基于代謝組學(xué)分析黑莓、黑樹莓果實(shí)代謝物的差異
2022-07-16 11:47:23吳雅瓊張春紅楊海燕閭連飛李維林吳文龍
江蘇農(nóng)業(yè)學(xué)報(bào) 2022年3期
關(guān)鍵詞:黃酮
吳雅瓊 張春紅 楊海燕 閭連飛 李維林 吳文龍


摘要: 為了探究黑莓、黑樹莓成熟果實(shí)中主要代謝物的差異,利用液相色譜-質(zhì)譜技術(shù)對其進(jìn)行比較分析。結(jié)果表明,在正離子、負(fù)離子模式下分別存在1 288 個(gè)、971個(gè)具有顯著表達(dá)差異的代謝物,通過KEGG數(shù)據(jù)平臺(tái)分析發(fā)現(xiàn),代謝通路是富集差異表達(dá)代謝物數(shù)量最多的(共104個(gè)),其次是次生代謝物合成通路(有65個(gè)差異表達(dá)代謝物)。此外,黃酮和黃酮醇生物合成通路也是主要差異代謝物顯著富集的代謝途徑之一,包含11個(gè)差異代謝物。而在花色苷合成通路上,與黑莓果實(shí)相比,黑樹莓果實(shí)中有6個(gè)代謝物表達(dá)量下調(diào),而矢車菊素-3- O -葡萄糖苷代謝物表達(dá)量上調(diào)。在異黃酮合成通路中,與黑樹莓果實(shí)相比,黑莓果實(shí)中的( S )-柚皮素相對表達(dá)量增加了2.7倍。研究結(jié)果為更好地了解黑樹莓、黑莓果實(shí)代謝物的異同和高效利用懸鉤子屬果實(shí)中黃酮、花色苷等次生代謝物提供理論依據(jù)。
關(guān)鍵詞: 黑莓; 黑樹莓; 黃酮; 差異代謝物; 合成通路
中圖分類號: S663.9?? 文獻(xiàn)標(biāo)識碼: A?? 文章編號: 1000-4440(2022)03-0790-08
Metabolomics analysis of differentially expressed metabolites in blackberry and black raspberry fruits
WU Ya-qiong 1 , ZHANG Chun-hong 1 , YANG Hai-yan 1 , LYU Lian-fei 1 , LI Wei-lin 2 , WU Wen-long 1
(1.Institute of Botany, Jiangsu Province and Chinese Academy of Sciences, Nanjing 210014, China; 2.Co-Innovation Center for the Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China)
Abstract: To explore the differences of main metabolites between mature fruits of black raspberry and blackberry, liquid chromatography-mass spectrometry (LC-MS) was used to compare and analyze. The results showed that there were 1 288 ?and 971 metabolites with significant differences in positive and negative ion modes, respectively. Through the analysis of Kyoto Encyclopedia of Genes and Genomes (KEGG) data platform, it was found that metabolic pathways were the most abundant (104 differentially expressed metabolites), followed by secondary biosynthesis (65 differentially expressed metabolites). In addition, flavone and flavonol biosynthesis was also one of the major metabolic pathways with significant enrichment, including 11 differentially expressed metabolites. In the anthocyanin biosynthesis pathway, compared with the mature fruit of blackberry, there were six differentially expressed metabolites down-regulated in the mature fruit of black raspberry. The expression level of cyanidin-3- o -glucoside metabolites was up-regulated in the black raspberry. In the isoflavonoid biosynthesis pathway, the relative expression of ( S )-naringenin in blackberry fruit was 3.7 times as much as that in black raspberry. The results can provide a theoretical basis for better understanding the similarities and differences of metabolites in black raspberry and blackberry fruits, and efficient utilization of secondary metabolites such as flavonoids and anthocyanins in ?Rubus ?fruits.
Key words: blackberry; black raspberry; flavonoid; differentially expressed metabolites; biosynthesis pathway
黑莓( Rubus ??fruticosus ?L.)和黑樹莓( Rubus occidentalis ?L.)都是薔薇科(Rosaceae)懸鉤子屬( Rubus )的多年生灌木,亦是重要的小漿果類果樹 [1-4] 。黑樹莓的栽培起源于歐洲,而黑莓原產(chǎn)于北美洲東部和歐洲。黑莓最早由江蘇省中國科學(xué)院植物研究所于1986年引入國內(nèi),經(jīng)過30多年的研究與推廣,目前已遍及全國各地,北起北京市,南到廣東省北部,東起東海之濱,西至云南省山地。懸鉤子屬植物果實(shí)中都含有豐富的黃酮、花色苷、糖類、有機(jī)酸、維生素和礦物質(zhì)等營養(yǎng)成分 [1,5] ,具有抗氧化、抗腫瘤等藥用價(jià)值 [4,6] 。
核磁共振(Nuclear magnetic resonance spectroscopy, NMR)、穩(wěn)定性同位素、氣相色譜-質(zhì)譜聯(lián)用(Gas chromatography-mass spectrometry, GC-MS)和液相色譜-質(zhì)譜聯(lián)用(Liquid chromatography-mass spectrometry,LC-MS)等技術(shù)是近年來常用于檢測代謝物的方法,其中LC-MS技術(shù)以其靈敏、高效、快速、檢測范圍廣泛等特點(diǎn)而在植物活性成分的分析檢測、植物品種和產(chǎn)地區(qū)分、果實(shí)品質(zhì)差異分析等方面得到廣泛應(yīng)用 [4,7-8] 。Wu等 [7] 采用GC-MS、LC-MS技術(shù)對東京野茉莉( Styrax tonkinensis )種仁發(fā)育過程中的代謝物變化規(guī)律進(jìn)行分析。劉龍桀等 [9] 基于GC-MS技術(shù)分析了 東北紅豆杉( Taxus cuspidata )野生種與栽培種的代謝物差異。高靜怡等 [10] 利用非靶向代謝組學(xué)對與砂梨水心病相關(guān)的不同代謝物進(jìn)行差異分析。Wu等 [4] 利用LC-MS技術(shù)對懸鉤子屬5個(gè)不同品種的成熟果實(shí)中與呈色相關(guān)的代謝物進(jìn)行分析,得出其成熟果實(shí)呈色與黃酮類組分密切相關(guān)。
黑莓、黑樹莓果實(shí)是黃酮等具有藥用價(jià)值的多酚類化合物的提取物來源以及提取花色苷的重要原材料,目前已有相關(guān)研究對不同黑莓品種或同一黑莓品種不同果實(shí)發(fā)育階段的總酚、總黃酮和6種常見花色苷類物質(zhì)進(jìn)行分析 [11] ,但尚未有針對性地比較黑莓、黑樹莓果實(shí)存在的初生代謝產(chǎn)物和次生代謝產(chǎn)物種類和組分含量差異的研究。為了有針對性地研究黑莓、黑樹莓代謝物的差異,本研究通過非靶向代謝組學(xué)技術(shù)分析黑莓、黑樹莓果實(shí)中與主要營養(yǎng)成分合成相關(guān)的代謝物異同,擬通過對參與生物體新陳代謝和維持正常功能的相對分子質(zhì)量小于1 000 的代謝物進(jìn)行定量分析,并對已鑒定的代謝物進(jìn)行分類,同時(shí)分析差異代謝物相關(guān)代謝通路信息,確定主要富集代謝途徑,從而闡明其代謝物的變化規(guī)律 [12-14] ,以期減少重要代謝物提取和育種相關(guān)技術(shù)研究的盲目性,為今后懸鉤子屬果實(shí)營養(yǎng)成分提取和藥用價(jià)值的開發(fā)與利用提供理論依據(jù)。
1 材料與方法
1.1 植物材料
于2020年6月采集黑莓品種Chester和黑樹莓品種Bristol的成熟果實(shí),每個(gè)品種設(shè)3個(gè)生物學(xué)重復(fù),將采集的果實(shí)暫時(shí)放在液氮罐中,帶回實(shí)驗(yàn)室后再放入-80 ℃ 超低溫冰箱中備用。本研究所用黑莓、黑樹莓品種均種植于江蘇省中國科學(xué)院植物研究所溧水白馬科研基地(119°09′E, 31°35′N)。
1.2 試驗(yàn)方法
提取代謝物的具體方法:各稱量200 mg黑莓Chester、黑樹莓Bristol果實(shí)樣品加入2 ml無酶離心管中,加入0.6 ml ?L -2-氯苯丙氨酸(4 mg/L ),渦旋振蕩30 s后在高通量組織研磨儀器中于50 Hz研磨60 s,在室溫下超聲提取15 min,于25 ℃、12 000 ?r/min 高速離心10 min。分別取300 μl上清液,經(jīng)0.22 μm膜過濾后,將濾液移入檢測瓶內(nèi)進(jìn)行LC-MS檢測。用于LC-MS上機(jī)檢測的儀器為Thermo Vanquish,具體的色譜、質(zhì)譜條件參考Wu等 [4] 的報(bào)道。
1.3 多元統(tǒng)計(jì)分析
為了初步顯示不同樣本組間的差異,首先對所有樣本采用無監(jiān)督降維方法——主成分分析(Principal component analysis,PCA)法進(jìn)行統(tǒng)計(jì)分析。然后基于有監(jiān)督的降維方法,采用偏最小二乘判別分析(Partial least squares discriminant analysis,PLS-DA)法和正交偏最小二乘判別分析(Orthogonal projection to latent structures-discriminant analysis,OPLS-DA)法來更好地區(qū)分2組樣品的代謝組學(xué)特征和類別,具體參照Yue等 [15] 的方法。
1.4 差異代謝物分析
采用(O)PLS模型的變量投影重要指標(biāo)( VIP )評分對黑莓、黑樹莓2組間最有區(qū)別/差異的代謝物進(jìn)行排序。當(dāng) t 檢驗(yàn)的 P < 0.06且 VIP ≥ 1時(shí),認(rèn)為2組樣品之間的代謝產(chǎn)物存在差異,即存在差異代謝物。
1.5 KEGG通路分析
京都基因與基因組百科全書(Kyoto Encyclopedia of Genes and Genomes,KEGG)是主要研究基因組、化學(xué)和系統(tǒng)功能信息的公共數(shù)據(jù)庫,不僅包括基因信息,還包括代謝物信息。代謝產(chǎn)物被映射到KEGG代謝途徑進(jìn)行途徑分析和富集分析。通過途徑富集分析發(fā)現(xiàn),與整個(gè)背景相比,差異代謝產(chǎn)物中的代謝途徑或信號轉(zhuǎn)導(dǎo)途徑明顯得到富集 [16] 。經(jīng)錯(cuò)誤發(fā)現(xiàn)率(False discovery rate, FDR)校正,計(jì)算的 P 值取 FDR ≤ 0.05作為閾值,滿足這一條件的途徑被定義為差異代謝物中顯著富集的途徑。
2 結(jié)果與分析
2.1 樣本質(zhì)量控制和定性、定量分析
首先對黑莓和黑樹莓果實(shí)的所有代謝物進(jìn)行峰識別、峰過濾和峰對齊,得到質(zhì)核比、保留時(shí)間和峰面積等信息的數(shù)據(jù)矩陣,再根據(jù)精確相對分子質(zhì)量(誤差≤ 1.5× 10 -5 )進(jìn)行代謝物的鑒定,后續(xù)根據(jù)MS/MS碎片模式比對至Human Metabolome Database (HMDB, http://www.hmdb.ca)、METLIN (http://metlin.scripps.edu)、Massbank (http://www.massbank.jp/)、LipidMaps (http://www.lipidmaps.org)、mzClound (https://www.mzcloud.org)數(shù)據(jù)庫。代謝物的鑒定結(jié)果顯示,在正離子模式下存在17 906個(gè)峰,其中15 788個(gè)峰對應(yīng)無法定性的物質(zhì),從二級譜圖(MS2)中鑒定到413個(gè)已知代謝物,從一級譜圖(MS1)中鑒定到1 705 個(gè)已知代謝物。在負(fù)離子模式下存在15 035 個(gè)峰,其中13 946 個(gè)峰對應(yīng)無法定性的物質(zhì),從二級譜圖、一級譜圖中分別鑒定出194個(gè)、895個(gè)已知代謝物。
2.2 代謝物的多元統(tǒng)計(jì)分析
研究結(jié)果表明,在正離子模式下,第一主成分(PC1)的方差貢獻(xiàn)率為65.4%,第二主成分(PC2)的方差貢獻(xiàn)率為11.7%,共解釋了77.1%的原始變量信息(圖1A)。在負(fù)離子模式下,前2個(gè)主成分共解釋了77.7%的原始變量(PC1的方差貢獻(xiàn)率為65.7%,PC2的方差貢獻(xiàn)率為12.0%)(圖1B)。由圖1A(正離子模式)、圖1B(負(fù)離子模式)可以看出,黑樹莓1和黑樹莓2間的代謝物差異較小,黑樹莓3與黑樹莓1、黑樹莓2間存在輕微的分散,而黑莓組的3個(gè)生物學(xué)重復(fù)間的代謝物差異較小。總體可見,不同組間的代謝物基本能夠得到區(qū)分,組內(nèi)代謝物的差異小于組間差異。
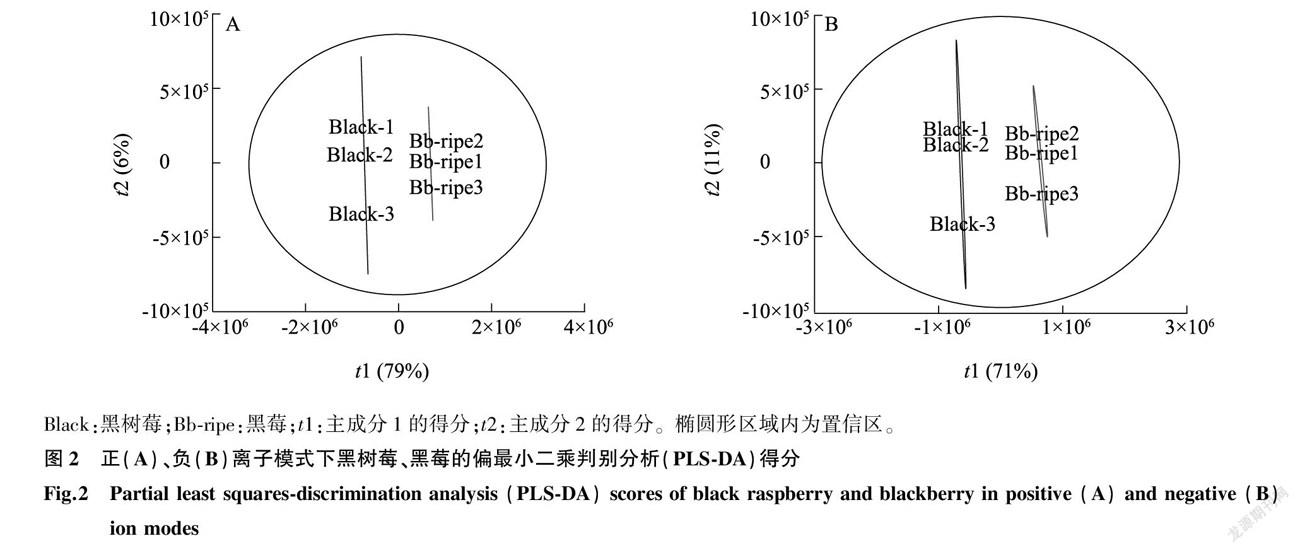
PLS-DA可以使組間區(qū)分效果最優(yōu)化,有利于尋找差異代謝物。本研究發(fā)現(xiàn),在正離子模式下,黑樹莓、黑莓果實(shí)代謝物符合 R? 2? X =0.853, R? 2? Y =1, Q? 2? Y = 0.998( R? 2? X 、 R? 2? Y 分別表示所建模型對 X 、 Y 矩陣的解釋率, Q? 2 表示模型的預(yù)測能力)(圖2A);在負(fù)離子模式下,符合 R? 2? X =0.827, R? 2? Y =0.999, Q? 2? Y =0.992(圖2B),這3個(gè)模型的值均大于0.8,符合我們對試驗(yàn)數(shù)據(jù)模型的預(yù)期,說明本研究建立的PLS-DA模型參數(shù)合理且穩(wěn)定,可用于后續(xù)代謝物的相關(guān)性分析。黑樹莓在圖2的置信區(qū)左側(cè),而黑莓在圖2的置信區(qū)右側(cè),說明該P(yáng)LS-DA模型能夠有效區(qū)分這2種懸鉤子屬小漿果品種。
OPLS-DA結(jié)合了正交信號矯正(OSC)、PLS-DA 2個(gè)方法,通過OPLS-DA建模分析同樣得出,黑樹莓在置信區(qū)左側(cè),而黑莓在置信區(qū)右側(cè)(圖3A、圖3D),與PLS-DA模型的分析結(jié)果類似。黑樹莓、黑莓的OPLS-DA模型有2個(gè)主成分,在正離子模式下, R 2 X =0.853, R 2 Y =1.000, Q 2 Y =0.997(圖3A);在負(fù)離子模式下, R 2 X = 0.827, R 2 Y =0.999, Q 2 Y =0.994(圖3D),這3個(gè)模型的值均大于0.6,說明建立的OPLS-DA模型能夠有效顯示黑樹莓、黑莓的代謝產(chǎn)物的差異。通過置換驗(yàn)證OPLS-DA模型,得出OPLS-DA模型的可靠性較高(圖3B、圖3E)。此外,載荷圖有助于找出對黑莓和黑樹莓果實(shí)組間代謝物模式變化貢獻(xiàn)最大的代謝物,在橫坐標(biāo)方向上遠(yuǎn)離原點(diǎn)的變量對區(qū)分這2組樣本的貢獻(xiàn)最大(圖3 C、圖3F)。
2.3 差異代謝物的篩選和聚類
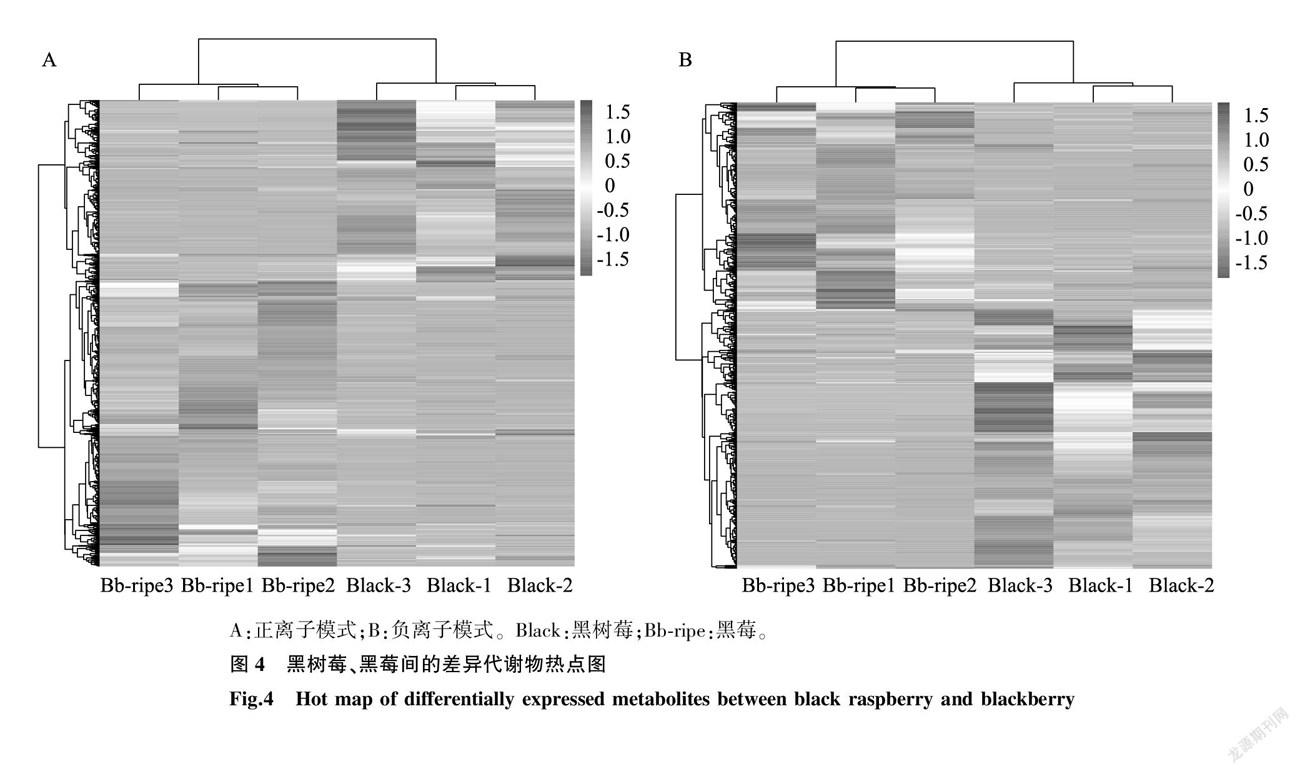
由于代謝組數(shù)據(jù)具有多維且某些變量間呈高度相關(guān)的特點(diǎn),代謝物的篩選過程往往比轉(zhuǎn)錄組中差異表達(dá)基因的分析更加復(fù)雜。基于此,我們結(jié)合單變量、多變量多元統(tǒng)計(jì)方法挖掘差異代謝物,對黑莓、黑樹莓成熟果實(shí)中的差異代謝物進(jìn)行數(shù)據(jù)的 Z -core標(biāo)準(zhǔn)化,然后進(jìn)行聚類分析。由圖4可以看出,在正離子模式下,與黑莓相比,黑樹莓有790個(gè)代謝物的表達(dá)量上調(diào),498個(gè)代謝物的表達(dá)量下調(diào),共有1 288個(gè)差異代謝物;在負(fù)離子模式下共有971個(gè)差異代謝物(與黑莓相比,黑樹莓有430個(gè)代謝物的表達(dá)量上調(diào),有541個(gè)代謝物的表達(dá)量下調(diào))。由此可見,不同懸鉤子屬品種間存在不同的初生、次生代謝產(chǎn)物積累差異。
2.4 差異代謝物通路分析
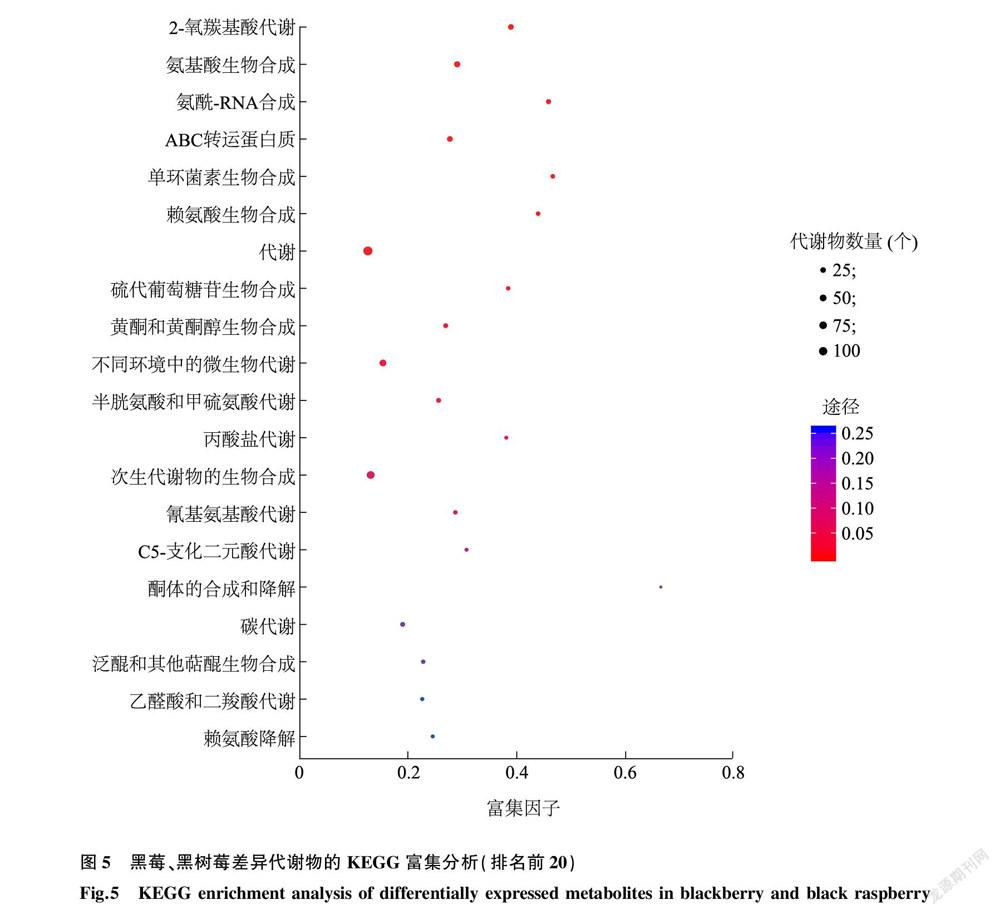
通過KEGG數(shù)據(jù)平臺(tái)對黑莓、黑樹莓果實(shí)的差異代謝物進(jìn)行代謝通路的差異富集分析,發(fā)現(xiàn)這些差異代謝物在84個(gè)KEGG通路中均有富集,筆者對所有差異代謝物中富集程度排名前20的代謝通路進(jìn)行分析,發(fā)現(xiàn)在排名前12的富集通路中,每條路徑均存在10個(gè)以上的差異代謝物。除了21個(gè)差異代謝物富集在ABC轉(zhuǎn)運(yùn)蛋白質(zhì)(ko02010, ABC transporters)通路[屬于環(huán)境信號處理類別(Environmental information processing class)]、13個(gè)差異代謝物富集在氨酰-RNA合成(ko00970, Aminoacyl-tRNA biosynthesis)通路[屬于遺傳信號處理類別(Genetic information processing class)]外,其余18個(gè)顯著富集差異代謝物的通路均屬于代謝物質(zhì)類別(Metabolism class)(圖5)。進(jìn)一步分析發(fā)現(xiàn),代謝通路(Metabolic pathways)是差異表達(dá)代謝物富集數(shù)量最多的,有104個(gè)差異表達(dá)代謝物,其次是次生代謝物的生物合成(Biosynthesis of secondary metabolites)通路,有65個(gè)差異表達(dá)代謝物。有趣的是,黃酮和黃酮醇生物合成(Flavone and flavonol biosynthesis)通路也是主要差異代謝物富集的代謝途徑之一,包含11個(gè)主要的差異表達(dá)代謝物,以黑莓果實(shí)為對照組,黑樹莓果實(shí)中有5個(gè)代謝物的表達(dá)量下調(diào),6個(gè)代謝物的表達(dá)量上調(diào)。此外,在花色苷合成(Anthocyanin biosynthesis)通路中,與成熟黑莓果實(shí)相比,香豆素(Pelargonidin)、香豆素3-(6- p -咖啡酰基)-葡萄糖苷[Pelargonidin 3-(6- p -caffeoyl)-glucoside]、紫草苷-5-葡萄糖苷-3-琥珀酸脂(Pelaronidin-5-glucoside-3-sambubioside)、矢車菊素-3-蕓香糖苷(Cyanidin-3-rutinoside)、矢車菊素-3-琥珀酸脂(Cyanidin-3-sambubioside)、花青苷(Cyanin)這6個(gè)差異代謝物的表達(dá)量下調(diào),而矢車菊素-3- O -葡萄糖苷(Chrysanthemin)代謝物的表達(dá)量上調(diào)。在異黃酮合成(Isoflavonoid biosynthesis)通路中,與黑樹莓果實(shí)相比,黑莓果實(shí)中( S )-柚皮素(Naringenine)相對表達(dá)量增加了2.7倍。
3 討論與結(jié)論
代謝組學(xué)是近年來發(fā)展較為迅速的一門組學(xué),代謝組被認(rèn)為是連接基因組和表型的橋梁,可為解析動(dòng)植物的代謝途徑和分子生化機(jī)制提供新的思路 [17-19] 。由于化學(xué)結(jié)構(gòu)與功能上的差異,代謝物主要分為初生代謝物和次生代謝物。研究發(fā)現(xiàn),大多數(shù)初生代謝物是植物產(chǎn)量、品質(zhì)變化的基礎(chǔ),亦是植物生長發(fā)育所必需的物質(zhì) [19] 。次生代謝物雖然不是植物生命活動(dòng)所必需的物質(zhì),但當(dāng)植物遭受外界逆境脅迫和病蟲危害時(shí),次生代謝物對維持植物體內(nèi)物質(zhì)的動(dòng)態(tài)平衡具有至關(guān)重要的作用 [19] 。不同物種間的次生代謝物的結(jié)構(gòu)、豐度存在著較大差異,不同組織間次生代謝物的含量、富集程度也存在顯著差異 [20] 。
懸鉤子類果樹因其果實(shí)中含有豐富的類黃酮、花色苷等多酚類物質(zhì)而聞名,具有較高的營養(yǎng)保健和藥用價(jià)值 [2, 4, 21-24] 。前人對黑莓、黑樹莓的研究多集中在不同營養(yǎng)物質(zhì)的積累模式或不同處理下代謝物的變化方面 [14] ,大多數(shù)研究僅對單一的物質(zhì)進(jìn)行研究,未有關(guān)于果實(shí)整體代謝水平的系統(tǒng)研究。基于此,本研究重點(diǎn)對黑樹莓、黑莓成熟果實(shí)進(jìn)行全面的代謝圖譜分析,結(jié)果發(fā)現(xiàn),在正離子模式下共有1 288個(gè)差異代謝物,在負(fù)離子模式下共有971個(gè)差異代謝物。由此可見,黑莓、黑樹莓成熟果實(shí)中的代謝物存在較大差異。
PCA和熱點(diǎn)聚類分析是用來評價(jià)生物學(xué)重復(fù)性是否可靠的方法 [25] 。本研究得出,黑樹莓、黑莓組內(nèi)的重復(fù)樣本最先聚為一類,說明樣本的重復(fù)性較好,數(shù)據(jù)分析結(jié)果可靠。有研究發(fā)現(xiàn),KEGG代謝通路的富集分析結(jié)果可為植物代謝過程中復(fù)雜的生物活動(dòng)研究提供有效的參考信息 [26] 。本研究通過對黑莓、黑樹莓的差異代謝物進(jìn)行KEGG代謝通路差異富集分析,得出84個(gè)KEGG通路均有富集,可能與果實(shí)成熟發(fā)育過程中代謝物不斷積累有關(guān),且前12個(gè)富集通路的每條路徑均存在10個(gè)以上的差異代謝物。有趣的是,差異代謝物富集數(shù)量最多(104個(gè))的是代謝通路,其次為次生代謝物(65個(gè))的生物合成通路。可能由于初生代謝物的不斷積累,為次生代謝物的合成和積累提供了物質(zhì)和能量,使得次生代謝物合成通路中的代謝物大量富集。
黑樹莓、黑莓果實(shí)均為提取天然花色苷類色素的原材料 [27-31] 。本研究發(fā)現(xiàn),香豆素、香豆素3-(6- p -咖啡酰基)-葡萄糖苷、紫草苷-5-葡萄糖苷-3-琥珀酸脂、矢車菊素-3-蕓香糖苷、矢車菊素-3-琥珀酸脂、花青苷等6個(gè)代謝物在成熟黑莓果實(shí)中的含量顯著高于成熟黑樹莓果實(shí),今后若要大量提取這6種代謝物,應(yīng)該選擇成熟的黑莓果實(shí)。矢車菊素-3- O -葡萄糖苷在黑樹莓成熟果實(shí)中含量較高,在提取該成分時(shí)可優(yōu)先考慮采用黑樹莓。綜上,本研究揭示的差異代謝物為今后高效利用懸鉤子屬果實(shí)中某一化學(xué)組分或營養(yǎng)成分如何優(yōu)先選擇含量較高的品種提供了現(xiàn)實(shí)依據(jù),同時(shí)為今后高效利用黃酮、花色苷等次生代謝物提供了理論基礎(chǔ)。
參考文獻(xiàn):
[1] THOLE V, BASSARD J E, RAMREZ-GONZLEZ R, et al. RNA-seq, de novo transcriptome assembly and flavonoid gene analysis in 13 wild and cultivated berry fruit species with high content of phenolics[J]. BMC Genomics, 2019, 20(1): 995.
[2] WU Y Q, ZHANG C H, YANG H Y, et al. Selection and validation of candidate reference genes for gene expression analysis by RT-qPCR in ?Rubus [J]. International Journal of Molecular Sciences, 2021, 22(19): 10533.
[3] FOSTER T M, BASSIL N V, DOSSETT M, et al. Genetic and genomic resources for ?Rubus ?breeding: a roadmap for the future[J]. Horticulture Research, 2019(1): 260-268.
[4] WU Y Q, ZHANG C H, HUANG Z J, et al. The color difference of rubus fruits is closely related to the composition of flavonoids including anthocyanins[J]. LWT, 2021, 149: 111825.
[5] ZURN J D, DRISKILL M, JUNG S, et al. A Rosaceae family-level approach to identify loci influencing soluble solids content in blackberry for DNA-informed breeding[J]. G3-Genes Genomes Genetics, 2020, 10(10): 3729-3740.
[6] KRZEPILKO A, PRAZAK R, SWIECILO A. Chemical composition, antioxidant and antimicrobial activity of raspberry, blackberry and raspberry-blackberry hybrid leaf buds[J]. Molecules, 2021, 26(2): 327.
[7] WU Q, ZHAO X, CHEN C, et al. Metabolite profiling and classification of developing ?Styrax tonkinensis ?kernels[J]. Metabolites, 2020, 10(1): 21.
[8] 張 舒,王長遠(yuǎn),馮玉超,等. 氣相色譜-質(zhì)譜聯(lián)用代謝組學(xué)技術(shù)分析不同產(chǎn)地稻米代謝物[J]. 食品科學(xué), 2021, 42(8): 206-213.
[9] 劉龍桀,吳可心,刁云飛,等. 基于GC-MS分析東北紅豆杉野生種與栽培種的代謝差異[J]. 植物研究, 2021,41(5):798-806.
[10] 高靜怡,馬 浩,劉東賀,等. 非靶向代謝組學(xué)對砂梨水心病不同代謝物質(zhì)的差異分析[J]. 分子植物育種, 2021, 19(24):8297-8304.
[11] 吳文龍,李維林,閭連飛,等. 黑莓引種栽培與利用[M]. 南京:江蘇科學(xué)技術(shù)出版社, 2010: 1-8.
[12] WECKWERTH W. Metabolomics in systems biology[J]. Annual Review of Plant Biology, 2003,54: 669-689.
[13] 路正禹, 王 堽, 李任任, 等. 基于組學(xué)技術(shù)探究甜菜耐鹽機(jī)理研究進(jìn)展[J]. 中國農(nóng)學(xué)通報(bào), 2021, 37(15): 92-98.
[14] KIM S, KIM J, KIM N, et al. Metabolomic elucidation of the effect of sucrose on the secondary metabolite profiles in ?Melissa officinalis ?by ultraperformance liquid chromatography-mass spectrometry[J]. ACS Omega, 2020, 5: 33186-33195.
[15] YUE S J, ZHAO D, PENG C X, et al. Effects of theabrownin on serum metabolites and gut microbiome in rats with a high-sugar diet[J]. Food & Function, 2019, 10(11): 7063-7080.
[16] KANEHISA M, ARAKI M, GOTO S, et al. KEGG for linking genomes to life and the environment[J]. Nucleic Acids Research, 2008, 36: 480-484.
[17] 張改君,苗 靜,郭麗穎,等. 多組學(xué)聯(lián)用在中藥作用機(jī)制研究中的應(yīng)用[J]. 中草藥, 2021, 52(10): 3112-3120.
[18] FAN M C, LIAN W J, LI T T, et al. Metabolomics approach reveals discriminatory metabolites associating with the blue pigments from ?Vaccinium bracteatum ?thunb. leaves at different growth stages[J]. Industrial Crops & Products, 2020, 147: 112252.
[19] FU A Z, WANG Q, MU J L, et al. Combined genomic, transcriptomic, and metabolomic analyses provide insights into chayote ( Sechium edule ) evolution and fruit development[J]. Horticulture Research, 2021, 8(1): 35.
[20] SCOSSA F, BROTMAN Y, ABREU L F, et al. Genomics-based strategies for the use of natural variation in the improvement of crop metabolism[J]. Plant Science, 2016, 242(S1): 47-64.
[21] WU Y Q, GUO J, ZHOU Q, et al. De novo transcriptome analysis revealed genes involved in flavonoid biosynthesis, transport and regulation in ?Ginkgo biloba [J]. Industrial Crops & Products, 2018, 124: 226-235.
[22] WU Y Q, ZHANG C H, WU W L, et al. Genome-wide identification and analysis of the MADS-box gene family and its potential role in fruit ripening in black raspberry ( Rubus occidentalis ?L.)[J]. Journal of Berry Research, 2021, 11(2): 301-315.
[23] 吳 林,張 強(qiáng),王 穎,等. 中國樹莓科學(xué)研究和產(chǎn)業(yè)發(fā)展的回顧與展望[J]. 吉林農(nóng)業(yè)大學(xué)學(xué)報(bào), 2021, 43(3): 265-274.
[24] 蔡成林,黃庭海,張青峰. 紅莓和黑莓葉黃酮類化合物抗氧化活性的比較研究[J]. 生物化工, 2020, 6(6): 33-35.
[25] LIU H, LI H F, GU J Z, et al. Identification of the candidate proteins related to oleic acid accumulation during peanut ( Arachis hypogaea ?L.) seed development through comparative proteome analysis[J]. International Journal of Molecular Sciences, 2018, 19(4): 1235.
[26] LI X R, PEI G S, LIU L S, et al. Metabolomic analysis and lipid accumulation in a glucose tolerant ?Crypthecodinium cohnii ?strain obtained by adaptive laboratory evolution[J]. Bioresource Technology, 2017, 235: 87-95.
[27] 唐曉姝,胡 博,陳雪梅,等. 6種小型漿果營養(yǎng)成分及DPPH自由基清除活性比較[J].江蘇農(nóng)業(yè)科學(xué),2021,49(20):182-187.
[28] 黃佳楠,管福琴,李林蔚,等. 黑果枸杞果實(shí)提取物中花色苷對酪氨酸酶活性的抑制作用初探[J].植物資源與環(huán)境學(xué)報(bào),2020,29(3):75-77.
[29] 戴意強(qiáng),吳 寒,單成俊,等. 藍(lán)莓酶解工藝優(yōu)化研究[J].江蘇農(nóng)業(yè)科學(xué),2020,48(17):219-224.
[30] 延海瑩,喬樂克,張京良,等. 樹莓營養(yǎng)及活性研究進(jìn)展[J]. 食品工業(yè), 2018, 39(7): 281-284.
[31] 趙慧芳,吳文龍,李維林,等. 黑莓Hull、Chester果實(shí)生長及品質(zhì)形成過程的分析[J]. 食品安全質(zhì)量檢測學(xué)報(bào), 2015, 6(8): 3199-3204.
(責(zé)任編輯:徐 艷)
收稿日期:2021-10-17
基金項(xiàng)目:國家自然科學(xué)基金項(xiàng)目(32101566);江蘇省自然科學(xué)基金青年基金項(xiàng)目(BK20210165);江蘇省中國科學(xué)院植物研究所博士人才科研啟動(dòng)基金項(xiàng)目(JIBTF202105)
作者簡介:吳雅瓊(1992-),女,江蘇南京人,博士,助理研究員,主要從事小漿果遺傳育種、果實(shí)營養(yǎng)品質(zhì)和基因組學(xué)方面的研究。(E-mail)ya_qiong@126.com
通訊作者:李維林,(E-mail)wlli@njfu.edu.cn;吳文龍,(E-mail)1964wwl@163.com
猜你喜歡
四川蠶業(yè)(2021年2期)2021-03-09 03:15:32
四川蠶業(yè)(2021年3期)2021-02-12 02:38:46
中成藥(2018年11期)2018-11-24 02:57:00
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
中成藥(2017年4期)2017-05-17 06:09:50
哈爾濱醫(yī)藥(2016年1期)2017-01-15 13:43:16
天然產(chǎn)物研究與開發(fā)(2016年11期)2016-06-15 20:29:17
湖南師范大學(xué)自然科學(xué)學(xué)報(bào)(2015年1期)2015-02-27 14:50:04
安徽醫(yī)藥(2014年12期)2014-03-20 13:15:15
