三江源區高寒退化草甸土壤細菌多樣性的對比研究
2022-06-25 01:24:52王英成姚世庭金鑫俞文政蘆光新王軍邦
生態環境學報 2022年4期
關鍵詞:差異
王英成,姚世庭,金鑫,俞文政,蘆光新*,王軍邦
1.青海大學,青海 西寧 810016;2.南京信息工程大學,江蘇 南京;3.中國科學院地理科學與資源研究所生態系統網絡觀測與模擬重點實驗室/生態系統大數據與模擬中心,北京 100101
土壤微生物種類繁多,主要包括細菌、真菌、古菌、原生動物以及病毒等(Torsvik et al.,2001)土壤微生物是土壤生態系統的重要組成成分,在土壤養分循壞、土壤肥力維持和土壤固氮等方面都發揮著重要作用,而且對于環境變化十分敏感(Martiny et al.,2011;劉洋熒等,2017;王朱珺等,2018)。在草地生態系統中,土壤微生物物種之間彼此相互依賴、相互制約,同時又和各類環境因子相互作用、往復調控(趙輕舟等,2018)。土壤微生物群落受到各種環境因子的影響,包括植被群落、土壤特征、氣候因子等(Gu et al.,2019),在草地生態系統中,草地類型(Wang et al.,2020)、草地退化(Che et al.,2019)等都會顯著改變草地土壤微生物的群落結構。
三江源區地處青藏高原腹部,其獨特的自然條件造就了該區域獨特的草地生態系統,高寒草甸作為該區域的主體植被類型,近年來,由于全球性的氣候變化、過度放牧等人為干預導致草地生態系統發生嚴重退化(Li et al.,2019;楊建平等,2004)。草地退化不僅引起地表植被群落發生顯著的變化,同時也會顯著影響土壤及地下微生物群落結構的改變。土壤是一個復雜的動態生物生態系統,它決定了陸地生態系統的植物生產力,并維持著生物地球化學循環(Wang et al.,2016;賀紀正,2013)。微生物對于土壤功能特別重要,尤其是在有機質分解和養分循環中(張衛信等,2020)。其中,土壤原核生物是土壤中數量最多、分布最為廣泛的微生物類群,在土壤生態過程中發揮著重要的作用。原核生物作為指示環境變化的敏感指標,對于預測土壤養分和環境質量的變化具有重要作用。之前有研究發現,退化土壤中的一些原核生物對于植物生長有害,因為草地退化改變了它們的代謝(Olff et al.,2000)。因此,高寒草地土壤養分狀況和植物多樣性的變化可能導致微生物群落結構的變化,進一步導致生態系統的退化。
近年來,分子生物學技術的發展和新一代高通量測序等技術的突破克服了傳統給微生物研究的局限性,極大的推動了微生物生態的發展(薛凱等,2019),越來越多的研究通過微生物這一微觀生物來揭示生態系統中的各種變化。Hou et al.(2019)通過高通量測序技術評估不同生態系統恢復方法對細菌群落多樣性的影響時,發現植被恢復后細菌群落多樣性顯著得到改善,且認為介導細菌群落多樣性的主要取決于恢復退化草地的植被類型。Zhou et al.(2019)利用高通量測序技術對未退化、輕度退化、中度退化、重度退化和極度退化5個退化階段土壤微生物群落的變化,結果發現在不同退化階段土壤微生物的群落組成并沒有很顯著的差異,但是土壤微生物的群落結構差異顯著,認為高寒草地退化對于土壤微生物的影響主要是通過介導土壤微生物群落的結構進而影響其功能。目前,很多關于退化草地的研究大多集中在微生物生物量、可培養微生物以及基因文庫的構建等(楊希智等,2015;李海云等,2019),這些傳統的方法對于我們解析地下微生物群落對草地退化的響應仍有所欠缺。因此,本研究采用巢式取樣法涵蓋足夠多的樣本量,并利用高通量測序技術手段全面的解析地下微生物群落多樣性,能夠很好地將我們所檢測的微生物用來評估退化草地微生物群落變化。以此來探究在高寒草地退化過程中土壤原核生物的群落組成、多樣性和結構變化,以期闡明并揭示在草地退化過程種土壤原核生物的變化規律,為我們進一步草地健康評價和退化草地恢復提供新的思考角度。
1 材料與方法
1.1 研究區概況
研究區位于青海省玉樹州稱多縣珍秦鄉,地理位置 33°21′35.04″N,97°20′37.38″E,平均海拔 4500 m,屬寒溫大陸性季風氣候,冷季長,暖季短,年均溫?6 ℃,無霜期較短,93—126 d左右。草地植被主要以高寒草甸為主,原生植被的主要優勢種有高山嵩草(Kobresiapygmaea),矮嵩草(Kobresia humilis),紫花針茅(Stipapurpurea)等,土壤為高寒草甸土。
1.2 研究方法
1.2.1 樣地設置與取樣方法
本研究依據裸露地塊地貌、草地植被群落特征以及草地的利用方式、強度選取兩塊典型的高寒草甸,即未退化草甸與退化草甸,采用巢氏取樣法進行土壤樣品的采集,即小樣方嵌套在大樣方內(圖1)。本研究中,以中心取樣區域(紅點,面積為 8 m2)作為參考點,12塊1 m2的采樣區域以垂直交叉方式分別設置在距采樣中心區域10、100、1000 m處(圖1)。其中,中心采樣區域包含17個土芯,每1 m2區域包含5個土芯。總共獲得154個樣本(2個采樣區域×77個采樣點)。于2019年8月12—13日,采用樣方法隨機用土鉆鉆取表層土壤(0—15 cm)二鉆,剔除可見植物殘體及土壤動物,封裝于帶有樣品編號的樣品袋中,一部分保存于實驗室4 ℃冰箱用于理化性質參數測定,一部分保存于?80 ℃冰箱用于DNA提取。
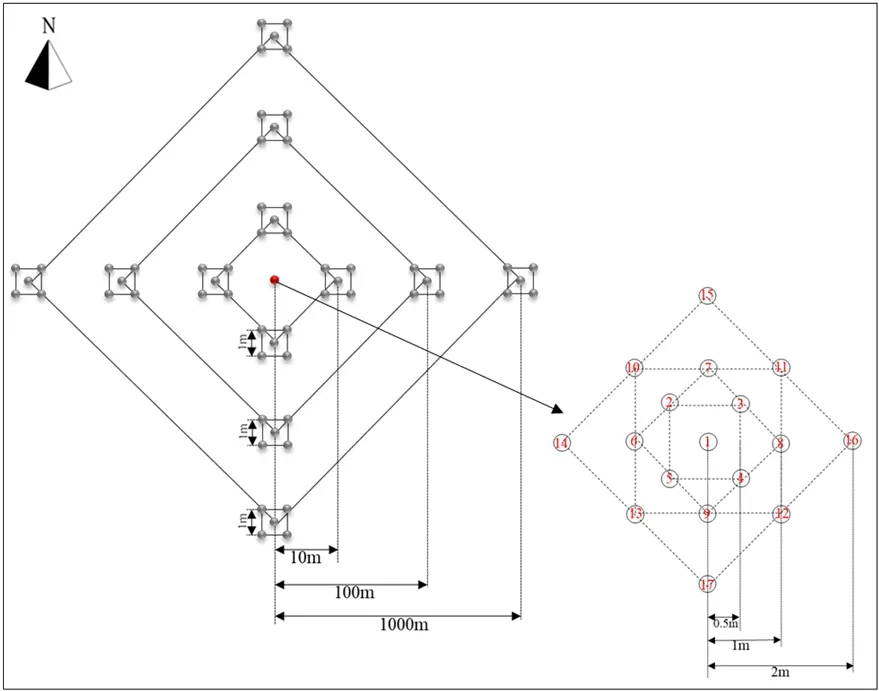
圖1 采樣設計圖Figure 1 Sampling plan
1.2.2 土壤理化參數指標測定
土壤部分主要是測定土壤全氮、土壤硝態氮、土壤銨態氮、土壤有機質、土壤pH值,土壤水分、土壤電導率。土壤pH用顯酸度計(PHS-3C)測得,水土比2.5?1。土壤全氮含量、有機質含量、銨態氮含量、硝態氮含量測定方法與姚世庭等(2021)方法一致。土壤水分、土壤電導率利用TDR 350儀器測定,重復3次,記錄數據。
1.2.3 土壤微生物測定
將采集的土壤樣品過 40目篩子預處理后,利用Mobio試劑盒(MOBIO Laboratories,Carlsbad,CA,美國)根據說明書從0.25 g土壤中提取總DNA(黨寧,2020),采用通用引物515F和806R擴增土壤中的16S rRNA基因的V3—V4區,基于Illumina Miseq測序平臺進行高通量測序。測序委托深圳美格基因公司進行測序。
1.3 數據分析
所有測序的原始數據均于中國科學院生態環境研究中心宏基因課題組構建的 Galaxy分析平臺(http://mem.rcees.ac.cn:8080/)進行相關的分析(Feng et al.,2017;Zhang et al.,2017)。將所獲得序列Reads根據標簽被分配到不同的樣品中并允許一個錯配的發生,然后去掉標簽和引物序列,使用FLASH步驟合并相同序列的正向和反向 reads(Magoc et al.,2011),使用Btrim對reads進行過濾(Kong,2011),保持平均質量分數大于20,16S最小長度為140 bp。任何帶有簡并堿基的序列都被刪除,只保留16S長度在245—260 bp之間的reads,使用 Greengenes參考數據集作為嵌合體檢測的參考,使用UPARSE(Edgar,2011)將序列聚類為可操作的分類單元(OTU),序列相似度閾值為97%。為了糾正樣本誤差,將所有的樣本隨機重抽到33159相同的reads數,重抽得到的OTU表進行后續的生物統計分析。
1.4 統計分析
采用ANOVA方差分析對土壤理化參數進行統計行檢驗,在P<0.05差異水平上檢測是否有顯著性差異。利用weighted UniFrac矩陣用于PCoA方法對兩個采樣地的土壤微生物群落的進化多樣性的差異進行研究分析,利用 Bray-curtis矩陣用于NMDS非度量多維尺度分析來可視化群落組成的差異,利用基于Bray-curtis和Jaccard距離矩陣用于Dissimilarity test檢驗不同群落之間組成的差異。使用Mantel和Partial mantel test以及典范對應分析CCA來確定植被因子和土壤理化參數對于微生物群落差異的相對貢獻。
2 結果與分析
2.1 高寒退化草甸植被群落結構差異分析
草地退化是一個復雜而長期的過程,草地退化最明顯的標志是植被群落結構的變化,包括植被生產力、群落組成結構等。本研究對高寒退化草甸與未退化草甸植被群落結構的分析(表1),結果發現未退化草甸的植被蓋度為87%,退化草甸的植被蓋度為72%。草地退化顯著降低植被物種數和地上生物量,退化草甸相比未退化草甸物種數減少了14.87%,地上生物量減少了19.47%。
結果顯示(表1),在試驗樣地共有草本植物15科 23屬 27種,其中種類最多的是菊科(Compositae),其次依次為莎草科(Cyperaceae)、薔薇科(Rosaceae)、龍膽科(Gentianaceae)、石竹科(Caryophyllaceae)、唇形科(Lamiaceae)、玄參科(Scrophulariaceae)、豆科(Leguminosae)、禾本科(Gramineae)等。未退化草甸植被以高山嵩草(Kobresiapygmaea)為主要優勢種,優勢度為47.58%,次優勢種為紫花針茅(Stipapurpurea)、重冠紫菀(Asterdiplostephioides)、矮生忍冬(Laniceraminuta),優勢度分別為21.35%、14.17%、10.40%。退化草甸的主要優勢種也是高山嵩草,優勢度為41.97%。次優勢種為紫花針茅、矮生忍冬、重冠紫菀、細葉亞菊(Ajaniatenuifolia)、高山唐松草(Thalictrumalpinum)、雪靈芝(ArenariakansuensisMaxim.)、苔草(Carexspp.)、麻花艽(Gentiana straminea),優勢度分別為19.47%、16.34%、16.19%、16.69%、12.16%、11.72%、11.35%、10.23%。
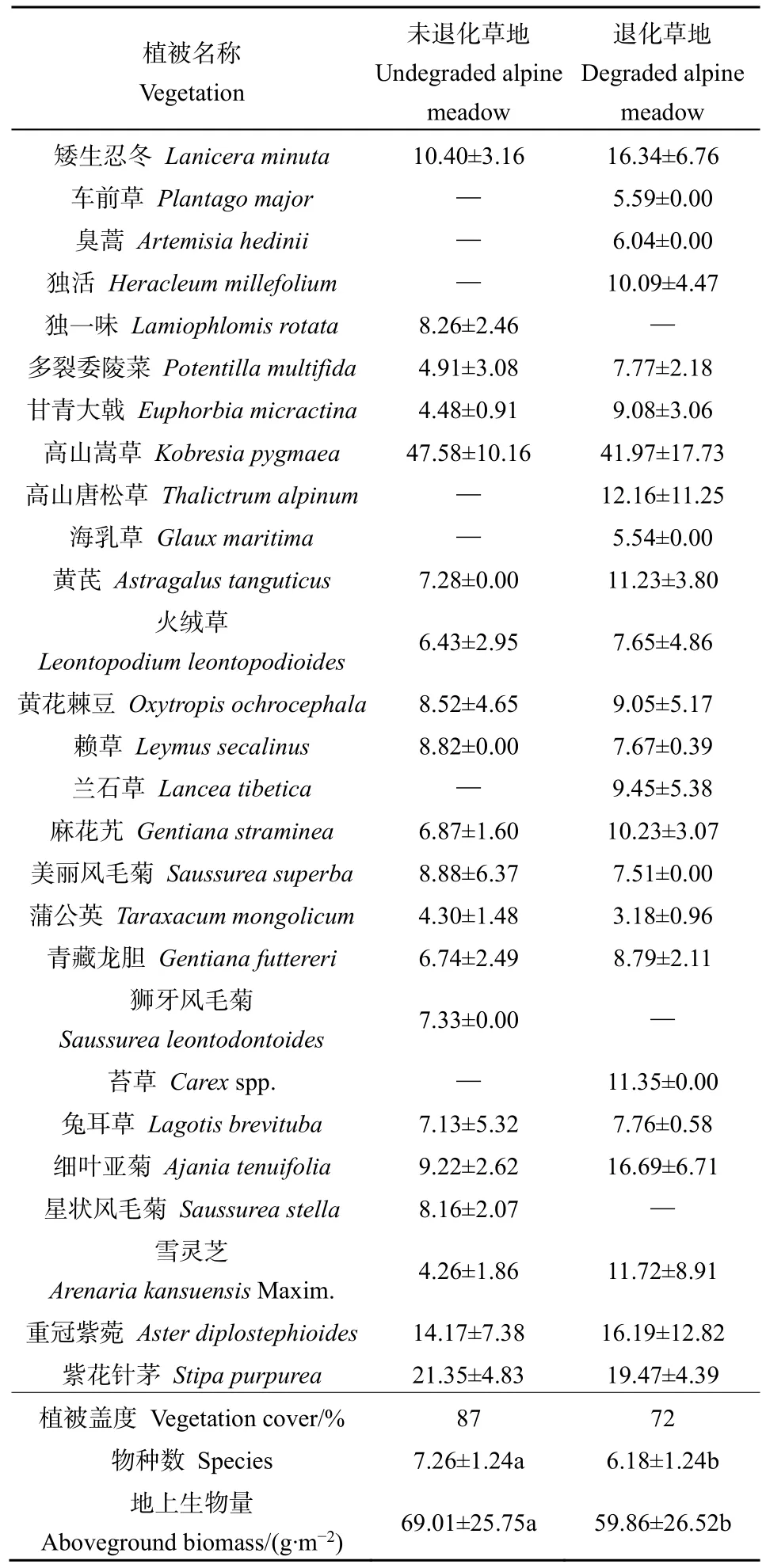
表1 高寒退化草甸植被群落組成(重要值)及其植被特征比較分析Table 1 Comparative analysis of vegetation community composition (significance value) and vegetation characteristics in alpine degraded meadow
2.2 高寒退化草甸土壤理化參數的差異分析
未退化高寒草甸與退化高寒草甸的土壤理化存在顯著的差異(表2)。從表2看到退化草甸的土壤全氮(TN)、有機質(OM)、土壤電導率(EC)顯著低于未退化草甸,分別低 16.00%、35.38%、46.50%。土壤pH值則是顯著高于未退化草甸,而土壤銨態氮(NH4+-N)、硝態氮(NO3?-N)、土壤水分(SM)并沒有顯著的差異。這些結果說明草地退化會對土壤理化性質產生顯著的影響,且主要體現在土壤pH值和土壤有機質方面。

表2 高寒退化草甸土壤理化差異比較Table 2 The measured vegetation factors and edaphic parameters of undegraded and degraded alpine meadow
2.3 高寒退化草甸土壤細菌多樣性分析
在三江源珍秦鄉的未退化樣地和退化樣地的154個樣本中,檢測到土壤細菌2168457條序列,在97%的相似性水平下,154個樣本得到71798種原核生物OTU(圖2a)。通過韋恩圖分析(圖2b),發現土壤原核生物在未退化樣地和退化樣地共有3776個OTU,占總OUT的38.8%,退化樣地獨有的OTU有3658個,占總OUT的37.6%,未退化樣地獨有的OTU有2287個,占總OTU的23.5%。
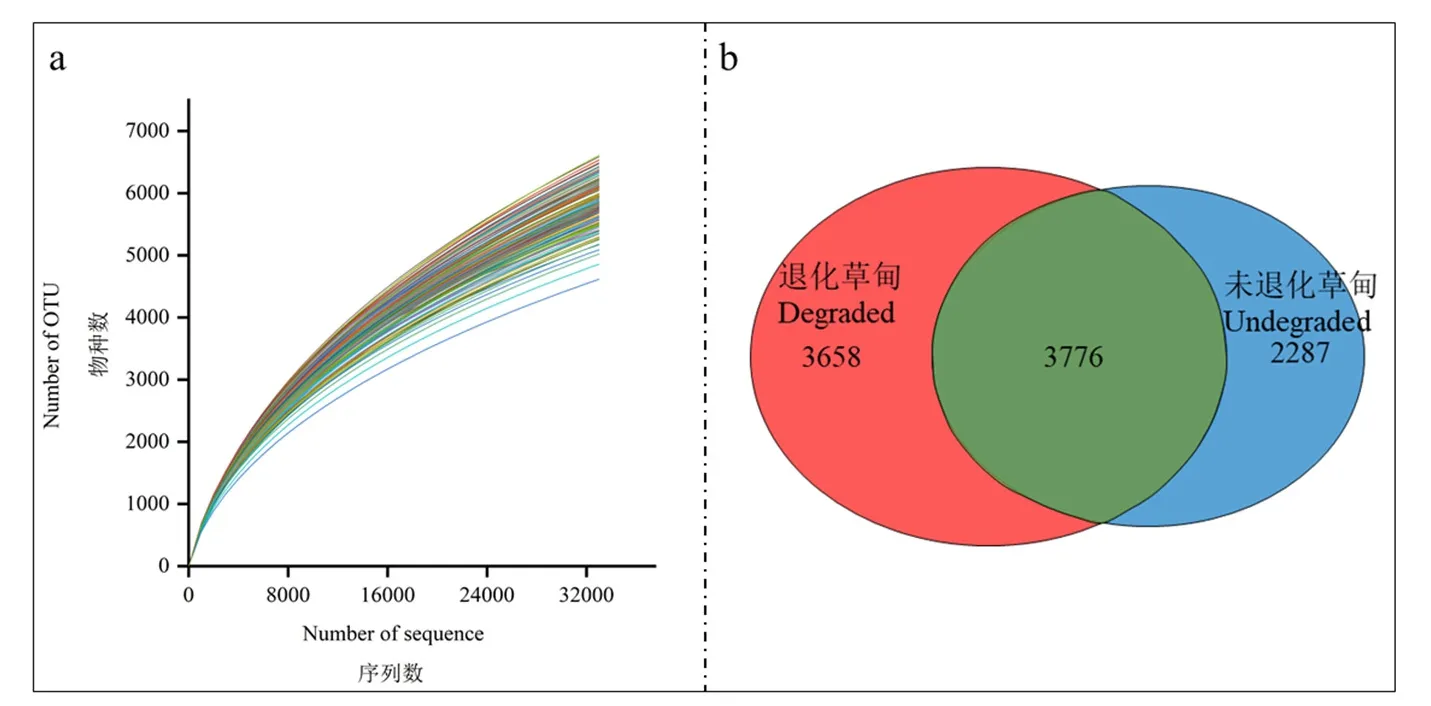
圖2 高寒退化草甸土壤樣品微生物OTU稀釋性曲線和韋恩圖Figure 2 Soil samples OTU dilution curve and Venn of alpine meadow degraded
對土壤細菌的α多樣性進行分析,結果發現三江源區高寒退化草甸土壤細菌在兩塊樣地之間存在顯著差異(圖3)。土壤細菌的物種豐富度指數(Observed Richness)、多樣性指數(Shannon Index)、chao1指數都是未退化草甸顯著高于退化草甸。

圖3 高寒退化草甸土壤細菌α多樣性分析Figure 3 α-diversity of soil bacterial of alpine meadow degraded
為了進一步探究土壤理化與土壤細菌多樣性之間的相互關系,對細菌多樣性與土壤理化性質進行了Pearson相關性分析(表3)。Pearson相關性分析表明土壤細菌α多樣性與土壤理化之間的相關性相對較弱,在未退化草甸只有Simpson指數與銨態氮含量存在顯著的負相關關系,而在退化草甸也僅只是Observed Richness與土壤電導率之間存在顯著的負相關關系,這一結果說明草地退化導致的土壤理化的改變對于土壤細菌群落的多樣性無顯著影響。
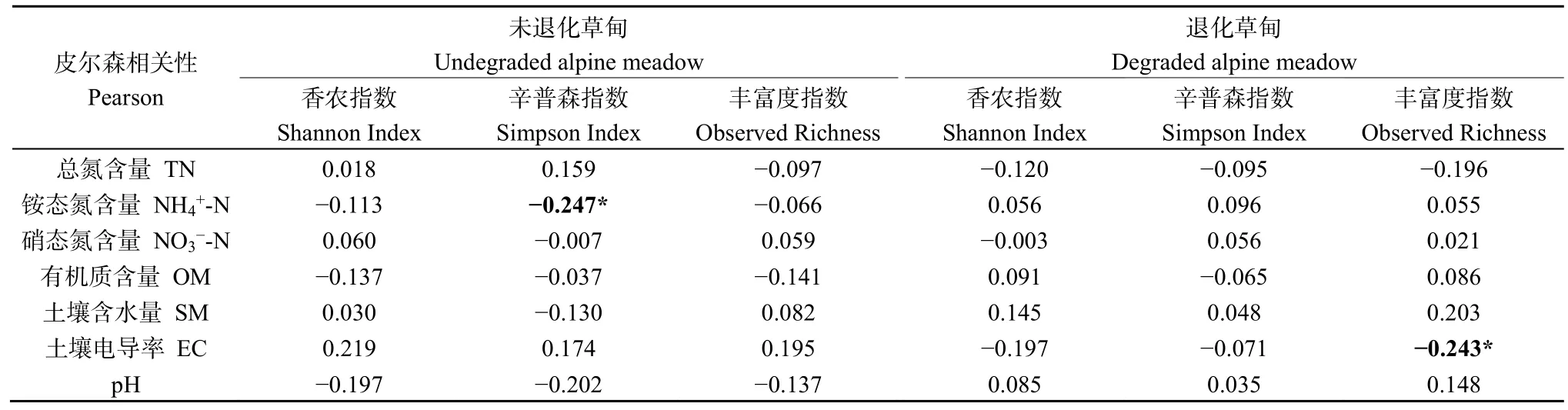
表3 高寒退化草甸土壤細菌α多樣性與土壤理化相關性分析Table 3 The α-diversity of bacterial community correction with soil chemical in degradation meadow
2.4 高寒退化草甸土壤細菌群落組成
為了研究三江源區退化高寒草甸土壤細菌群落組成成分,使用 Greengene基因庫對土壤細菌OTU進行注釋,分別在門水平和屬水平對土壤細菌組成成分進行比較(圖4)。在門水平上(圖4a),退化高寒草甸土壤細菌群落中變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、酸桿菌門(Acidobacteria)、擬桿菌門(Bacteroidetes)、疣微菌門(Verrucomicrobia)、奇古菌門(Thaumarchaeota)為主要優勢菌門,其相對豐度超過 1%。在不同樣地土壤中相對豐度最高的細菌菌門存在差異,如Actinobacteria和Bacteroidetes在退化草甸樣地中的相對豐度高于未退化草甸樣地,平均相對豐度分別為 25.02%,7.79%;而Verrucomicrobia在未退化草甸樣地的相對豐度高于退化草甸樣地,平均相對豐度為7.54%。土壤細菌屬的研究表明(圖 4b),在不同草甸樣地土壤中可鑒定細菌屬的總相對豐度比較均勻,平均相對豐度在39.72%。Spartobacteriageneraincertaesedis屬在退化草甸和未退化草甸中相對豐度最高,分別為4.19%、6.42%。Nitrososphaera屬在退化草甸中的相對豐度高于未退化草甸,為4.35%;Gaiella屬在未退化草甸中的相對豐度高于退化草甸,為3.74%;其中,其他優勢屬Pseudonocardia、WPS-1genera incertaesedis、Gemmatimonas、Terrimonas在退化草甸和未退化草甸中相對豐度比較均勻,在1.11%—1.80%之間;Sphingomonas、Nocardioides、Arthrobacter屬在退化草甸中相對豐度高于未退化草甸,分別為1.29%、1.04%、1.17%。
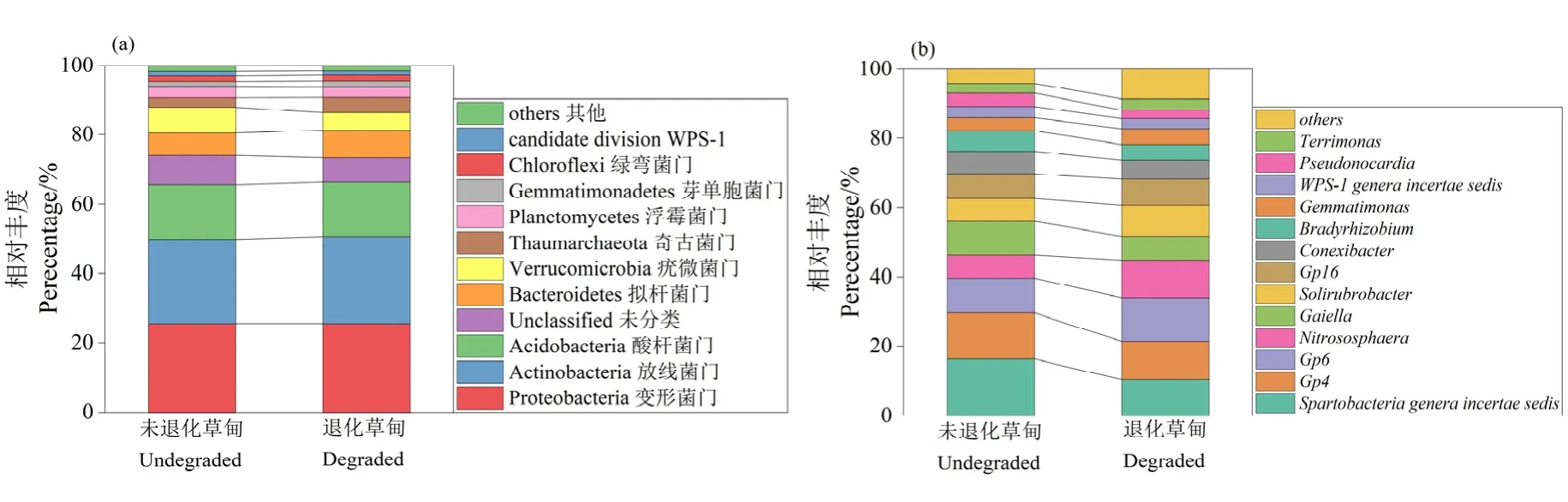
圖4 高寒退化草甸土壤細菌群落組成Figure 4 Composition of soil bacterial community in alpine meadow degraded
2.5 高寒退化草甸土壤細菌群落結構
采用基于weighted UniFrac的PCoA方法對兩個采樣地的土壤細菌群落的進化多樣性的差異進行研究分析;基于Bray-curtis距離矩陣的NMDS分析兩個采樣地的土壤細菌群落結構的差異,同時結合Bray Curtis距離和Jaccard距離的樣本組間差異顯著性檢驗(Dissimilarity)進行進一步驗證(表4)。對兩塊樣地土壤細菌PCoA的分析發現(圖5a),主成分軸PC1可以解釋24.26%的差異,PC2軸可以解釋15.55%的群落差異,通過圖5a看到,土壤細菌群落在兩塊樣地的所有樣本中能夠明顯區分,表明兩塊樣地的土壤細菌群落存在明顯的差異。
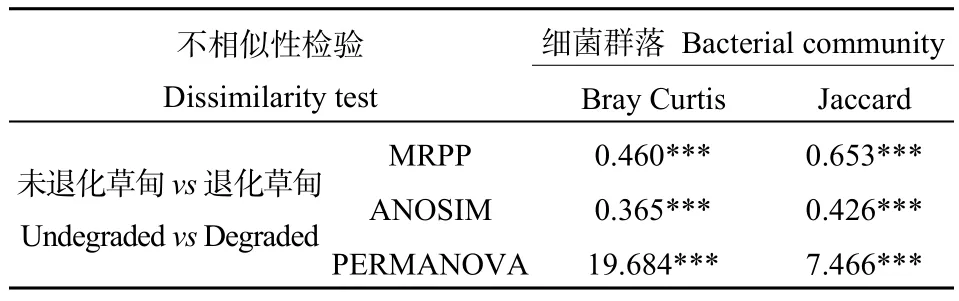
表4 高寒退化草甸土壤細菌群落的不相似性檢驗Table 4 Dissimilarity test of soil bacterial community in alpine meadow degraded
為了進一步研究高寒退化草甸土壤細菌群落 β多樣性,基于Bary Curtis距離,利用NMDS方法對未退化高寒草甸和退化高寒草甸的土壤細菌群落β多樣性進行分析。結果發現不同樣地之間的土壤細菌存在顯著的差異。基于Bray Curtis距離的土壤細菌群落NMDS圖(圖5b)可知,未退化高寒草甸與退化高寒草甸都可以明顯區分,且未退化高寒草甸細菌群落內部更加聚集,而退化高寒草甸細菌群落內部比較松散,這些結果表明,高寒草甸退化會對土壤細菌群落結構產生顯著的影響。Dissimilarity分析(表4)進一步表明土壤細菌群落在未退化樣地和退化樣地之間存在顯著的差異。這一結果說明高寒草甸退化會改變土壤細菌群落的 β多樣性。
2.6 土壤細菌群落與環境因子間的相互作用
土壤是土壤微生物的基質,土壤的理化性質和營養物質對于土壤中微生物的群落變化具有重要的作用,為了研究高寒退化草甸土壤細菌群落與土壤理化性質之間的相關關系,采用典范對應分析CCA方法對兩塊樣地的土壤細菌群落與土壤理化性質進行相關關系分析(圖 5c)。結果發現對于未退化高寒草甸樣地和退化高寒草甸樣地,土壤細菌群落與土壤總氮含量、硝態氮含量(有機質含量、pH值、電導率存在相關關系。Partial Mantel test檢驗結果(表5)進一步表明,對細菌群落影響最強的環境因子主要是土壤pH值、土壤有機質含量、土壤總氮含量。
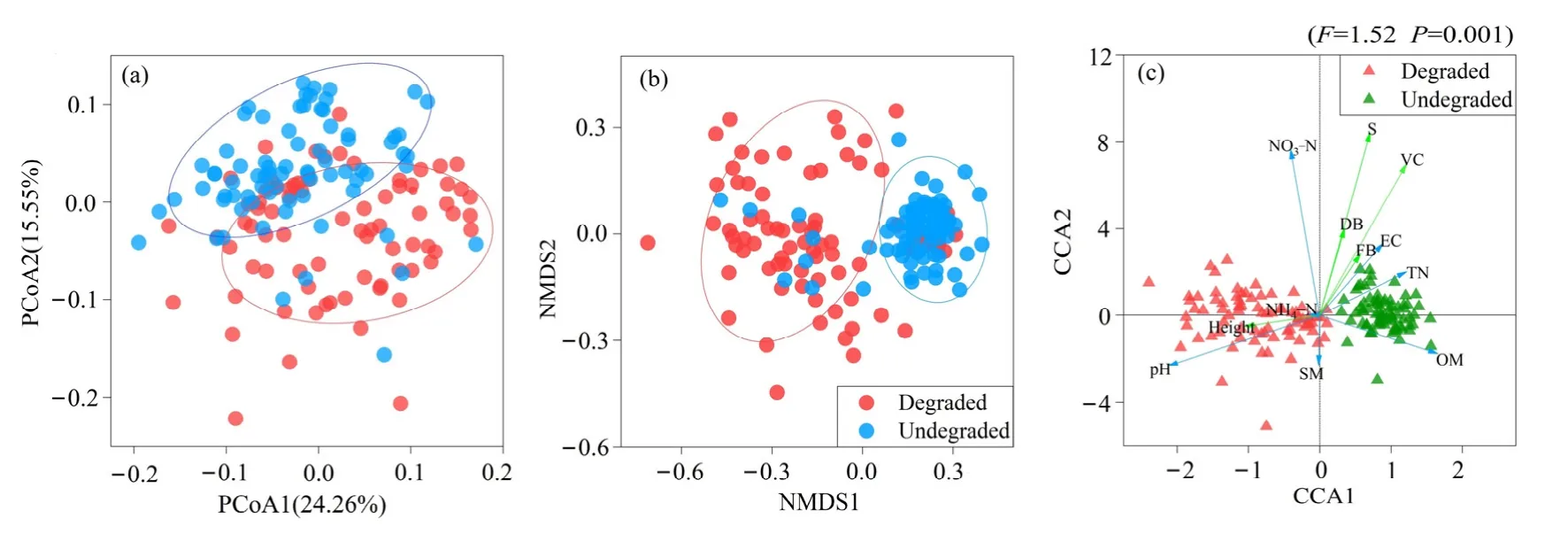
圖5 高寒退化草甸土壤細菌群落結構和環境因子相關性Figure 5 Correlation between soil bacterial community structure and environmental factors in alpine degraded meadow
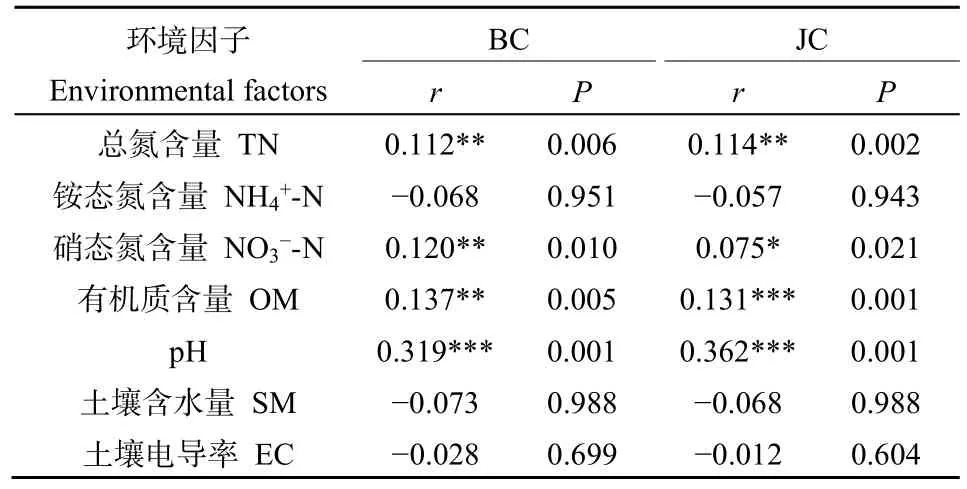
表5 土壤細菌群落差異與土壤理化的Partial Mantel檢驗Table 5 The correlation between the dissimilarity of soil bacterial communities and soil chemical using Partial Mantel test
3 討論
3.1 高寒草甸退化對植被和土壤影響
草地土壤微生物在維持草地生態系統穩定中發揮著重要的作用,草地退化最直觀的表現是影響地上植被群落,植被蓋度和物種豐度的下降等,而土壤退化是草地退化的本質核心,土壤結構和營養組成的變化對于地下生物和地上的生物都會產生影響。本研究結果表明,草地退化顯著降低了植被蓋度、物種數和生物量,這與之前的研究一致(劉玉等,2013;陳樂樂等,2016),認為草地退化必然的后果是植被群落的改變,這可能與草地的放牧壓力過大有關,由于家畜對優良牧草的采食,致使草地植被群落改變(李雪萍等,2020),同時鼠蟲害也會影響植被群落的改變。同時,草地退化也會對土壤理化性質產生顯著的影響,使得土壤更加貧瘠和向堿性化過渡(王英成等,2021)。
3.2 高寒草甸退化對土壤細菌群落多樣性和物種組成影響
在草地退化過程中,全球環境的變化、放牧和土地利用方式的改變都會對土壤造成一定的影響進而影響到地下微生物的群落,在不同程度的退化草地,地下微生物的分布和群落也存在顯著的不同(姚拓等,2006)。本研究結果發現,高寒草甸退化顯著改變了土壤細菌群落的α多樣性(圖3)和β多樣性(圖5)以及群落組成(圖4),這一結果與其他研究者對于青藏高原高寒退化草甸土壤微生物群落的研究相一致(Li et al.,2016;Zhou et al.,2019)。草地退化后,土壤細菌群落物種豐富度和多樣性顯著下降(圖2),這可能是因為草地生態系統的逆向演替在一定程度上抑制了土壤細菌群落的生長和繁殖,進而導致土壤細菌群落多樣性的減少。王啟蘭等(2007)研究認為高寒草地因其在低溫條件下土壤凍結期長、通透性差等會對地下微生物群落活動產生較強的抑制作用,進而不利于土壤微生物的生長繁殖。另外,本研究發現隨著高寒草甸的退化,土壤細菌在門水平和屬水平上的群落組成會發生顯著改變,其中,相對豐度較高的細菌群落菌門主要是變形菌門、放線菌門、酸桿菌門、擬桿菌門、疣微菌門、奇古菌門,這與多數對于高寒草地土壤微生物群落組成所得的研究結果基本一致(李海云等,2019)。有研究表明微生物的群落組成與微生物介導的生態功能有關(Bier et al.,2015;Che et al.,2015),根據本研究結果,推測高寒草甸的退化可能改變了微生物介導的生態功能,這一改變主要是通過改變微生物的群落組成實現而不是改變微生物的物種豐度。
3.3 高寒草甸退化土壤細菌群落與環境因子間的相互關系
土壤微生物群落的變化主要是因為土壤環境變量和其他活動的影響所造成的(Guo et al.,2018;Fan et al.,2020)。有研究表明,在草地生態系統中肥沃土壤的細菌含量是高于貧瘠土壤的,而這一變化主要是通過土壤中氮元素的調控所實現的(Bardgett et al,1997),草地退化引起的植被物種和生物量的減少,在一定程度上減少了凋落物和根源碳氮輸入到土壤中,進而影響土壤微生物基質的有效性(Wu et al.,2014),因此,認為草地退化引起的植被物種組成和生物量的減少會對微生物群落產生影響。另一方面,草地退化使得高寒草甸的優勢物種莎草科植被減少,這可能也是造成微生物變化的一個原因,已有文章報道植被物種對于微生物有選擇性效應(Haney et al.,2015)。Guo et al.(2015)研究發現在高寒草地土壤微生物群落結構主要是受控于溫度、土壤養分、水分和植被類型,Han et al.(2018)研究發現植被類型也是一個影響微生物群落結構變化的重要影響因子。但在高寒草地系統中,土壤原核生物與植被多樣性之間的相關性較弱,而與土壤異質性存在較強的相關性,草地退化導致土壤性質的變化,進而顯著影響土壤細菌群落,不同的微生物在生物地球化學循環中表現出不同的生態功能,而微生物群落結構可以顯著影響生態過程(Allison et al.,2013)。本研究結果表明隨著草甸的退化,土壤pH值會發生明顯變化,且驅動退化高寒草甸土壤細菌群落的主要環境因子是土壤pH值(圖5c)。這與其他研究結果一致,在多種土壤生態系統中pH值通常與細菌群落有顯著的相關性,當土壤pH值偏離中性時,地下微生物群落便會受到環境脅迫壓力,微生物多樣性也因環境脅迫的選擇而降低(Wang et al.,2017)。高寒草地退化過程中植被和土壤質地都會產生明顯的變化,這種變化在一定程度會對地下土壤細菌群落產生一定的影響。綜上可知,高寒草地退化不但引起地上植被群落和土壤質地的變化,也會顯著影響土壤微生物群落結構發生變化,因此探究高寒草地退化過程中土壤微生物群落活動變化機制對于草地恢復具有重要意義。
4 結論
草甸退化對地上植被群落結構、土壤理化性質和土壤細菌群落多樣性和結構都產生了顯著的影響。其中,草地退化降低了土壤全氮、有機質和電導率含量,增加了土壤pH值。另外,草地退化顯著降低了土壤細菌群落的α多樣性,并改變了細菌群落物種組成和群落結構。土壤異質性對土壤細菌群落有一定的影響,驅動土壤細菌群落變化的主要環境因子是土壤pH值、有機質含量和總氮含量。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
