小相嶺山系非飛行小型獸類物種多樣性海拔分布格局
2022-07-28 08:26:30劉芝秀周恩華張勘王大勇胡杰李艷紅吉晟男
四川動物 2022年4期
劉芝秀,周恩華,張勘,王大勇,胡杰*,李艷紅*,吉晟男
(1.西華師范大學西南野生動植物資源保護教育部重點實驗室,四川 南充 637009;2.四川栗子坪國家級自然保護區管理局,四川 石棉 625400;3.四川冶勒自然保護區管理處,四川 冕寧 615600;4.中國環境科學研究院,北京 100012)
小型獸類因種類多、分布廣、適應能力強,在生態系統中占據著重要地位(秦嶺等,2007),同時在維持食物鏈、食物網、營養循環以及生態系統的結構和功能方面也發揮著重要的作用(Liu,2009)。另外,小型獸類遷移性小,對環境變化敏感且種群數量穩定(Umetsu,2008),其群落結構的變化不僅可以反映生境變化,還可以反映人類活動的干擾程度,是評估環境質量的良好實驗對象(傅琰華等,2015)。非飛行小型獸類(簡稱小型獸類)主要是指除翼手類外的所有體型較小、體質量較輕的哺乳動物。本文研究的對象主要是勞亞食蟲目Eulipotyphla、嚙齒目Rodentia和兔形目Lagomorpha的物種。
物種多樣性包括α、β和γ 3個不同的空間尺度,相互影響、相互作用、不可分割。在對物種多樣性海拔空間分布格局的研究上,α、β和γ多樣性出現并駕齊驅的趨勢(權壽瑛等,2009;吳永杰等,2012;Krasnov,2018)。全球物種豐富度海拔梯度分布格局主要包括4種模式,即單調遞減模式、前峰模式、駝峰模式和單調遞增模式(Rahbek,2005)。在小型獸類中,前峰模式和駝峰模式更普遍(Rahbek,2005;McCain,2006)。我國大多數學者對小型獸類海拔梯度格局的研究也表明,物種豐富度在中海拔地區最高(權壽瑛等,2010;王作波等,2020)。相鄰海拔之間的物種更替地理格局則表明,多峰和不規則模式最普遍(Tang,2012;Wang,2012)。目前,對小型獸類多樣性海拔分布格局形成機制的探討尚未有定論,因此需要在更多的地區開展相關研究。
小相嶺山系位于全球36個生物多樣性熱點區域之一的西南山地(Habel,2019),區域內植被類型豐富、動植物種類繁多,在生物多樣性保護和維持方面具有重要的研究價值。迄今為止,該區域內物種多樣性的相關研究著重于大中型獸類和鳥類(李健威等,2020),小型獸類僅有簡單的描述性報道(張君,胡錦矗,2004;李艷紅等,2007)。為此,在小相嶺山系開展有關小型獸類的研究,一方面可以進一步豐富該區域小型獸類多樣性的基礎信息,另一方面,可以探討小相嶺山系小型獸類的海拔梯度分布格局,進而為該區域小型獸類生物多樣性的保護和管理提供科學依據。
1 研究地區和研究方法
1.1 研究區概況
小相嶺山系屬大雪山支脈,面積約11 500 km,介于石棉、冕寧、越西、喜德和九龍5縣交界處(圖1),境內地形崎嶇,山嶺重疊,地勢西高東低,地貌切割破碎強烈,以中山和高山峽谷為主,南北長約147 km,東西寬約107 km,總體呈南北走向,且北高南低(冉江洪等,2004;胡錦矗,吳攀文,2007)。小相嶺山系由焦頂山、鏵頭尖、窩洛幾峨等山峰組成,主山脊海拔大多在4 000 m以上,最高峰為則爾山,海拔5 299 m,最低海拔處于大洪村板栗樹溝(1 330 m),海拔平均相對高差2 000 m以上(Hu,2010)。山系四周由于高山屏障常產生強烈的焚風效應,且有明顯的干濕季節變化,屬于以亞熱帶季風為基帶的山地氣候,冬春多風,夏秋多雨,光照少,年均日照率為30%左右。由于地勢原因,“雨屏效應”顯著,年均降水量800~1 250 mm,年均氣溫11.7~14.4℃,年均相對濕度87.9%,垂直變化明顯(青菁等,2016)。

圖1 小相嶺山系樣方布設Fig.1 Sample sites in the Xiaoxiangling Mountains
小相嶺山系植被類型豐富多樣,發育形成了完整的植被垂直帶譜:海拔1 330~2 400 m為常綠闊葉林,山楠、曼青岡、巴東櫟、卵葉釣樟等為優勢種;海拔2 400~2 700 m為落葉闊葉林,野核桃細葉青岡槲櫟、華西楓楊、大葉楊等為優勢種;海拔2 500~2 700 m為針闊葉混交林,扇葉槭、紅樺、鐵杉、麥吊云杉等為優勢種;海拔2 700~3 700 m為亞高山針葉林,鐵杉、麥吊云杉、川滇冷杉、冷杉等為優勢種;海拔2 800~3 700 m為亞高山灌叢,大白杜鵑、小葉栒子、四川嵩草等為優勢種;海拔3 700~4 400 m為高山灌叢草甸,凝毛杜鵑、金露梅、峨眉薔薇、高山嵩草、高原毛茛等為優勢種;海拔4 400 m以上為高山流石灘稀疏植被,風毛菊、水母雪蓮花、梭砂貝母等為優勢種(劉守江等,2011)。
1.2 試驗設計
2019年7—8月和2020年7—9月,根據小相嶺區域的生境類型和地形地貌特征等因素,采用夾日法對該區域內的小型獸類進行野外調查。選取海拔1 300~4 100 m為研究區域,每400 m設置樣方,在7個連續海拔段共設置53個樣方。樣方間距大于100 m,并盡可能覆蓋所有生境類型。樣方大小為100 m×100 m,夾線行距25~30 m,夾距3~5 m,每個樣方放置66~88個鼠夾。以新鮮花生米為誘餌,每日下午放置,翌日上午收夾,連捕3 d。收夾時,收集捕獲動物并補放誘餌,同時對捕獲到標本的鼠夾進行清洗和烘干。將捕獲的標本進行編號、稱重和常規測量(包括頭體長、尾長、吻長、耳長和后足長),鑒定性別和年齡段并記錄數據。所有標本用99%無水乙醇浸制,帶回實驗室鑒定。
1.3 數據處理
小型獸類物種鑒定主要參考《中國獸類野外手冊》(Smith,解焱,2009)和《四川資源動物志(第二卷)》(胡錦矗,王酋之,1984);分類系統參考《中國哺乳動物多樣性(第2版)》(蔣志剛等,2017)。
通過物種累積曲線判斷野外調查是否滿足取樣需求(Ugland,2003)。
物種豐富度用觀察到的物種數表示,插值物種豐富度指的是如果一個物種在較低和較高海拔都有記錄,那么就認為該物種在這2個海拔之間的所有區域都有分布。Chao2豐富度作為另外一種矯正指數,其計算原理是將實際捕獲的物種數通過非參數的統計方法,估算出總的物種數量(Colwell&Coddington,1994)。物種多度為每個樣方的個體數之和,相對多度由物種捕獲率表示。
群落多樣性指數采用Shannon-Wiener多樣性指數(′)和Simpson多樣性指數();優勢種判斷采用Berger-Parker優勢度指數();群落相似性采用Chao-S?rensen相似性指數()。利用一元線性方程評估隨海拔距離增加的衰減率,斜率的絕對值表示衰減率,值越大表示物種相似性隨海拔距離的增加衰減越快,說明該地區β多樣性越高(Soininen,2007)。

式中,為物種數,P為種的個體數占全部個體數的比例,N為優勢種的個體數,為群落中全部物種的個體數,分別為2個群落各自的物種數,為2個群落共有物種數。
所有數據利用Excel 2016、SPSS 21.0和EstimateS 9.0進行處理分析。
2 結果
2.1 物種組成
累計布設11 902個夾日,捕獲小型獸類標本317只,隸屬于3目7科15屬23種,總捕獲率為2.7%。其中,勞亞食蟲目3科(鼩鼱科Soricidea、鼴科Talpidae、猬科Erinaceidae)5屬6種;兔形目鼠兔科Ochotonidae 1屬2種;嚙齒目3科(鼠科Muridae、松鼠科Sciuridae、倉鼠科Cricetidae)9屬15種。嚙齒目數量最多,有225只,占總捕獲量的71.0%;勞亞食蟲目和兔形目較少,分別占18.3%和10.7%。在物種組成上,優勢種為康定絨鼠(18.3%)、中華姬鼠(12.9%)和高山姬鼠(10.1%)。不同物種具有不同的海拔分布區:分布在2 000 m及以上的4種,分別是高山姬鼠、中華姬鼠、大耳姬鼠和川西白腹鼠;其次是分布在1 500 m左右的微尾鼩、小紋背鼩鼱、康定絨鼠和藏鼠兔(表1)。以上物種為寬域分布物種(海拔分布范圍超過研究區域的一半),其余物種為狹域分布。

表1 小型獸類物種組成及海拔分布Table 1 Species composition and elevational distribution of small mammals
結合文獻資料(張君,胡錦矗,2004;李艷紅等,2007;李健威等,2020)和調查訪問情況,小相嶺山系記錄小型獸類45種,隸屬于3目8科28屬。其中,勞亞食蟲目3科12屬16種,占總物種數的35.6%;嚙齒目4科15屬27種,占60.0%;兔形目1科1屬2種,占4.4%。中國特有種16種,占35.6%。列入《中國生物多樣性紅色名錄(第一卷哺乳動物)》(蔣志剛,2021)的瀕危(EN)物種1種,即滇攀鼠;易危(VU)物種2種,為寬齒鼴和峨眉鼩鼴;近危(NT)物種4種,為白尾鼴、紋背鼩鼱、中華絨鼠和康定絨鼠。新增分布記錄物種1種,即滇攀鼠。
2.2 物種豐富度取樣評估
在捕獲了15種物種后,物種累積曲線由直線上升變為舒緩上升,逐漸成一漸近線(圖2),說明本次野外調查取樣較為充分,可以滿足后續的處理分析需要。

圖2 物種累積曲線Fig.2 Species accumulation curves
2.3 物種多樣性海拔分布格局
本研究使用3種不同的物種豐富度指數驗證小相嶺山系小型獸類的海拔分布格局,均得到一致的研究結果,即物種豐富度在海拔2 500~2 900 m段最高。實際物種豐富度和插值物種豐富度的分布格局為單峰型,且變化趨勢較為一致,都在達到峰值后,呈遞減趨勢。Chao2豐富度的海拔分布格局為雙峰型,除了在海拔2 500~2 900 m段出現峰值外,在3 300~3 700 m也出現了峰值。另外,捕獲率作為反映群落物種相對多度的指標,在中低海拔段變化明顯,呈“W”形,隨后在高海拔段緩慢上升,最高點出現在3 700~4 100 m處(圖3)。

圖3 小相嶺山系的物種豐富度指數和物種多度海拔分布格局Fig.3 Elevational distribution patterns of species richness indices and species abundance in the Xiaoxiangling Mountains
小相嶺區域小型獸類的生物多樣性指數(Shannon-Wiener和Simpson多樣性指數)最大峰值出現在中海拔段,即2 500~2 900 m處,與物種豐富度海拔分布格局相似。Chao-S?rensen相似性指數結果表明,低海拔區域(1 700~2 100 m vs.2 100~2 500 m)和高海拔區域(3 300~3 700 m vs.3 700~4 100 m)的物種相似性最高,達到0.93和0.95,而中海拔段的物種相似性較低。群落相似性隨海拔的衰減率表明,小相嶺山系整個海拔梯度小型獸類的物種更替隨海拔增加而明顯衰減(圖4)。
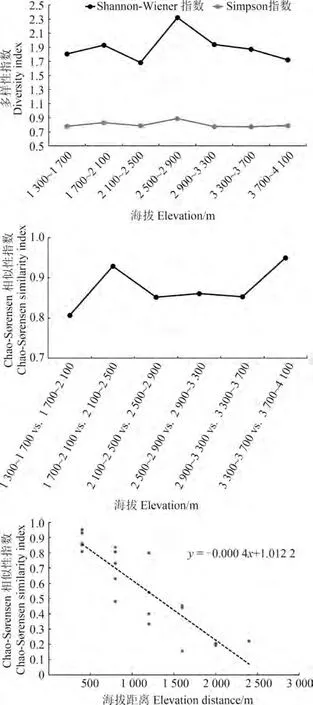
圖4 小相嶺山系物種多樣性海拔分布格局Fig.4 Elevational patterns of species diversity in the Xiaoxiangling Mountains
2.4 不同海拔小型獸類群落組成分析
在1 300~1 700 m內捕獲小型獸類9種,第一優勢種為高山姬鼠(0.40),第二優勢種為微尾鼩、北社鼠和川西白腹鼠(0.13);在1 700~2 100 m捕獲9種,第一優勢種為北社鼠(0.26),第二優勢種為微尾鼩(0.22),第三優勢種為中華姬鼠和高山姬鼠(0.15);在2 100~2 500 m捕獲7種,第一優勢種為中華姬鼠(0.32),第二優勢種為微尾鼩(0.24),第三優勢種為大耳姬鼠(0.21);在2 500~2 900 m捕獲13種,第一優勢種為中華姬鼠和高山姬鼠(0.17),第二優勢種為微尾鼩和大耳姬鼠(0.11);在2 900~3 300 m捕獲12種,第一優勢種為康定絨鼠(0.43),第二優勢種為中華姬鼠(0.13);在3 300~3 700 m捕獲10種,第一優勢種為康定絨鼠(0.43),第二優勢種為小紋背鼩鼱(0.12);在3 700~4 100 m捕獲8種,第一優勢種為高原松田鼠(0.35),第二優勢種為大耳姬鼠和藏鼠兔(0.17)(表2)。
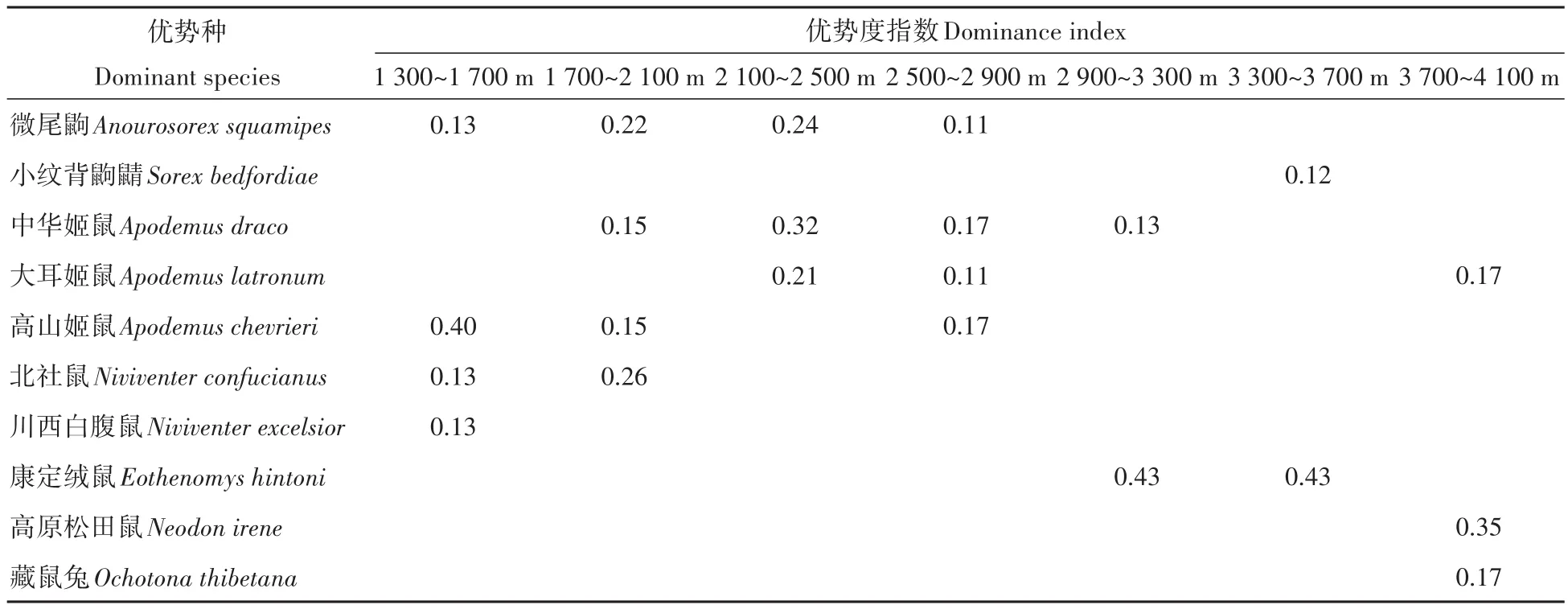
表2 不同海拔段小型獸類優勢種組成Table 2 Composition of dominant species of small mammals at different elevations
3 討論
3.1 物種組成
本次調查捕獲小型獸類23種,與以往調查相比(張君,胡錦矗,2004),有22種小型獸類未被調查到,但新增新分布物種滇攀鼠,核實原有的獸類名錄發現,部分物種的分布值得商榷,如根田鼠和黑唇鼠兔:根田鼠僅分布于新疆(劉少英等,2019),而黑唇鼠兔在四川僅分布于若爾蓋、甘孜、德格、石渠等川西高原地區(王酉之,胡錦矗,1999)。陳鵬(2012)關于姬鼠屬的研究表明,長尾姬鼠并非有效種,因此,此處的長尾姬鼠實為中華姬鼠。康定絨鼠原為西南絨鼠康定亞種,Liu等(2012)將該亞種提升為種。汪巧云等(2020)研究表明,喜馬拉雅水鼩僅分布在云南,而四川等地的實為利安德水鼩。此外,Liu等(2019)通過分子學實驗,對絨鼠屬的幾個亞種進行鑒定研究,認為石棉絨鼠和美姑絨鼠應為獨立有效種。因此,本文對這些物種進行了刪除或補充修訂。部分物種可能因其數量稀少或棲息于特殊生境內,未被捕獲,如林跳鼠s、蹼足鼩利安德水鼩等。
3.2 不同海拔的優勢種
小相嶺山系不同海拔的小型獸類優勢種主要由高山姬鼠、微尾鼩、北社鼠、川西白腹鼠、中華姬鼠、大耳姬鼠、康定絨鼠、小紋背鼩鼱、高原松田鼠和藏鼠兔組成。其中,中華姬鼠和大耳姬鼠在所有海拔段內均有分布。廣域分布種在不同海拔優勢種中的占比高達80%,說明其具有很強的繁殖、生存和適應能力(張云智等,2005)。對大相嶺山系小型獸類群落及生境的研究結果顯示,廣布種在群落優勢種中占比較大,且優勢種的種類(齊敦武,2005)與本研究結果較為相近,這表明大、小相嶺山系的小型獸類可能具有相似的群落特征。隨著海拔的升高,第一優勢種出現由高山姬鼠、北社鼠替換成中華姬鼠,再到康定絨鼠和高原松田鼠的現象,而第二與第三優勢種的變化無明顯規律。四川黑竹溝地區(岳先濤等,2020)和云南農耕區(陳星等,2019)小型獸類群落組成的海拔變化也表明,優勢種存在隨海拔升高而發生替換的現象。說明海拔的變化是影響小型獸類群落結構組成的重要因素,但地形、氣候和食物資源等其他因素的作用同樣不可忽視(胡一鳴等,2018)。
3.3 小型獸類物種多樣性海拔梯度分布格局
物種豐富度和物種多度作為反映物種生物多樣性最直接的2個指標,常被用于預測山地物種的分布格局,但由于其對常見種和稀有種的評價存在一定的缺陷,通常將Shannon-Wiener和Simpson多樣性指數綜合起來以彌補這一不足(栗忠飛,高吉喜,2018)。本研究中,小型獸類的物種數明顯在2 500~2 900 m最高,而物種相對多度的峰值則出現在2 900 m以上,二者的海拔梯度分布格局存在一定的差異。Shannon-Wiener和Simpson多樣性指數獨立于物種多度分布格局外,可以較為準確地評價常見種或稀有種的貢獻率,本研究中,這2種多樣性指數的海拔分布格局峰值都出現在海拔2 500~2 900 m段,因此將這2種多樣性指數與物種豐富度相結合,可以更好反映物種的垂直分布變化情況。對西南山地小型獸類物種豐富度海拔梯度分布格局的研究結果表明,該區域小型獸類物種豐富度的海拔格局存在明顯的空間變化,權壽瑛等(2009)發現橫斷山區100~5 000 m范圍內的小型獸類物種豐富度海拔分布格局為寬峰型;馬俊等(2010)發現螺髻山中海拔段的小型獸類物種多樣性最高;吳永杰等(2012)發現貢嘎山東坡小型獸類的物種豐富度在中海拔地區最高。本研究結果與上述結論一致,小相嶺山系的小型獸類物種豐富度在2 500~2 900 m段最高,符合中峰模式。這可能是由于西南山地2 500~3 000 m的中海拔地區位于最大降水帶,該海拔范圍內濕度大、植被類型多樣、食物豐富,相對于低海拔和高海拔結構簡單的生境,更利于小型獸類生存(張榮祖等,1997)。
3.4 小型獸類物種更替海拔梯度分布格局
對于物種β多樣性的分布格局,既往的研究主要通過相鄰海拔之間的物種更替和相似性距離衰減率2種不同空間尺度來分析:Mena和Vázquez-Domínguez(2005)發現鼠類的相鄰海拔間物種更替海拔格局分布模式均為多峰與不規則型;Wen等(2014)對橫斷山區小型獸類的物種更替地理格局的研究同樣表明,貢嘎山、唐家河和響古菁3條樣線的相鄰海拔間物種更替格局均不一致。本研究對小相嶺山系海拔1 300~4 100 m段小型獸類相鄰海拔間的物種更替格局分析發現,低海拔區域和高海拔區域的物種相似性最高,而中海拔段的物種相似性相對較低,說明中海拔段的物種更替更快、β多樣性更高。Rapoport法則預測,物種組成在高海拔地區的變化小于低海拔地區(Stevens,1992,1996),高海拔地區存在的物種分布區域重疊導致β多樣性隨海拔的升高而降低。在更大尺度上,不同海拔間的物種相似性隨海拔距離的增加而減小,表現為衰減率越大,β多樣性越高(Soininen,2007)。小相嶺山系小型獸類的相似性隨海拔距離的衰減率比西南山地其他地區的(Wen,2014)更低,這可能是由于小相嶺山系的群落優勢種主要為廣域分布種,擴散能力較強、物種聚集度較低,因而β多樣性相對較低。
本研究得到四川栗子坪國家級自然保護區管理局和四川冶勒自然保護區管理處的大力支持和協助;感謝張德軍、吳大鵬、李勇、陳紅、魏勇等人在野外采集工作中提供的幫助。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
