外源鈣緩解花生亞低磷光合障礙的機制
2022-07-18 08:59:34孫志宇劉欣悅張思威馬明珠易伯濤韓曉日劉軼飛
植物營養與肥料學報 2022年6期
孫志宇,劉欣悅,張思威,馬明珠,白 蕊,劉 歡,易伯濤,韓曉日,劉軼飛
(沈陽農業大學土地與環境學院/土肥高效利用國家工程研究中心,遼寧 沈陽 110866)
磷是地球上所有生物維持生命所必需的元素,也是植物中核酸、蛋白質、磷脂、ATP、ADP以及各種含磷酶的重要組分[1–3],因此,磷對作物生長發育和產量形成至關重要[4]。然而,磷限制是全世界作物生產中的主要制約因素之一,同時化學磷肥來自于磷酸鹽礦石,這也是一種極其有限的不可再生資源[5–6]。全球大約 70% 的耕地缺乏有效磷[7–8],但是磷肥當季利用率卻僅有10%左右,沒有被作物吸收利用的磷肥也引發了許多嚴重的環境污染問題[9]。喜磷喜鈣作物—花生 (ArachishypogaeaL.)是國際重要油料經濟作物,尤其在維持我國糧油安全上具有舉足輕重的地位[10–14]。特別是,基于后疫情時代“雙循環”新發展格局下、中國大豆產需缺口加大等背景下,我國明確了油料作物結構調整主要策略之一,就是多措并舉發展國內花生生產部分替代大豆進口[15]。因此,我國油料作物供給 (食用油)安全和國際新形勢新挑戰已然將花生產業綠色發展推到一個前所未有的國家戰略安全高度[13, 14, 16]。值得注意的是,我國花生中低產田面積大,尤其土壤有效磷多處于花生低磷 (<10 mg/kg)或者亞低磷 (17.5±7.5) mg/kg 脅迫狀態下[10, 17–19],這嚴重制約著花生產業的可持續發展。
光合作用是對鈣、磷養分變化最為敏感的代謝過程之一,同時也是花生生長發育和產量形成的基礎[10, 19–22]。鈣離子作為第二信使,在植物逆境適應性調節方面具有重要作用[23–25]。研究表明,增加外源鈣素供應可以改善大豆、木豆、豇豆、瓜爾豆等的磷素吸收利用效率[26–28]。另外,葉面噴施外源鈣也可促進低夜溫脅迫下花生根、莖、葉的生長和磷素積累,并且外源鈣激發了花生葉片的環式電子傳遞,提高了葉綠體ATP合酶活性[20]。鈣被認為參與了卡爾文本森循環(Calvin-Benson-Bassham,CBB)中關鍵酶的調節,鈣可介導CBB循環酶來調控光合作用[29]。磷素對光合器官進行光合碳同化作用是必不可少的,特別是花生中低產田有效磷多處于花生亞低磷脅迫狀態,顯著影響花生的葉片生長、相對葉綠素含量和光合物質積累,顯著減少了花生葉面積和比葉質量,并造成了花生葉片光合碳同化障礙和光抑制[10],而生產中發現,增施鈣肥可有效改善中低產田花生的生長發育、磷素利用效率和產量[14, 28, 30–32],但是其作用機制尚待探究。為此,本研究將主要探討外源鈣對亞低磷脅迫下花生生長發育和光合作用的影響,以期明確外源鈣緩解花生亞低磷光合障礙的關鍵生理機制,旨在為花生高產栽培及優化養分管理技術提供理論基礎和技術指導。
1 材料與方法
1.1 試驗設計
試驗于2020年在沈陽農業大學土地與環境學院人工氣候室內進行,以花生(ArachishypogaeaL)品種‘遼寧白沙’為試驗材料,溫湯浸種催芽,花生帶芽單粒播種于砂培盆(外徑14 cm、底徑9.2 cm、高12 cm)中,每兩天澆透全量Hoagland’s營養液,采用人工氣候室盆栽培養2周。人工氣候室溫度、濕度、光照的設置參數:晝間為[28℃、60%、600 μmol/(m2·s)],夜間為 [20℃、60%、0 μmol/(m2·s)],氣候室內 CO2濃度始終設置為 (400±5) μmol/mol。在第14天,每盆澆透ddH2O (1.5 L)洗鹽,砂培盆底孔(5個孔)自然外排。基于Hoagland’s營養液,本課題組前期篩選出了花生亞低磷脅迫濃度[–P,0.5 mmol/L,相當于花生中低產田土壤有效磷(15.5±3)mg/kg]及正常磷濃度營養液(CK,1 mmol/L,相當于花生高產田土壤有效磷≥30 mg/kg)[10],在不同磷濃度水平下使用KNO3和NH4NO3來平衡氮、鉀濃度。在第15天,開始進行花生中低產田亞低磷脅迫模擬試驗,選取人工氣候室內長勢整齊一致的盆栽花生幼苗100株,并平均分為四組,即:1) CK,正常磷營養液 (P 1 mmol/L)+噴清水對照;2) –P,亞低磷脅迫 (P 0.5 mmol)+噴清水;3) –P+Ca,亞低磷脅迫+葉面噴施 CaCl215 mmol/L;4)–P+TFP,亞低磷脅迫+鈣調蛋白CaM抑制劑—三氟啦嗪(trifluoperazine, TFP)。每天早7:00澆透CK或者–P營養液(多余營養液底孔自然排出),并且早8:30對應以上各個處理,進行葉面均勻噴施ddH2O、CaCl2和TFP等。花生中低產田亞低磷脅迫試驗處理10天,在亞低磷脅迫處理的第9天進行活體指標測定,在亞低磷脅迫處理的第10天進行破壞性取樣。
1.2 花生植株干物質重、葉面積、相對葉綠素含量及葉片氣體交換參數的測定
在亞低磷脅迫處理的第10天,使用便攜式葉面積儀 AM-300 (ADC BioScientific,UK)測定各處理花生單株葉面積,記錄參數,將植株洗凈后放入牛皮紙袋,于電熱恒溫鼓風干燥箱DHG-9204A (上海精宏)中105℃殺青30 min后,65℃下烘干至恒重,使用千分之一天平測量各處理花生植株干物質重,記錄參數。相對葉綠素含量采用葉綠素儀(SPAD-502 plus,日本)測定。
在亞低磷脅迫處理的第9天上午10:00,使用高級光合測量系統 GFS-3000 (Heinz Walz, Effeltrich,Germany)測量各處理花生倒三葉位置葉片的相關氣體交換參數。測定時參數設置為:葉室面積3 cm2,CO2濃度 400 μmol/mol,測定光強 600 μmol/(m2·s)。CO2濃度通過二氧化碳鋼瓶保持穩定。分別記錄各處理花生葉片的凈光合速率(net photosynthetic,Pn)、蒸騰速率(transpiration rate,Tr)、氣孔導度(stomatal conductance,Gs)、胞間 CO2濃度 (intercellular CO2concentration,Ci)。
1.3 慢速葉綠素熒光誘導動力學曲線和P700活性的測定
在亞低磷脅迫處理的第9天,使用調制葉綠素熒光儀 Dual-Pam 100 (Heinz Walz, Effeltrich, Germany)對花生倒三葉位置葉片的葉綠素熒光和P700活性進行測量。待測花生植株暗適應30 min后,選擇同時測量PSⅡ 和PSⅠ的Fluo+P700測量模式,分析模式選擇SP-Analysis,測量開始后,打開測量光,測量光很弱,小于 1 μmol/(m2·s),只激發色素的本底熒光但不足以引起任何的光合作用,記錄最小熒光(FO)。之后打開一次飽和脈沖,記錄最大熒光(Fm)。關閉飽和脈沖后,熒光迅速回到FO附近,之后打開光化光,期間以30 s為周期打開一次飽和脈沖,記錄最大熒光(Fm),待熒光穩定后記錄穩定熒光(Fs),300 s后熒光曲線處于穩態,關閉光化光,待熒光值穩定后記錄最小熒光(F′o),結束測量程序。測量的PSⅡ熒光參數包括:PSⅡ最大量子產量Fv/Fm、PSⅡ實際量子產量Y(Ⅱ)、PSⅡ非調節性能量耗散的量子產量Y (NO)、PSⅡ調節性能量耗散的量子產量Y (NPQ)。測量的PSⅠ參數包括:PSⅠ實際量子產量Y(Ⅰ)、由PSI供體側限制引發的非光化學能量耗散的量子產量Y(ND)、由PSⅠ受體側限制引發的非光化學能量耗散的量子產量Y(NA)。
1.4 P700氧化動力學曲線和P700+暗還原曲線的測定
在亞低磷脅迫處理的第9天,使用Dual-Pam 100 (Heinz Walz, Effeltrich, Germany)可以在遠紅光(FR)條件下,通過多周轉脈沖(MT)和單周轉脈沖(ST)所誘導形成峰的面積比來量化電子載體質體醌(PQ)庫的大小[33],關閉遠紅光后,P700+暗還原曲線下降斜率的快慢可以表征環式電子傳遞(cyclic electron flow,CEF)的速率[34]。待測花生幼苗暗適應30 min 后,遠紅光下照射 20 s,然后打開 ST,15 s后打開MT,曲線平穩后關閉遠紅光,此時曲線將迅速下降,20 s后關閉程序,測量結束記錄參數。使用Origin來計算MT和ST所形成峰的峰面積,之后計算MT峰與ST峰面積比值。將遠紅光關閉后P700信號下降曲線經標準化處理后,比較下降斜率。
1.5 P515信號測定
在亞低磷脅迫處理的第9天,使用Dual-Pam 100 (Heinz Walz, Effeltrich, Germany)的 P515/535 模塊測量550—515 nm波長的信號變化。使用Dual Pam v1.19軟件的自動化腳本在每次測量之前校準和平衡P515信號[35]。暗適應1 h后,記錄單周轉飽和脈沖誘導的P515信號變化情況,可以反映類囊體膜的完整性。在 630 μmol/(m2·s)光照 10 min 和暗適應4 min后,記錄單周轉飽和脈沖誘導的P515信號變化,可以反映葉綠體ATP合酶活性。暗適應12 h后550—515 nm信號的慢速暗光暗誘導曲線反映了膜電位和玉米黃質的變化情況。在分析p515信號變化情況的基礎上,對組成質子動力勢PMF的跨膜電勢ΔΨ和跨膜質子勢ΔpH進行了量化分析。
1.6 數據分析
采用Excel 2019軟件進行數據匯總,Origin 2021軟件對數據進行單因素方差分析,結果以3次獨立生物重復的平均值和標準誤表示,顯著性檢驗采用 Fisher LSD 方法。采用 Graphpad Prism 8 和Origin 2021 軟件作圖。
2 結果與分析
2.1 亞低磷脅迫下外源鈣對花生生長發育及葉片光合氣體交換的調控作用
如圖1所示,亞低磷處理顯著降低了花生干物質重和總葉面積。與CK相比,–P處理的花生干物質重和總葉面積分別下降了21.1%和30.3%,這說明亞低磷脅迫下花生的生長受到限制。與–P處理相比,–P+Ca處理的總干物質重和總葉面積分別提高了26.7%和31.9%,達到了CK水平。–P處理顯著降低了花生葉片的相對葉綠素含量,而–P+Ca處理相比–P處理顯著提高了花生葉片的相對葉綠素含量。亞低磷脅迫下,外源鈣有效緩解了花生生長發育所受的限制,促進了花生的生長。
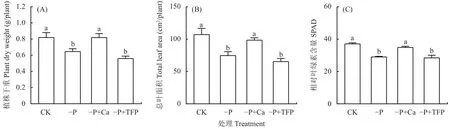
圖1 外源鈣對亞低磷脅迫下花生總葉面積、總干物質重和相對葉綠素含量 (SPAD) 的影響Fig.1 Effects of exogenous Ca2+ on total dry matter, total leaf area, and relative chlorophyll concentration (SPAD)of peanut under P deficiency
如圖2所示,與CK相比,–P處理下花生葉片的凈光合速率Pn顯著下降了19.1%,–P+Ca處理相比–P處理顯著提高了亞低磷脅迫下花生葉片的Pn(圖2A),提高了20.7%;–P處理下花生葉片的蒸騰速率Tr顯著下降,下降了19%,而–P+TFP處理則進一步誘導了亞低磷脅迫下Tr的顯著下降,外源鈣提高了亞低磷脅迫下花生葉片的Tr,但差異不顯著(圖2B);–P處理下花生葉片的胞間CO2濃度Ci上升,–P+TFP處理則進一步加大了這種上升趨勢,–P+Ca降低了亞低磷脅迫下花生葉片的Ci,但各處理間差異不顯著(圖2C);–P處理下花生葉片的氣孔導度Gs顯著下降,而–P+Ca處理顯著提高了亞低磷脅迫下花生葉片的Gs(圖2D),比–P處理提高了12.9%。
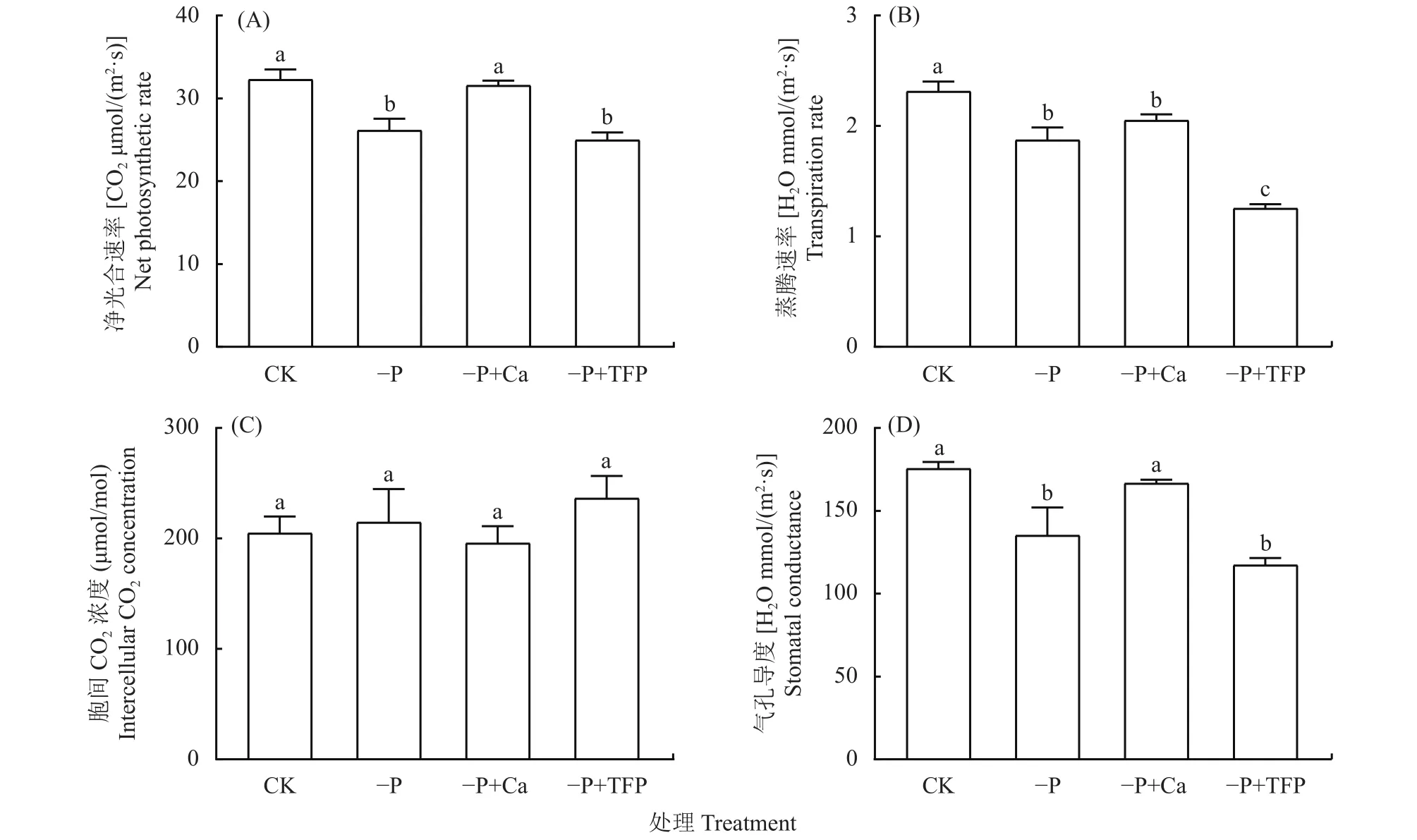
圖2 亞低磷脅迫下外源鈣對花生葉片氣體交換參數的影響Fig.2 Effects of exogenous Ca2+ on net photosynthetic rate, transpiration rate, intercellular CO2 concentration,and stomatal conductance of peanut under P deficiency
2.2 亞低磷脅迫下外源鈣對花生葉片PSⅡ活性的影響
如圖3所示,亞低磷脅迫顯著降低了花生葉片Y(Ⅱ),與CK相比,–P處理花生葉片Y(Ⅱ)下降了18.0%,而 Y(NO)和Y(NPQ)顯著增高,表明亞低磷脅迫下,PSⅡ光化學反應受阻,造成光能過剩。與–P處理相比,–P+Ca處理降低了Y(NO)和Y(NPQ),Y(Ⅱ)顯著提高,這說明外源鈣提高了PSⅡ活性,減緩了PSⅡ的光抑制。
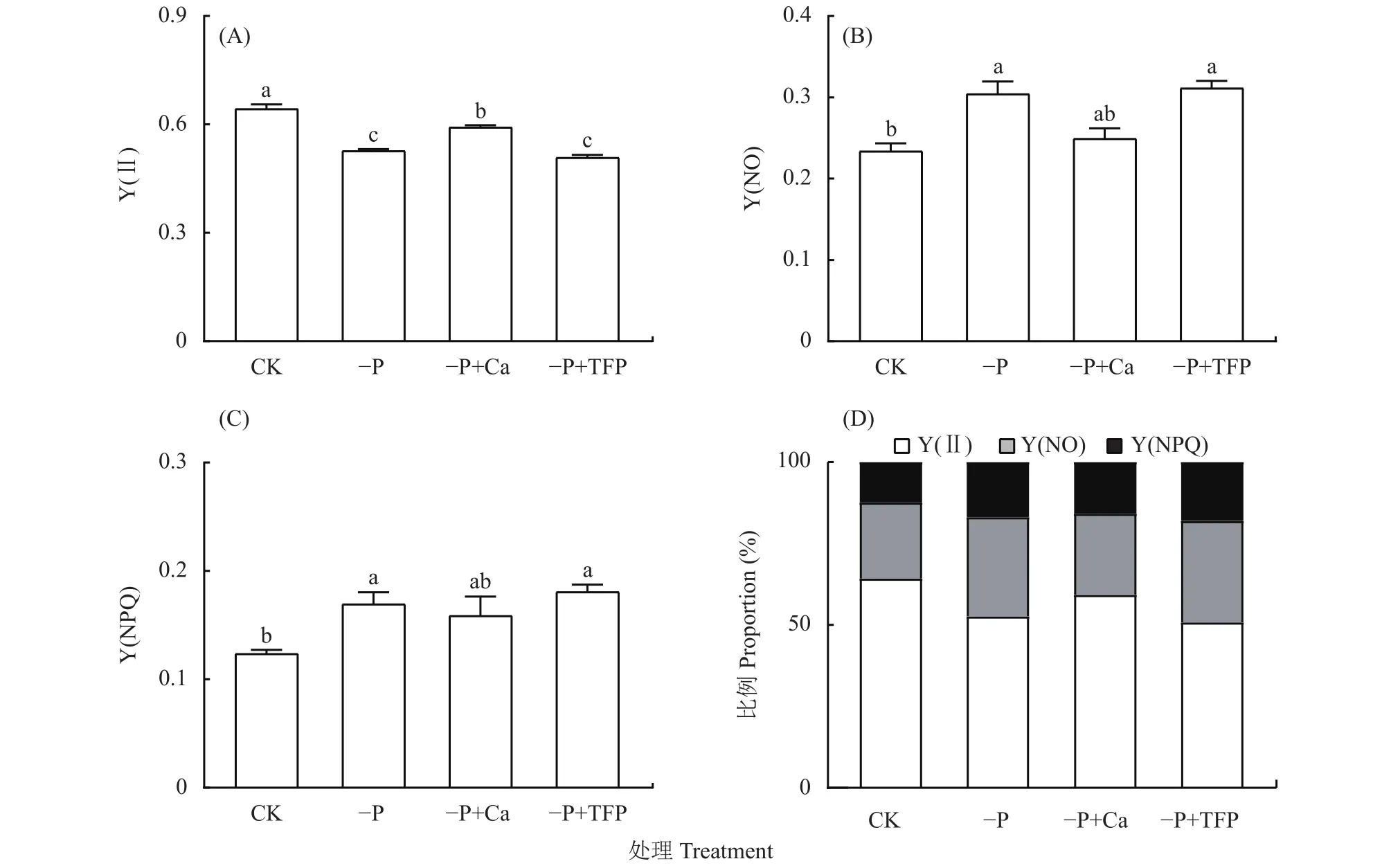
圖3 亞低磷脅迫下外源鈣對花生葉片光系統Ⅱ光化學活性的影響Fig.3 Effects of exogenous Ca2+ on the Y(Ⅱ), Y(NO), Y(NPQ), and their proportional allocation of peanut leaves under P deficiency
2.3 亞低磷脅迫下外源鈣對花生葉片PSⅠ活性的影響
如圖4所示,亞低磷脅迫顯著降低了花生葉片的Y(Ⅰ)。與CK相比,–P處理Y(ND)和Y(NA)上升,Y(Ⅰ)下降了5.4%,這說明亞低磷脅迫造成花生葉片PSⅠ光抑制。與–P處理相比,–P+Ca處理Y(ND)和Y(NA)下降,Y(Ⅰ)提高并達到了同CK無顯著差異的水平。說明外源鈣緩解了亞低磷脅迫對PSI供體側和受體側限制,緩解了PSⅠ光抑制。
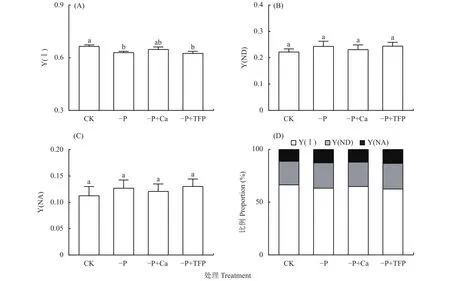
圖4 亞低磷脅迫下外源鈣對花生葉片PSI活性的影響Fig.4 Effects of exogenous Ca2+ on the Y(Ⅰ), Y(ND), Y(NA) and their proportional allocation of peanut leaves under P deficiency
2.4 亞低磷脅迫下外源鈣對花生葉片PQ庫和環式電子傳遞的影響
如圖5A和B所示,CK曲線下降斜率小,此時受脅迫程度小,未完全啟動環式電子傳遞。與CK相比,–P處理下降斜率升高,此時葉片已啟動環式電子傳遞。與–P處理相比,–P+Ca處理的曲線下降斜率進一步升高,說明外源鈣促進了亞低磷脅迫下花生葉片的環式電子傳遞。而–P+TFP處理的曲線下降斜率則小于CK和–P處理,這表明亞低磷脅迫下外源TFP處理進一步降低了環式電子傳遞速率,反向證明了鈣離子參與調節環式電子傳遞。如圖5C所示,亞低磷脅迫顯著降低了花生葉片的PQ庫大小,與CK相比,–P處理PQ 庫大小下降了22.1%,亞低磷脅迫誘發了花生葉片PQ庫的顯著減少。而外源鈣處理顯著提高了亞低磷脅迫下花生葉片的PQ庫大小,與–P處理相比,–P+Ca處理提升了21.3%,亞低磷脅迫下,外源鈣緩解了PQ庫的限制。
2.5 亞低磷脅迫下外源鈣對花生葉片類囊體膜的完整性和ATP合酶活性的影響
如圖6A所示,CK花生葉片經充分暗適應后,P515信號呈緩慢降低趨勢。與CK相比,–P處理下降斜率變大,這反映了亞低磷脅迫降低了花生葉片類囊體膜的完整性,造成類囊體膜損傷;–P+Ca處理下降斜率處于CK和–P處理之間,該結果說明亞低磷脅迫下外源鈣有效緩解了花生葉片類囊體膜的損傷程度。–P+TFP處理的下降斜率最高,類囊體膜損傷最嚴重。亞低磷脅迫下,外源TFP進一步降低了類囊體膜的完整性,反向說明了Ca2+對類囊體膜的保護作用。如圖6B所示,充分照光后,CK花生葉片的P515信號下降最快,其次是–P+Ca處理和–P處理,–P+TFP處理下降最慢。該結果說明,亞低磷脅迫降低了ATP合酶的活性,而外源鈣提高了ATP合酶的活性。亞低磷脅迫下,外源TFP進一步降低了ATP合酶活性。
2.6 亞低磷脅迫下外源鈣對花生葉片質子動力勢的影響
如圖7A所示,各處理中,CK花生葉片 ?Ψ最高,–P+TFP處理的 ?Ψ最低。亞低磷脅迫顯著降低了花生葉片的跨膜電勢?Ψ,與CK相比,–P處理的?Ψ下降了26.7%;而–P+TFP處理進一步降低了花生葉片 ?Ψ。外源鈣顯著提高了亞低磷脅迫下花生葉片的 ?Ψ,與–P處理相比,–P+Ca處理花生葉片的?Ψ提高了26.9%,–P+Ca處理達到CK水平。如圖7B所示,在各處理中,跨膜質子勢?pH 與 ?Ψ的變化趨勢相反。CK花生葉片?pH最低,–P+TFP處理的?pH最高。與CK相比,–P處理?pH升高了35.8%,這說明亞低磷脅迫造成花生葉片類囊體腔酸化。–P+TFP處理的?pH與–P處理處于同一水平。外源鈣顯著降低了亞低磷脅迫下花生葉片的?pH,與–P處理相比,Ca處理花生葉片的ΔpH降低了33.7%,–P+Ca處理與CK處于同一水平,這說明亞低磷脅迫下外源鈣有效緩解了花生葉片類囊體腔的酸化。
3 討論
花生具備較強根瘤固氮能力,但其對土壤有效磷含量具有較高要求[18]。本研究結果表明,亞低磷脅迫可造成花生干物質積累、總葉面積和相對葉綠素含量顯著降低,嚴重限制花生生長發育(圖1)。前人研究也發現,磷供應不足顯著降低高產花生品種的葉片生長和干物質量[10]。另外,亞低磷脅迫顯著降低花生功能葉片凈光合速率Pn、蒸騰速率Tr和氣孔導度Gs,全面抑制光合作用水平(圖2);許多研究也證實,磷限制條件下庫活性和生長抑制可導致植物光合反饋抑制[36–37]。亞低磷脅迫下,花生葉片葉綠體ATP合酶活性下降,質子在類囊體腔中積累,?pH上升,造成類囊體腔酸化。有研究表明,缺磷會限制Benson-Calvin循環,影響CO2同化[38]。Benson-Calvin循環的限制可能會積累磷酸化的中間產物并固定基質中的Pi[39],同時Benson-Calvin循環受限也會降低對NADPH和ATP的需求。CO2同化受限,會導致ATP/ADP值升高,降低ATP合酶活性[40]。有研究表明,缺磷會導致葉綠體基質中的Pi減少,而ATP合酶對葉綠體中Pi濃度變化非常敏感,基質Pi濃度的輕微下降也會對ATP合酶活性產生重大影響,從而顯著抑制質子的外流[41]。盡管Pi是ATP合酶合成ATP的底物,但Pi降低也會對其它代謝過程產生實質性的影響[42]。總而言之,缺磷會減少葉綠體基質中的Pi濃度,降低ATP合酶活性,并造成花生葉片類囊體腔酸化。
本研究發現,亞低磷脅迫不僅可以造成花生光合反饋抑制,同時還導致花生功能葉片光損傷(光抑制) (圖3、圖4)。在葉綠體中,質子動力勢PMF驅動ATP合酶合成ATP,而PMF由?pH和?Ψ組成,這兩個組分對ATP合成的貢獻相等,但只有?pH組分通過類囊體腔酸化來誘導光合電子傳遞速率的下調。PMF的調節要滿足兩個生理需求:1)滿足CO2固定所需的ATP;2)下調電子傳遞避免光損傷[43]。在本試驗中,亞低磷脅迫下花生葉片ΔpH上升和?Ψ下降(圖7)也說明,花生生長受限、ATP需求降低情況下,自身通過酸化類囊體腔下調光合電子傳遞速率避免光損傷,其中CEF的促進(圖5)和ATP合酶活性的下降(圖6)促進了ΔpH的升高,保護了光合膜、減少了ROS的產生。?pH具有兩方面功能:一是酸化的類囊體腔會下調細胞色素b6f復合體的活性,減少電子向PSⅠ傳遞,保護PSⅠ;二是誘導非光化學猝滅(NPQ)的qE組分來增加能量耗散[44]。有研究表明,環式電子傳遞在線性電子傳遞受限的條件下維持較大的ΔpH,從而誘導NPQ的qE組分耗散過多的光能。qE組分的激發會下調PSⅡ天線色素的光捕獲效率,從而防止PSⅡ的過度光損傷(光抑制),但因此也會降低PSⅡ的電子傳遞速率[45],這一結果與本試驗結果一致。在本試驗中,亞低磷脅迫下,?pH升高,類囊體腔酸化(圖8),亞低磷脅迫下Y(Ⅱ)顯著降低,而Y(NPQ)顯著增加,這表明包括qE在內的調節性能量耗散機制已啟動,消耗過剩光能。另一方面,升高的ΔpH會減緩PQH2在細胞色素b6f復合體處的氧化,限制電子向PSⅠ傳遞[46–49],而本試驗中,亞低磷脅迫下,Y(Ⅰ)顯著下降,光系統Ⅰ也發生光抑制。

圖5 亞低磷脅迫下外源鈣對花生葉片環式電子傳遞和PQ庫的影響Fig.5 Effects of exogenous Ca2+ on CEF and PQ pool of peanut under P deficiency
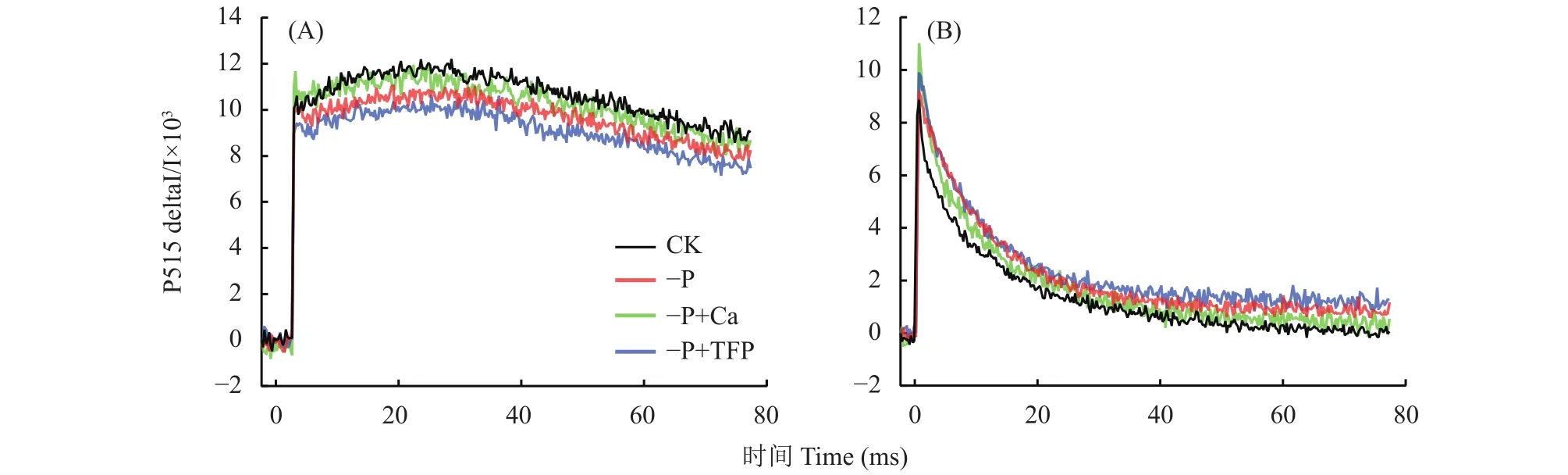
圖6 亞低磷脅迫下外源鈣對花生葉片類囊體膜的完整性和ATP合酶活性的影響Fig.6 Effects of exogenous Ca2+ on 515 rapid relaxation kinetics of peanut leaves under P deficiency
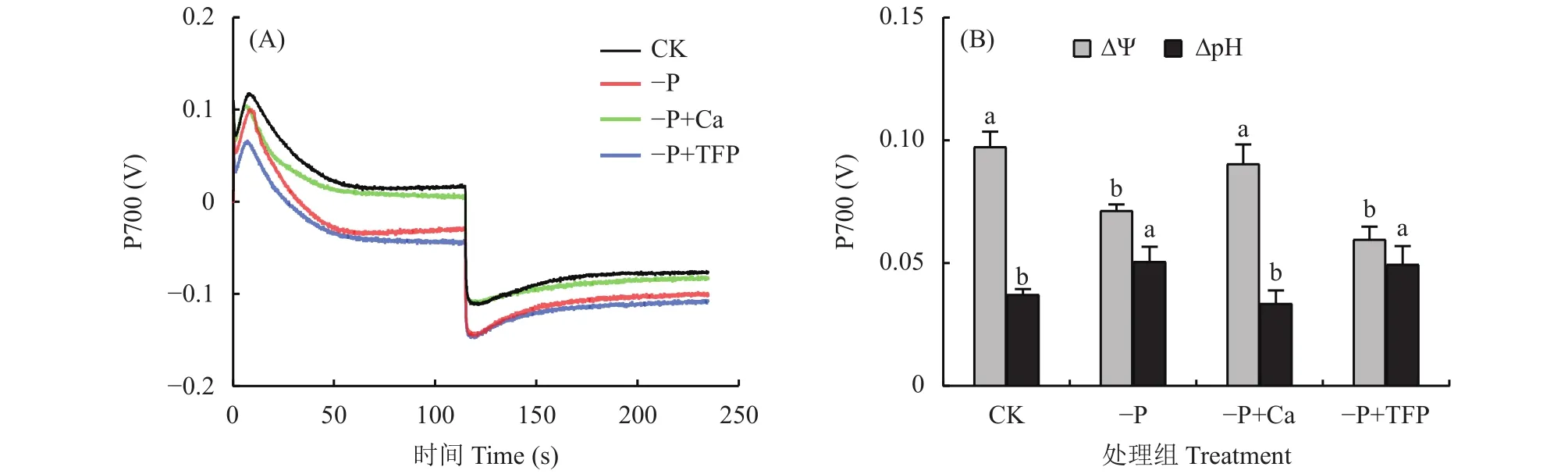
圖7 亞低磷脅迫下外源鈣對花生葉片質子動力勢的影響Fig.7 Effects of exogenous Ca2+ on proton motive force of peanut under P deficiency

圖8 亞低磷脅迫下外源鈣對花生葉片光合電子傳遞鏈的影響Fig.8 Effects of exogenous Ca2+ on photosynthetic electron transport chain of peanut under P deficiency
Ca2+作為第二信使[24],在植物外源磷感知和脅迫信號傳遞中具有重要調節作用[50]。有研究表明,Ca調節了植物葉片磷的分配[51–52]。有些物種通過優先將磷分配到光合細胞中,以便更有效地利用磷,提高光合磷利用效率(PPUE)[23]。本試驗結果表明,外源鈣提高了亞低磷脅迫下花生的干物質重和總葉面積(圖1),有效緩解了亞低磷脅迫對花生植株生長發育的限制。這與Ding等[53]的研究結果一致,提高鈣供應可以緩解羽扇豆的缺磷癥狀,并促進其生物量的積累。外源鈣提高了花生的凈光合速率(圖2),增加了花生植株對同化物的需求,緩解了花生光抑制;特別是亞低磷脅迫下,外源鈣可激發花生葉片環式電子傳遞,提高花生葉片光保護能力,緩解光損傷[54–56]。本研究結果表明,外源鈣提高了亞低磷脅迫下花生植株葉片 Y(Ⅰ)和 Y (Ⅱ),其中 Y(NO)和 Y(NPQ)的降低說明PSⅡ提高了對光能的轉化效率,Y(NA)和Y(ND)的下降說明了PSI供體側、受體側限制減少,外源鈣緩解了光系統的光抑制,并激活了環式電子傳遞(圖8)。另外,亞低磷脅迫下,外源鈣提高了ATP合酶的活性,降低了花生葉片的ΔpH (圖7)。外源鈣緩解了亞低磷脅迫誘導的類囊體腔酸化,這具有兩方面作用:一方面,恢復了細胞色素b6f復合體的活性,平衡了PQ庫的氧化還原狀態,減緩了PSⅠ的光抑制;另一方面,降低了非光化猝滅NPQ的qE組分耗散的能量,上調PSⅡ天線色素捕光效率,減緩了PSⅡ的光抑制,提高了Y(Ⅱ) (圖8)。
4 結論
亞低磷脅迫顯著抑制花生的生長發育,降低花生植株干物質積累、葉面積和相對葉綠素含量,降低ATP合酶活性和造成類囊體腔酸化,進而降低Y(Ⅰ)和Y(Ⅱ),造成花生光抑制。亞低磷脅迫下,外源鈣能有效解除花生生長發育所受的限制和光抑制,緩解亞低磷脅迫誘導的ATP合酶活性限制和類囊體腔的酸化,顯著提高了花生葉片的光合作用水平。經過亞低磷脅迫下TFP反向調控試驗證實,花生鈣調蛋白(Ca2+-modulin)作為外源鈣(Ca2+)的受體,可在鈣離子緩解亞低磷光合障礙的營養信號轉導中起到重要作用。
