
2022-07-31 01:45:20陳援援馬凱華張雨婷鄭翔飛張新渝馬儷珍
食品與機械 2022年7期
陳援援 馬凱華 張雨婷 鄭翔飛 張新渝 馬儷珍
(天津農學院食品科學與生物工程學院,天津 300384)
風干腸是一種天然發酵香腸,以其獨特的風味、色澤和質地而聞名于中國東北地區。傳統的風干腸主要依靠內源酶和存在于原料中的“野生”微生物起作用,這些“野生”微生物主要包括革蘭氏陰性菌、氧化酶陽性菌、需氧菌和少量革蘭氏陽性菌[1]。當生長環境適宜時,這些革蘭氏陽性菌,如乳酸菌和葡萄球菌,可能成為優勢菌群[1-2]。因此,對天然菌群的依賴會導致風干腸品質不一[3]。為了使生產過程標準化,一些發酵劑被用于縮短發酵時間,提高產品質量,并保持最終發酵產品的獨特風味[4]。近年來,有關接種微生物發酵以改善產品風味,確保食品安全的研究較多[5],但對其發酵機理和風味化合物形成的研究還需深入。Lu等[6]研究了不同發酵溫度下酸肉菌群與代謝產物的關系,己醛、苯甲醛、壬醛、(E,E)-2,4-癸二烯醛、1-辛烯-3-醇和辛醛被確定為酸肉中乳酸桿菌產生的揮發性有機化合物,為風味的主要來源。Hongthong等[7]研究了接種植物乳桿菌對Isan香腸品質的影響,發現接種107CFU/g植物乳桿菌后的樣品在感官評價上具有更高的硬度和更強烈的風味,并提高了香腸的亮度值。Bao等[8]研究發現,接種混合發酵劑乳酸乳球菌和竇氏魏斯氏乳酸菌能改善臭鱖魚的品質、促進風味的形成和縮短發酵時間。發酵肉制品中產生風味的主要途徑有蛋白質降解、脂質氧化、美拉德反應和微生物4種[9]。蛋白質降解過程中產生的氨基酸代謝產物占總揮發性化合物的6%[10],在風味形成中起著重要作用。氨基酸可以通過微生物轉氨酶轉化為酮酸,作為重要的中間體,α-酮酸可以直接或間接代謝成相應的醛類化合物。此外,醛可以被醇脫氫酶和醛脫氫酶轉化為相應的醇和羧酸[11]。Chen等[12]研究發現,接種混合發酵劑組風干腸氨基酸代謝產生的化合物水平高于對照組(P<0.05),可以促進乙基酯的形成,改善發酵風味。因此,乳酸菌發酵有助于形成復雜的風味成分,如氨基酸、維生素和有機酸,這些成分改善了產品的風味、滋味和其他特性[13]。
實驗室[14]前期篩選得到的PRO-MIX5(木糖葡萄球菌、清酒乳桿菌、類植物乳桿菌)能夠有效抑制紅腸和風干腸中N-亞硝胺(NAs)的形成,其抑制機理主要是其菌體碎片即微生物亞硝化抑制劑(MNI)的作用,且0.05%的MNI對紅腸和培根中的NAs有較好的抑制效果,并能提高產品的感官品質。目前,關于MNI在風干腸中的應用尤其是MNI與混合發酵劑協同作用對風干腸風味方面的研究尚未見報道。研究擬對風干腸的工藝參數進行調整,提高發酵溫度為30 ℃,降低風干成熟過程的溫度(14~16 ℃);將前期優選出的能提高風干腸安全品質的PRO-MIX發酵劑、發酵牛骨調味基料和復配抗氧化劑(FBFA)以及MNI應用于風干腸加工中,探討3種抑制劑尤其是MNI對風干腸成品風味物質形成的影響,旨在為MNI在風干腸中的開發應用提供依據。
1 材料與方法
1.1 材料與儀器
冷卻排酸成熟24 h的豬后腿肉、豬肥膘:天津二商迎賓肉類食品有限公司;
食鹽、白沙糖、曲酒、味精、醬油:市售;
茶多酚、迷迭香:豫中生物科技有限公司;
維生素E、異抗壞血酸鈉:蘇州佰億鑫生物科技有限公司;
PRO-MIX5復合商業發酵劑(木糖葡萄球菌、清酒乳桿菌、類植物乳桿菌):活菌數1011CFU/g,封裝于無菌玻璃瓶中凍藏(-20 ℃),意大利薩科公司;
人工膠原蛋白腸衣:牛二層皮提取、孔徑30 mm,神冠控股(集團)有限公司;
2-甲基3-庚酮:分析純,美國Supelco公司;
氣相色譜—質普聯用儀:GCMS-QP2010Plus型,日本島津公司;
超快速氣相電子鼻:HeraclesII型,法國Alpha M.O.S公司;
電子舌:TS-5000Z型,日本INSENT公司;
氨基酸分析儀:S-43000型,德國Sykam公司;
恒溫恒濕箱:CLIMACELL型,艾力特國際貿易有限公司;
真空攪拌機:BVBJ-30F型,浙江嘉興艾博實業有限公司。
1.2 方法
1.2.1 FBFA制備 參照樊曉盼等[15]的方法制備FBF,復合抗氧化劑(CA)添加量:茶多酚、迷迭香、VE和抗壞血酸鈉添加量60.14,60.11,60.00,60.00 mg/kg[16]。
1.2.2 MNI制備 參照李秀明等[17]的方法。
1.2.3 風干腸制作流程
原料肉→預處理→腌制→拌餡→罐腸→風干成熟→成品
操作要點:
(1) 腌制:將豬后腿肉剔除筋膜、脂肪,切成8 cm×5 cm×3 cm的方塊,用絞肉機攪碎(篩板孔徑8 mm),放入真空攪拌機中,加入占肉總質量(風干腸肥瘦質量比1∶9)1.8%的食鹽、0.01%的亞硝酸鈉(預先用少量水溶解)和抗壞血酸鈉0.55 g/kg,真空攪拌5 min,取出后放入不銹鋼盆中,緊貼肉表面蓋一層保鮮膜,4 ℃腌制24 h。
(2) 拌餡:將腌制好的肉倒入真空攪拌機中,依次加入4%糖、1.5%曲酒、0.2%味精、0.3%生抽、10%水、真空攪拌8 min。
(3) 灌腸:將制好的肉餡灌入膠原蛋白腸衣中,結扎(每節13~15 cm)、排氣。
(4) 發酵:將罐制好的肉餡于30 ℃,RH 90%,風速100%的恒溫恒濕培養箱中進行發酵,使其pH降至接近5.4后,終止發酵。
(5) 成熟:按表1調節恒溫恒濕培養箱的溫度、RH和風速。

表1 風干腸工藝參數Table 1 Process parameters of air-dried sausage
1.2.4 試驗設計方案
(1) CK組:按風干腸的基礎配方和工藝制作。
(2) MNI組:按風干腸的基礎配方和工藝制作,按肉重的0.05%加入由PRO-MIX5制備的菌體碎片(微生物亞硝化抑制劑,MNI)。
(3) MNIP組:按風干腸的基礎配方和工藝制作,按肉重的0.05%加入MNI,接入20 g/100 kg的PRO-MIX5商業復合菌(木糖葡萄球菌、清酒乳桿菌、類植物乳桿菌)。
(4) FBFAP組:按風干腸的基礎配方和工藝制作,按肉重的2%加入FBF和CA,接入20 g/100 kg的PRO-MIX5商業復合菌(糖葡萄球菌、清酒乳桿菌、類植物乳桿菌)。
1.2.5 指標測定
(1) 游離氨基酸含量:參照牛樹彬等[18]的方法,按式(1)計算單個游離氨基酸的滋味強度值(TAV)。
(1)
式中:
TAV——滋味強度值;
C1——呈味化合物含量,mg/100 g;
C2——呈味化合物的滋味閾值,mg/100 g。
(2) HeraclesII 超快速電子鼻分析:參照陳援援等[19]的方法。
(3) 電子舌分析:參照陳援援等[19]的方法。
(4) 揮發性風味物質(GC-MS)檢測:參照雷虹[20]的方法。
(5) 氣味活度值(OAV) 計算:參照張凱華等[21]的方法。
1.3 數據處理
采用Excel 2010軟件計算平均值和標準差,用SPSS 19.0軟件進行顯著性分析,Origin 2018軟件作圖。顯著性水平P<0.05。
2 結果與分析
2.1 風干腸成品揮發性風味物質PCA分析
由圖1可知,主成分1和主成分2的方差貢獻率分別為94.127%,4.495%,累計方差貢獻率為98.622%,表明這兩個主成分足以代表樣品的大部分信息。4組風干腸成品之間不存在交叉和重合,說明CK、MNI、MNIP和FBFAP組風干腸中揮發性成分的種類和含量存在差異。其中MNI組和MNIP組之間的相對距離較近,說明兩組樣品之間存在相同的揮發性風味物質,可能是兩者的特征揮發性風味物質比較突出所以將兩者區分開來。

圖1 基于氣相電子鼻檢測風干腸成品揮發性 風味物質的PCA分析Figure 1 PCA analysis of volatile flavor substances in air-dried sausage products based on gas phase electronic nose detection
2.2 風干腸的有效味覺
由圖2可知,主成分1和主成分2的方差貢獻率分別為98.873%,0.921%,累積方差貢獻率為99.794%。4組風干腸成品在味覺上存在一定差異,但MNI組和MNIP組在8個味覺指標上比較接近,FBFAP組和CK組的比較接近。酸味的無味點為-13,苦味回味、澀味回味和豐富性的無味點為0,4組風干腸成品的酸味、苦味回味、澀味回味和豐富性在傳感器上的響應值低于無味點,說明4組樣品在這4個味覺指標的滋味特征不明顯;咸味的無位點為-6,苦味、澀味和鮮味的無味點為0,4組樣品的苦味、澀味、鮮味和咸味在傳感器上的響應值大于無味點,說明這4個味覺指標是4組風干腸成品有效且重要的味覺指標。MNI組和MNIP組的鮮味響應值最高,苦味和咸味響應值低于CK組,說明加入MNI、MNIP能降低苦味和咸味,提高風干腸的鮮味。

圖2 基于電子舌檢測風干腸成品的PCA和有效味覺指標分析Figure 2 PCA and effective taste index analysis of air-dried sausage based on electronic tongue
2.3 風干腸成品的游離氨基酸組成
由表2可知,谷氨酸、丙氨酸、牛磺酸和肌肽是4組風干腸中的主要游離氨基酸,谷氨酸和天冬氨酸呈鮮味,在MNI組和MNIP組中的含量顯著高于CK組和FBFAP組,說明加入MNI和MNIP有利于風干腸鮮味氨基酸的形成。牛磺酸是一種具有顯著抗氧化活性的化合物[22],在3個處理組中的含量明顯高于CK組(P<0.05)。甜味氨基酸總量在FBFAP組中最高,苦味氨基酸總量在CK組中最高,說明加入FBFAP能改善終產品的甜味、抑制苦味。研究[23]表明,蛋白質過度水解會給肉制品帶來苦味或金屬味。4組風干腸游離氨基酸總量大小為MNIP組>CK組>FBFAP組>MNI組,FBFAP組中的游離氨基酸總量低于CK組(P<0.05),與Hu等[24]的結果不一致,可能是由于產品配方、加工條件和微生物種群不同。4組風干腸游離氨基酸總量不同,是因為肌肉和細菌來源的酶活性受加工參數的影響,如溫度、相對濕度、發酵時間和pH值變化等。在前期試驗的基礎上改變工藝條件,新增了一個較高溫度30 ℃的發酵階段,MNI組和MNIP組的產酸速度快,分別在發酵第15.0,12.5 h結束發酵,CK組和FBFAP組分別在發酵第22,21 h結束發酵,由于發酵時間、肉餡pH值和添加物不同,4組風干腸的游離氨基酸含量有差異,此外,也有可能是游離氨基酸參與風味物質的形成而減少。綜上,加入MNIP能縮短風干腸的發酵時間,促進風干腸鮮味氨基酸的生成和提高游離氨基酸總量。

表2 風干腸成品的游離氨基酸組成?Table 2 Free amino acid composition of 4 groups of air-dried sausages
2.4 風干腸成品游離氨基酸的味覺特征及TAV值
由表3可知,4組風干腸中TAV值>1的氨基酸有4種,包括谷氨酸、丙氨酸、纈氨酸和組氨酸。谷氨酸的TAV值最高,對風干腸滋味貢獻度最大。其次是丙氨酸、纈氨酸和組氨酸,丙氨酸為甜味氨基酸,纈氨酸和組氨酸為苦味氨基酸,TAV較小,對風干腸滋味貢獻度較小,所以風干腸主要呈鮮甜味。MNIP組中谷氨酸的TAV值最高,其次是MNI組,由于MNIP組中對鮮味有貢獻的游離氨基酸含量均高于其他3組,因此MNIP組發酵的風干腸鮮味更強烈,同時說明在接種有發酵劑PRO-MIX5時,添加MNI促進風干腸中鮮味強度增加的效果優于FBFA。

表3 風干腸成品游離氨基酸的味覺特征及TAV值Table 3 Taste characteristics and TAV values of free amino acids in 4 groups of air-dried sausages
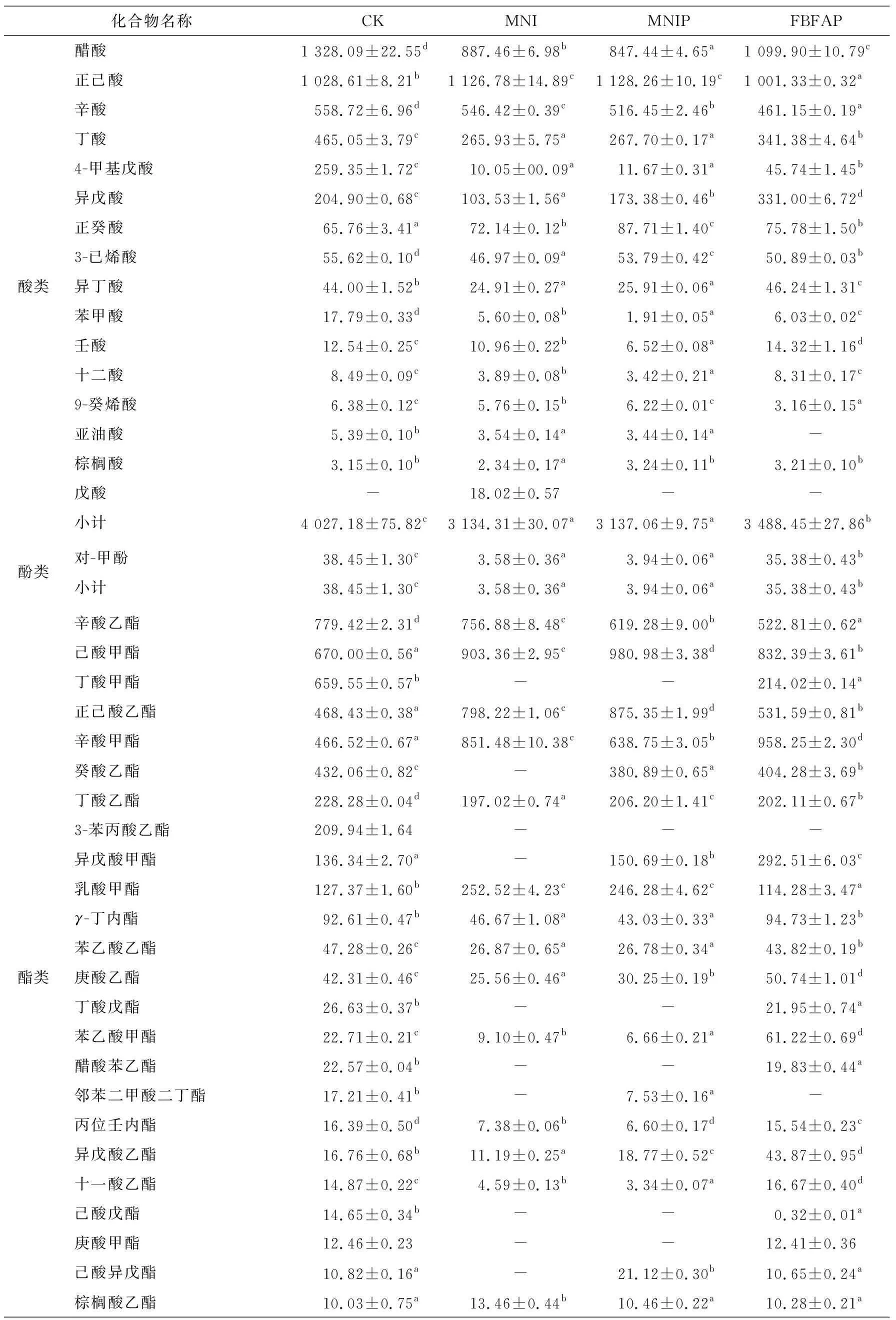
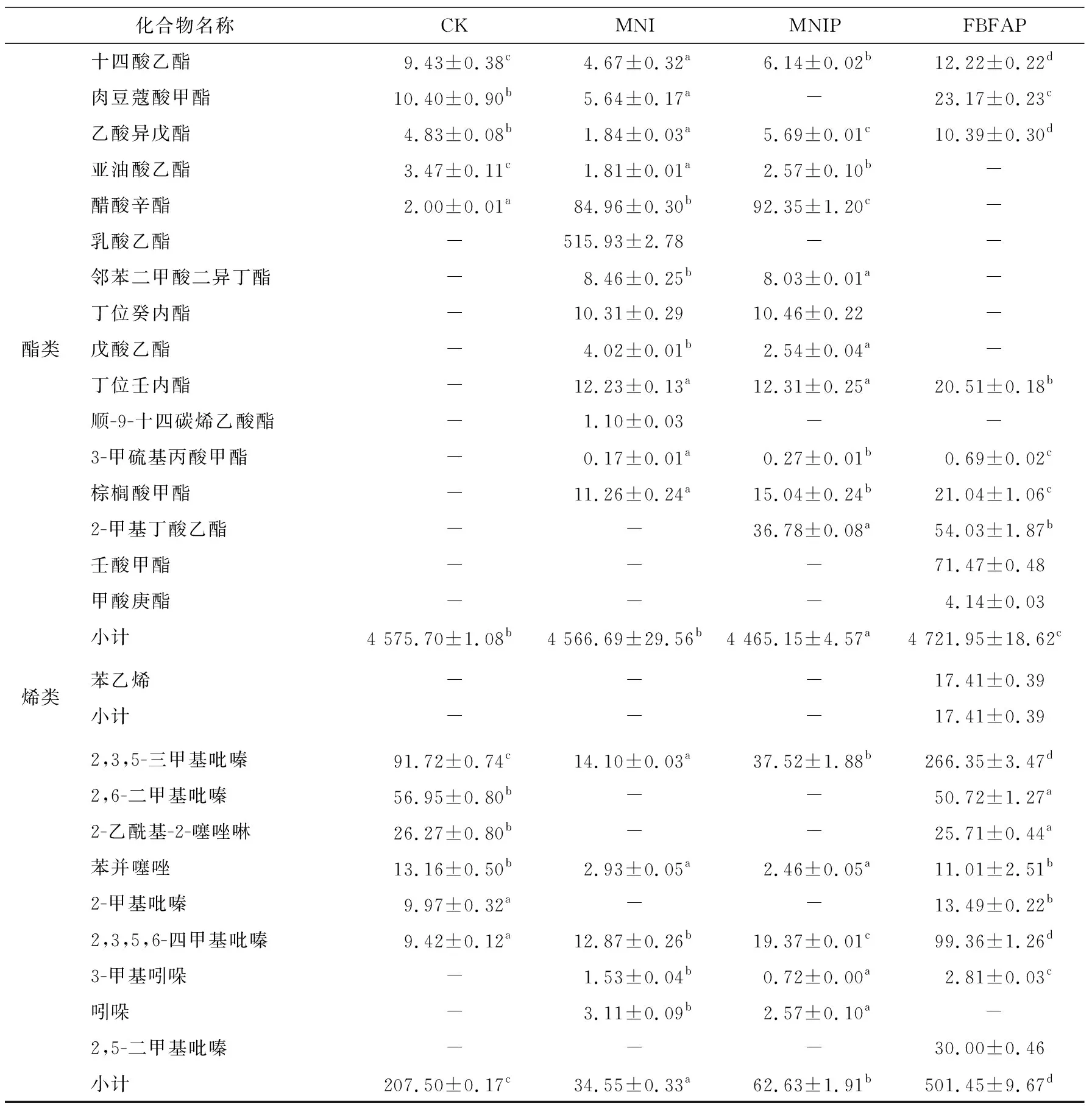
2.5 基于GC-MS分析風干腸成品揮發性風味物質
由表4可知,酯類、醇類、酸類、醛類和雜環化合物類是4組風干腸主要的風味成分。MNI、MNIP和FBFAP組中檢出的風味物質總量顯著高于CK組(P<0.05),主要歸因于發酵過程中發生的生化反應,如碳水化合物發酵、氨基酸代謝、蛋白質氧化、脂質自氧化、脂質β-氧化和酯化等[25-26]強度不一致。

表4 4組風干腸成品揮發性風味物質相對含量?Table 4 Relative mass concentration of volatile flavor substances in 4 groups of air-dried sausage products μg/kg
續表4
續表4
2.5.1 醛類物質 一般來說,脂質自動氧化會產生醛類物質,由于其低閾值,對風干腸整體風味貢獻較大[9]。4組風干腸中醛類物質總量大小為MNI組>MNIP組>FBFAP組>CK組,說明加入MNI、MNIP能促進風干腸中醛類風味物質的形成與積累。4組樣品中均檢測到苯甲醛和苯乙醛,主要是由細菌代謝芳香族氨基酸產生[1]。醛類物質中,正己醛含量最高且在4組樣品中差異顯著(P<0.05),可能是由于乳酸菌中相關酶的活力不同。反式-2-辛烯醛、正己醛、反式-2,4-壬二烯醛、反式-2,4-庚二烯醛、反-2-癸烯醛、反式-2,4-癸二烯醛、2-十一烯醛、反式-2-庚烯醛和正辛醛是MNI組和MNIP組的特征醛類物質。線性醛類主要來自不飽和脂肪酸的氧化降解,如油酸、亞油酸、亞麻酸和花生四烯酸[23],而支鏈醛的主要形成途徑為氧化脫氨脫羧,可能通過Strecker降解產生。
2.5.2 醇類、酮類物質 醇的產生有多種方式,如氨基酸代謝、脂質氧化和甲基酮還原[27]。乙醇是4組風干腸中含量最豐富的化合物,在MNI組和MNIP組中含量最高,可能與乳酸菌主導的碳水化合物發酵、氨基酸代謝和風干腸制作過程中加入的曲酒有關,Lin等[28]研究發現,在乳酸菌的氨基酸代謝過程中,其中一條氨基酸途徑與兩種脫氫酶有關,即α-酮酸脫氫酶和羥基酸脫氫酶,具有抗氧化能力的乳酸菌菌株可以增強氨基酸的降解,并通過還原反應促進醛生成更多的醇。還可能是加入的MNI具有一定的抗氧化性,MNI是PRO-MIX5發酵劑的細胞破碎物,該細胞破碎物含有多種生物活性成分,如胞外多糖。研究[29]發現,乳酸菌是優良的胞外多糖產生菌。植物乳桿菌c88胞外多糖具有顯著清除活性氧效果,其他植物乳桿菌的胞外多糖在高濃度下對DPPH自由基和超氧自由基有較強的清除能力[30-31]。Nehal等[32]研究發現,乳酸菌產生的胞外多糖具有抗氧化作用,且抗氧化效率與胞外多糖濃度有關。1-辛烯-3-醇具有蘑菇味和花香,是發酵肉制品中常見的不飽和醇[33],4組風干腸中均有檢出,其在MNI組和MNIP組中的相對質量濃度最高。正庚醇、正丁醇可通過風干腸中的乳酸菌和酵母介導的某些氨基酸轉化而得[34]。4組風干腸中檢測到含量較高的3-羥基-2-丁酮,尤其是在MNIP組中最高,丙酮酸可通過一系列代謝反應轉化為2,3-丁二酮,而2,3-丁二酮將通過各種還原酶轉化為3-羥基-2-丁酮[35],Chen等[12]研究發現接種清酒乳桿菌會增加發酵香腸中3-羥基-2-丁酮含量。
2.5.3 酸類物質 風干腸發酵成熟過程中產生的有機酸可增強對香氣的感知[36]。4組風干腸中共鑒定出16種酸類物質,其中含量較高有醋酸、正己酸、辛酸、丁酸、4-甲基戊酸、異戊酸、正癸酸、3-已烯酸和異丁酸,醋酸可以由乳酸菌、葡萄球菌的脂肪酸氧化和丙氨酸分解代謝形成,丁酸可能來自于脂質氧化或纈氨酸脫氨[37]。CK組中酸類物質總量顯著高于其他3個處理組(P<0.05),FBFAP組次之,MNI組和MNIP組中酸類物質總量差異不顯著。Lin等[15]研究表明,乳酸菌代謝產生的醇和有機酸在酯酶和烷基轉移酶的作用下生成相應的酯,進而使MNI、MNIP和FBFAP組中酸類物質含量降低。
2.5.4 酯類物質 酯類是4組風干腸中含量最豐富的揮發性風味物質,共檢出40種酯類。FBFAP組中酯類物質總量最高,其生成取決于醇和酸的存在情況,以及葡萄球菌的酯化酶活性,特別是通過乙酰輔酶A或酰基輔酶A與酒精的酯化反應生成的酯,使發酵肉制品具有水果味、甜味和花香的風味[38]。辛酸乙酯、己酸甲酯、正己酸乙酯、辛酸甲酯、丁酸乙酯和乳酸甲酯是4組風干腸中最主要的酯類物質,短鏈酸形成的酯有水果味,這有助于形成風干腸的特殊風味。具有清酒曲香味和菠蘿香味的正己酸乙酯、己酸甲酯和乳酸甲酯在MNI組和MNIP組中的相對含量最高,丁酸甲酯在CK組和FBFAP組中的含量較高,在MNI組和MNIP組中未檢出,說明加入的MNI、MNIP和FBFAP能改變風干腸中酯類物質的含量。
2.5.5 雜環化合物類 肉在發酵過程中會形成雜環類化合物包括吡嗪、呋喃(杏仁味和甜味)、噻唑和吲哚類。美拉德反應產生的2-甲基吡嗪類物質,即使在較低的濃度水平,也有利于風干腸風味的形成。4組風干腸中檢測出9種雜環類化合物,FBFAP組中的雜環類化合物相對含量最高,CK組次之,MNI組和MNIP組較低。研究[23]表明,接種菌株或風干腸中的原始微生物對美拉德反應底物(還原糖和氨基酸)的分解代謝可能導致這些化合物的形成或降解,但是關于其形成或降解機制需進一步深入研究。
2.6 風干腸成品揮發性風味物質OAV值
由表5可知,4組風干腸中有13種揮發性化合物的OAV>1,分別為1-辛烯-3-醇、芳樟醇、反式-2-辛烯醛、正己醛、癸醛、反式-2,4-壬二烯醛、正辛醛、辛酸乙酯、正己酸乙酯、癸酸乙酯、丁酸乙酯、庚酸乙酯和異戊酸乙酯,是風干腸發酵氣味的主要來源。MNI組和MNIP組的醇類(1-辛烯-3-醇)OAV值、醛類如反式-2-辛烯醛、正己醛、癸醛、反式-2,4-壬二烯醛和正辛醛的OAV值遠高于CK組,說明加入MNI和MNIP有利于風干腸中醇類物質和醛類物質的呈味。正己酸乙酯和異戊酸乙酯在MNIP組中的OAV值最大,對該組風味貢獻最大,芳樟醇僅在MNI組中檢出,且OAV>1,說明芳樟醇是MNI組的特征風味物質。癸醛和異戊酸乙酯在FBFAP組中的OAV>1,且遠大于其余3組,說明癸醛和異戊酸乙酯是FBFAP組的特征風味物質。短鏈脂肪酸在4組風干腸中相對含量均較高,但由于其嗅覺閾值高,對4組風干腸的風味貢獻不大。

表5 4組風干腸成品揮發性風味物質OAV值Table 5 OAV value of volatile flavor substances in 4 groups of air-dried sausage products
3 結論
利用電子鼻和電子舌分別對4組風干腸的揮發性化合物和滋味進行了分析。結果表明,4組風干腸的風味和滋味均不同,且能將其明顯區分開,相比較而言,微生物亞硝化抑制劑組和微生物亞硝化抑制劑與PRO-MIX5商業發酵劑協同組風味和滋味特征相似,發酵牛骨調味基料和復配抗氧化劑與PRO-MIX5商業發酵劑協同組與空白對照組的氣味和滋味特征相似。鮮味、咸味、苦味和澀味是4組風干腸中有效且重要的味覺指標,加入微生物亞硝化抑制劑和微生物亞硝化抑制劑與PRO-MIX5商業發酵劑協同能提高風干腸的鮮味,降低咸味和苦味。谷氨酸、丙氨酸、牛磺酸和肌肽是4組風干腸中的主要游離氨基酸,加入微生物亞硝化抑制劑與PRO-MIX5商業發酵劑協同能縮短風干腸的發酵時間,促進風干腸鮮味氨基酸的生成和提高游離氨基酸總量;谷氨酸和丙氨酸這兩種呈鮮味和甜味的氨基酸對風干腸的滋味貢獻最大,4組風干腸主要呈鮮甜味。4組風干腸中共鑒定出105種揮發性風味物質,其中正己酸乙酯和異戊酸乙酯是微生物亞硝化抑制劑與PRO-MIX5商業發酵劑協同組的特征風味物質,芳樟醇是微生物亞硝化抑制劑組的特征風味物質,癸醛和異戊酸乙酯是發酵牛骨調味基料和復配抗氧化劑與PRO-MIX5商業發酵劑協同組的特征風味物質。單獨添加微生物亞硝化抑制劑或微生物亞硝化抑制劑與PRO-MIX5商業發酵劑協同均能促進風干腸風味的形成。乳酸菌和微生物亞硝化抑制劑能促進風干腸中風味物質的形成,關于其產生醛類、酸類、醇類和酯類等風味物質的代謝途徑和影響機制仍有待進一步研究。
