陸地棉COI家族基因鑒定及在干旱和鹽脅迫下的表達分析
2022-06-14 01:02:06李美麗宿俊吉楊永林秦江鴻李鮮鮮楊德龍馬麒王彩香
中國農(nóng)業(yè)科技導報 2022年4期
關鍵詞:分析
李美麗, 宿俊吉, 楊永林, 秦江鴻, 李鮮鮮, 楊德龍*,馬麒, 王彩香*
(1.甘肅農(nóng)業(yè)大學生命科學技術學院,省部共建干旱生境作物學國家重點實驗室,蘭州 730070;2.石河子農(nóng)業(yè)科學研究院棉花研究所,新疆 石河子 832000;3.新疆農(nóng)墾科學院棉花研究所,新疆 石河子 832000)
茉莉素(jasmonates,JAs)是具有廣泛作用的植物激素,它不僅調(diào)控植物生長發(fā)育,而且還參與植物生物或非生物脅迫響應[1-2]。冠菌素不敏感蛋白1(coronatine-insensitive 1,COI1)是茉莉素信號途徑的關鍵受體,含有富含亮氨酸重復(leucinerich repeat,LRR)和F-box結(jié)構(gòu)域,最早從擬南芥(Arabidopsis thaliana)中獲得[3]。該蛋白能夠與SCF(skp-cullin-F-box)型 E3泛素連接酶組成SCFCOI1復合物,構(gòu)成茉莉素信號轉(zhuǎn)導途徑中的核心組分,能夠識別茉莉素信號傳導途徑中的一些調(diào)控因子,通過26S蛋白酶體降解,達到調(diào)控植物生長發(fā)育和抗逆性的作用[4]。在擬南芥中,COI1參與調(diào)控葉片衰老[5-8]和頂端優(yōu)勢[9]。在小麥中,TaCOI1參與小麥雄性不育信號調(diào)控[10]。此外,水稻OsCOI1的RNAi株系表現(xiàn)出株高、節(jié)間長度和籽粒長度的改變,并增加了對咀嚼昆蟲的敏感性[11],而油菜中沉默COI1基因表現(xiàn)出蚜蟲抗性下降和雄性不育等現(xiàn)象[12]。目前,關于COI1基因功能的研究主要集中在植物的生長發(fā)育和生物脅迫方面,而在植物非生物脅迫中的研究甚少。
隨著植物基因組信息不斷釋放,已從小立碗鮮、中華卷柏、云杉、水稻、高粱、毛果楊中分別鑒定到12、7、2、9、8、10個COI基因家族成員[13]。棉花不僅是天然纖維的重要來源,也是重要的抗旱、耐鹽堿先鋒作物,鑒定分析陸地棉(Gossypium hirsutum L.)COI1家族基因,為改良棉花耐逆性提供重要基因資源。本研究基于最新公布的升級版陸地棉TM-1參考基因組[14],對GhCOI家族成員進行全基因組鑒定,并對其家族成員的理化性質(zhì)、基因結(jié)構(gòu)、染色體分布、系統(tǒng)進化和共線性關系進行分析,結(jié)合干旱、鹽、高溫和冷脅迫處理轉(zhuǎn)錄組數(shù)據(jù),鑒定參與非生物脅迫的GhCOI基因,利用熒光定量PCR(qRT-PCR)驗證其脅迫表達模式,為陸地棉COI家族基因功能研究奠定基礎。
1 材料與方法
1.1 供試材料
本研究所用材料為抗旱、耐鹽堿陸地棉品種新石K18,種子由石河子農(nóng)業(yè)科學研究院棉花研究所提供。將新石K18棉花種子用75%乙醇浸泡2 min,用無菌水漂洗至無乙醇味后,再用無菌水浸泡24 h左右,將露白的棉花種子放置于自制水培盒(30 cm×20 cm×15 cm,內(nèi)置泡沫板24 cm×15 cm,11個孔,孔距7 cm)中進行萌發(fā),挑選長勢一致的棉花幼苗,移入1/2改良型霍蘭格營養(yǎng)液中進行培養(yǎng),待棉花幼苗長至4葉期分別用200 mmol·L-1NaCl和 15% PEG-6000 溶液進行脅迫處理,每個處理設置3個生物學重復,并分別在處理0、0.5、1.0、3.0、6.0、12.0和24.0 h進行葉片采樣,樣本收集后液氮速凍,-80℃保存?zhèn)溆谩?/p>
1.2 數(shù)據(jù)來源
棉花全蛋白序列、全基因組序列和基因注釋GFF3 文 件 下 載 自 CottonFGD[15](http://www.cottonfgd.org/),擬南芥COI蛋白序列下載自TAIR數(shù)據(jù)庫[16](https://www.arabidopsis.org/index.jsp),陸地棉4種非生物脅迫(鹽、干旱、冷和熱)轉(zhuǎn)錄組數(shù)據(jù)下載自NCBI SRA數(shù)據(jù)庫(https://www.ncbi.nlm.nih.gov/sra/?term=PRJNA248163)。
1.3 GhCOI基因家族鑒定及蛋白特性分析
根據(jù)擬南芥已鑒定出的COI基因及其編碼的蛋白序列,利用Pfam[17]數(shù)據(jù)庫(http://pfam.xfam.org/)和Hmmer3.0軟件構(gòu)建隱馬氏模型,對棉花全蛋白序列進行檢索和去冗余,得到候選蛋白序列,閾值設為e<1.0×10-5。利用在線軟件平臺PfamScan(https://www.ebi.ac.uk/Tools/pfa/pfamscan/) 和SMART[18](http://smart.embl-heidelberg.de/)對棉花所有候選基因的氨基酸序列結(jié)構(gòu)域進行鑒定,凡含有LRR和F-box結(jié)構(gòu)域的蛋白即為棉花COI基因家族成員,并根據(jù)陸地棉COI基因家族在染色體上的定位及親緣關系遠近進行命名。
利用在線工具ExPASy(https://web.expasy.org/protparam/)分析陸地棉GhCOI蛋白的理化性質(zhì),包括氨基酸數(shù)目(number of amino acids)、分子量(molecular weight)和等電點(pI)等;在亞細胞定位網(wǎng)站 WoLF PSORT(https://wolfpsort.hgc.jp/)對其進行亞細胞定位預測分析。
1.4 GhCOI家族基因染色體定位、系統(tǒng)進化樹構(gòu)建及共線性分析
GhCOI基因染色體信息從基因注釋文件中獲得,用MapInspect軟件繪制GhCOI家族基因在染色體上的位置;利用MEGA7.0軟件中的Align by Clustal W工具進行多重序列比對,各參數(shù)設定為默認值,采用Neighbor-Joining法構(gòu)建系統(tǒng)發(fā)育進化樹,Bootstrap值設定為1 000;采用TBtools軟件中One step MCScanX工具對GhCOI家族基因進行共線性及同義替換率/非同義替換率(Ka/Ks)分析。
1.5 GhCOI基因結(jié)構(gòu)和蛋白保守基序的分析
分別從陸地棉全基因組序列和基因注釋文件中篩選GhCOI基因家族的DNA和CDS序列,利用Gene Structure Display Server(GSDS)(http://gsds.gao-lab.org/index.php)對GhCOI家族基因進行基因結(jié)構(gòu)分析。利用MEME Suite 5.3.3(https://meme-suite.org/meme/)對GhCOI蛋白進行保守基序預測,motif數(shù)為5個,借助TBtools軟件對該基因家族的保守基序進行可視化分析。
1.6 GhCOI家族基因順式作用元件分析
利用TBtools軟件Gtf/Gff3 Sequences extractor工具提取陸地棉CDS序列上游2 000 bp序列,通過 PlantCare[19]數(shù)據(jù)庫(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)預測順式作用元件。
1.7 GhCOI家族基因表達特征分析
GhCOI家族基因的鹽、干旱、冷和熱脅迫的轉(zhuǎn)錄組數(shù)據(jù)從NCBI SRA數(shù)據(jù)庫(https://www.ncbi.nlm.nih.gov/sra/?term=PRJNA248163)下載,利用基迪奧生物信息繪圖云平臺(https://www.omicshare.com/tools/)繪制該家族基因的非生物脅迫表達量熱圖。
1.8 GhCOI家族基因qRT-PCR驗證
篩選含抗逆順式作用元件且高表達的GhCOI家族基因,利用qRT-PCR進行驗證。利用NCBI Primer-BLAST工具(www.ncbi.nlm.nih.gov/tools/primer-blast/)設計qPCR引物(表1),產(chǎn)物片段大小在70~150 bp之間,由上海生工生物工程技術服務有限公司合成。將1.1中所采集的樣品用多糖多酚總RNA提取試劑盒(天根生化科技有限公司)提取總RNA,超微量濃度檢測儀和瓊脂糖凝膠電泳檢測其純度、含量和完整性。采用FastKing gDNA Dispelling RT SuperMix試劑盒(天根生化科技有限公司)合成cDNA。采用SuperReal熒光定量預混試劑增強版試劑盒(天根生化科技有限公司)進行qRT-PCR,用2-ΔΔCT法[20]計算相對表達量。
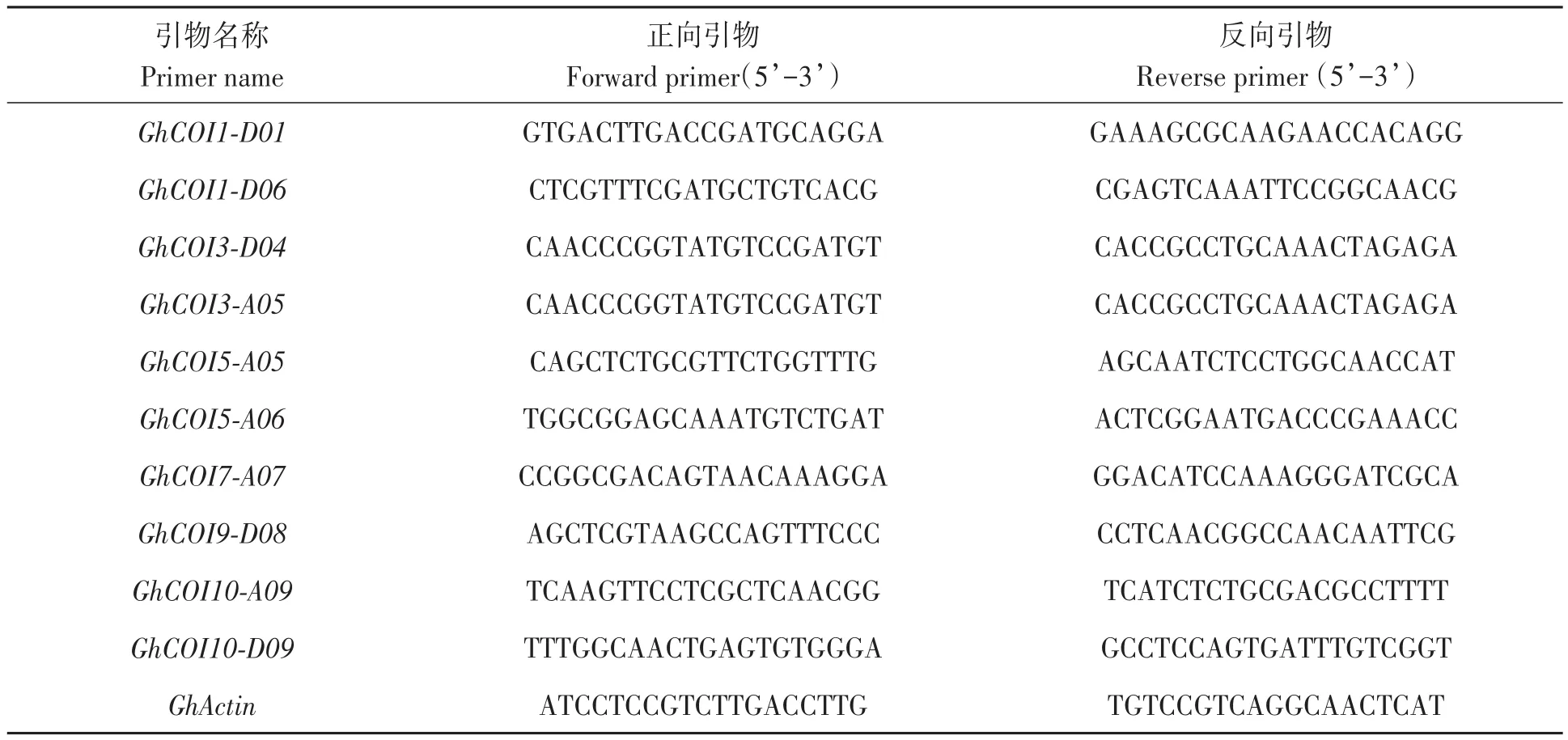
表1 實時熒光定量PCR引物Table 1 Primer sequence for qRT-PCR
2 結(jié)果與分析
2.1 GhCOI基因家族鑒定及理化性質(zhì)分析
從陸地棉中鑒定得到48個COI基因,根據(jù)陸地棉COI基因在染色體上的位置和進化關系將其命名,如表2所示。序列分析(表2)顯示,48個GhCOI基因編碼的蛋白長度為362(GhCOI4-A05)~702 aa(GhCOI6-A12和GhCOI6-D12);蛋白質(zhì)分子量在 39.99(GhCOI4-A05)~79.13 kD(GhCOI6-A12)之間;等電點分布在5.50(GhCOI8C-A08)~8.86(GhCOI1-A05、GhCOI1-D06)之間。
對48個GhCOI基因進行亞細胞定位預測,發(fā)現(xiàn)該家族基因主要定位在細胞質(zhì)、細胞核、葉綠體和線粒體(表2),其中43.75%的GhCOI基因定位在細胞核(21個),39.58%的GhCOI基因定位在細胞質(zhì)中(19個),其他基因則定位在葉綠體或線粒體,表明GhCOI基因主要在細胞質(zhì)和細胞核中表達。
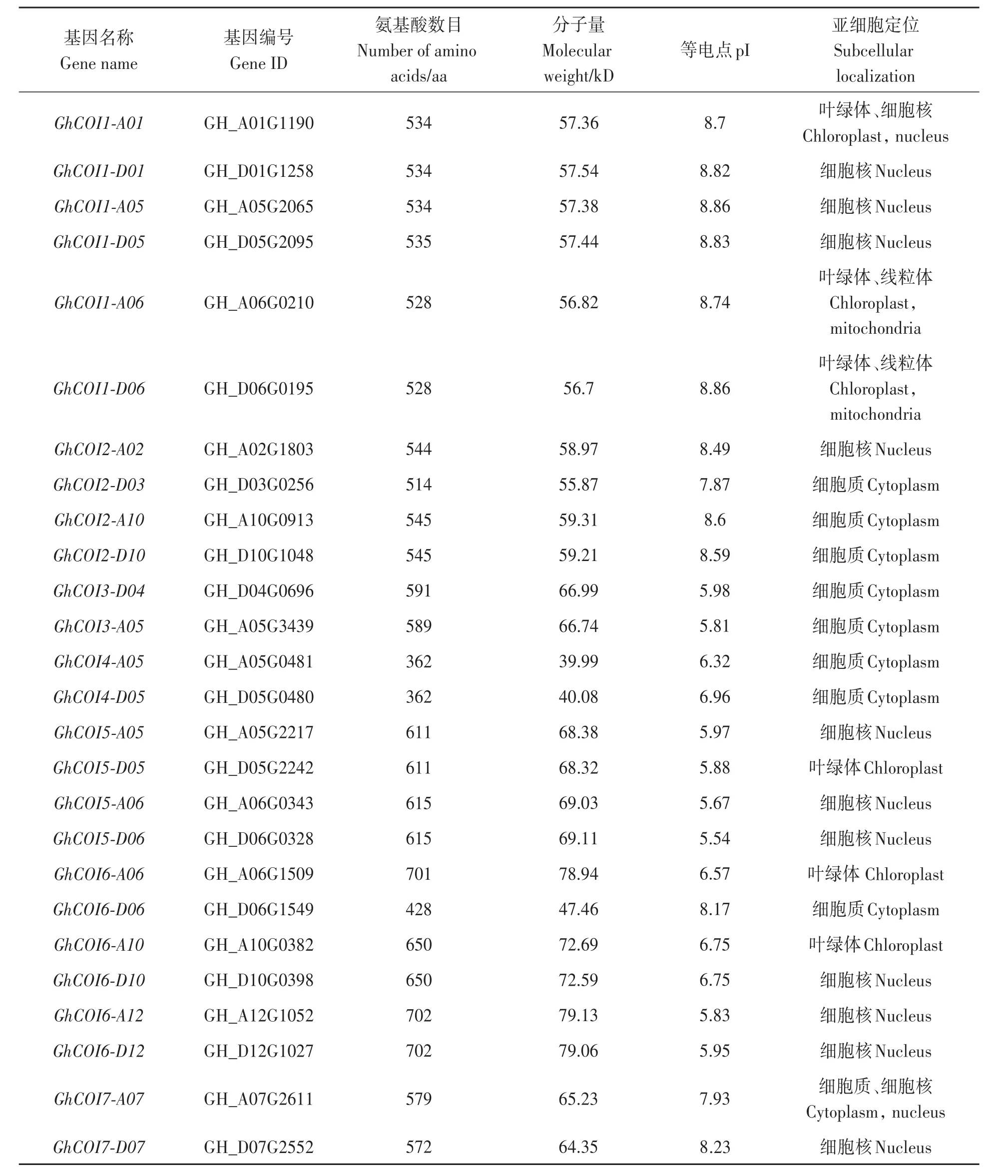
表2 GhCOI基因理化性質(zhì)及亞細胞定位預測Table 2 Characteristics and subcellular localization prediction of GhCOI genes

表2 GhCOI基因理化性質(zhì)及亞細胞定位預測Table 2 Characteristics and subcellular localization prediction of GhCOI genes 續(xù)表Continuted
2.2 GhCOI基因的染色體定位
對48個GhCOI基因進行染色體定位,結(jié)果(圖1)表明:GhCOI基因在染色體上呈不均勻分布,其中染色體 A03、A04、A13、D02和 D13無GhCOI基因分布,染色體 A01、A02、D03、A07、A09、D01、D04、D07和D09各有1個基因分布,其他染色體上GhCOI基因分布數(shù)目2~4個不等,此外還發(fā)現(xiàn)有42%的GhCOI基因主要分布在染色體的兩端。
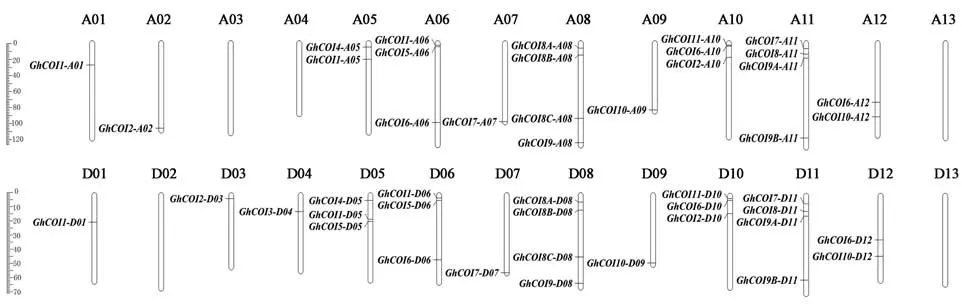
圖1 GhCOI基因在染色體上的分布Fig.1 Chromosome distribution of GhCOI genes
2.3 GhCOI基因的系統(tǒng)發(fā)育和共線性分析
2.3.1 系統(tǒng)發(fā)育分析 為了研究陸地棉與其他3個棉種及擬南芥COI基因的系統(tǒng)發(fā)育關系,分別從 海 島 棉(Gossypium barbadenseL.)、亞 洲棉(Gossypium arboreumL.)和雷蒙德氏棉(Gossypium raimondiiL.)得到 48、25和 23個COI基因,運用MEGA7.0軟件進行聚類分析,結(jié)果如圖2所示。根據(jù)系統(tǒng)發(fā)育樹中親緣關系的遠近,151個COI基因分為3個亞族:Ⅰ、Ⅱ和Ⅲ,而Ⅰ和Ⅱ亞族又分別可細分為5個不同的亞類,即Ⅰ-1、Ⅰ-2、Ⅰ-3、Ⅰ-4、Ⅰ-5和Ⅱ-1、Ⅱ-2、Ⅱ-3、Ⅱ-4、Ⅱ-5,且不同亞族和亞類所含物種種類和COI基因數(shù)量存在一定差異。其中,Ⅰ-3亞類、Ⅱ亞族和Ⅲ亞族均不包含AtCOI基因,且Ⅰ-3和Ⅱ-1亞類包含成員最少(6個),Ⅰ-5成員最多(24個),其他亞類所包含成員數(shù)量11~18個不等。
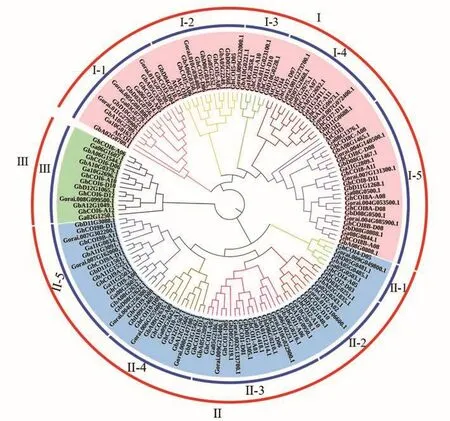
圖2 COI基因系統(tǒng)發(fā)育分析Fig.2 Phylogeny analysis of COI genes
2.3.2 共線性分析 為了更好地了解GhCOI基因的擴展模式,將GhCOI基因進行了共線性分析,結(jié)果(圖3)發(fā)現(xiàn):部分GhCOI基因同時與家族內(nèi)多個基因存在共線性關系,如GhCOI1-A01與GhCOI1-D01和GhCOI1-D05之間有共線性關系,因此共發(fā)現(xiàn)62對基因具有共線性關系;48個GhCOI基因全為片段復制,這表明片段復制對該家族的擴大起到了關鍵作用。
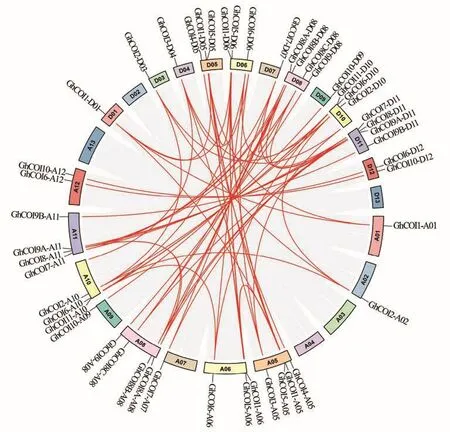
圖3 COI基因系統(tǒng)共線性分析Fig.3 Collinearity analysis of COI genes
GhCOI家族基因復制事件的Ka/Ks范圍為0.09~0.65,平均值是0.21(數(shù)據(jù)未列出),所有重復事件GhCOI基因的Ka/Ks值均小于1,說明這些基因均在純化選擇壓的作用下進化。
2.4 GhCOI基因結(jié)構(gòu)及蛋白保守域的分析
對GhCOI基因的結(jié)構(gòu)進行分析(圖4A),發(fā)現(xiàn)21個GhCOI基因無內(nèi)含子,內(nèi)含子數(shù)最多的基因GhCOI10-A09含有8個內(nèi)含子,其余基因所含內(nèi)含子數(shù)目1~7個不等。內(nèi)含子的不均勻分布表明,GhCOI家族基因在進化的過程中可能發(fā)生內(nèi)含子的插入或缺失。
對GhCOI蛋白的保守基序進行預測(圖4B),發(fā)現(xiàn)48個GhCOI蛋白均含Motif 1和Motif 3,除GhCOI4-A05和GhCOI4-D05外,其余均含有Motif 4,其序列如圖4C所示;另外,41.7%(20個)的蛋白含有的Motif結(jié)構(gòu)最多(5個),4.2%(2個)的蛋白含有的Motif結(jié)構(gòu)最少(Motif 1和Motif 3)。
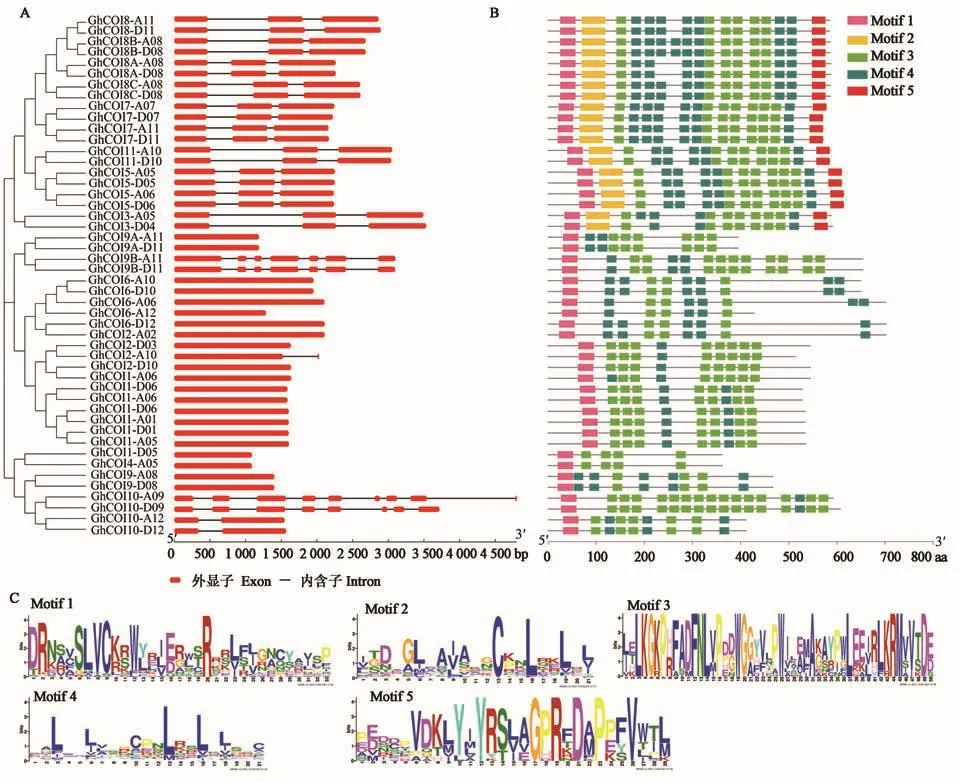
圖4 GhCOI家族成員結(jié)構(gòu)特征和保守基序分析Fig.4 Structure and conservative motif analysis of the GhCOIs
2.5 GhCOI基因非生物脅迫相關順式作用元件預測和表達分析
對48個GhCOI基因上游2.0 kb的啟動子序列順式作用元件進行分析(圖5A),在GhCOI啟動子區(qū)域發(fā)現(xiàn)了6種脅迫響應調(diào)控元件,即富含 TC 的重復序列(TC-rich repeats)、LTR(lowtemperature re-sponsiveness),GC 基序/ARE(GC-motif/anaerobic induction)、MBS(MYB binding site)和WUN基序(wound element),它們分別與防御/應激、低溫、缺氧、干旱和創(chuàng)傷反應相關。不同的GhCOI基因啟動子中存在不同類型和數(shù)量的調(diào)控元件,表明GhCOI基因可能通過多個順式作用元件共同參與植物的非生物脅迫響應。
對鹽、干旱、冷和熱4種脅迫下的48個GhCOI家族基因的表達模式進行分析,結(jié)果(圖5B和5C)發(fā)現(xiàn):該家族基因在NaCl、PEG、熱和冷脅迫下,分別發(fā)現(xiàn)有66.7%、56.25%、68.8%和27.1%的基因表達量升高,其中,同時響應NaCl和PEG脅迫的高表達基因有20個,同時響應PEG和熱脅迫的有25個。
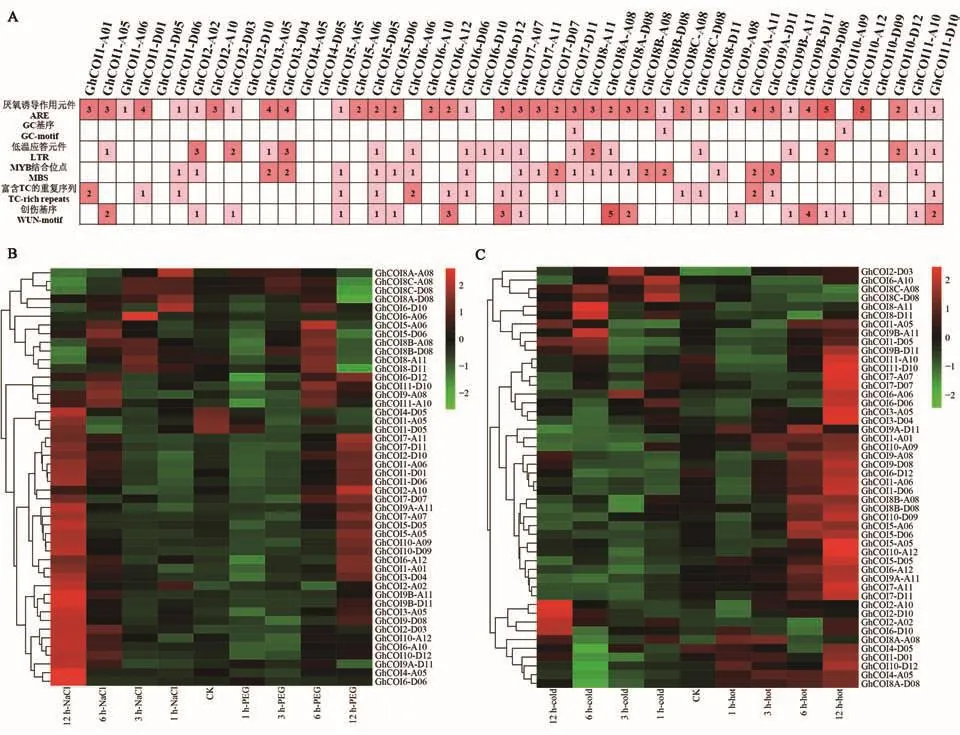
圖5 GhCOI基因順式作用元件預測和在非生物脅迫下的表達Fig.5 cis-acting elements and expression analysis of GhCOI genes
2.6 GhCOI家族基因qRT?PCR分析
對含較多抗逆順式作用元件且同時在PEG和NaCl脅迫下高表達的20個基因進行分析,選出表達較高的9個基因進行qRT-PCR驗證,結(jié)果如圖6所示。NaCl和PEG均能顯著誘導GhCOI基因的表達,但不同基因的響應程度存在差異。在NaCl處 理 下(圖 6A),GhCOI1-D06、GhCOI3-D04、GhCOI3-A05、GhCOI5-A06和GhCOI10-A09基因表達量顯著提高,其中GhCOI5-A06的表達量最高,GhCOI3-A05次之。在PEG脅迫下(圖6B),除GhCOI1-D01基因外,其余8個基因均呈現(xiàn)高表達,其中GhCOI1-D01、GhCOI1-D06、GhCOI3-D04、GhCOI3-A05、GhCOI5-A05和 GhCOI5-A06基因的表達量均呈現(xiàn)先增后減再上升的趨勢,且GhCOI5-A05表達量最高,GhCOI5-A06次之。由此得出,GhCOI5-A06能同時強烈響應NaCl和PEG兩種脅迫。
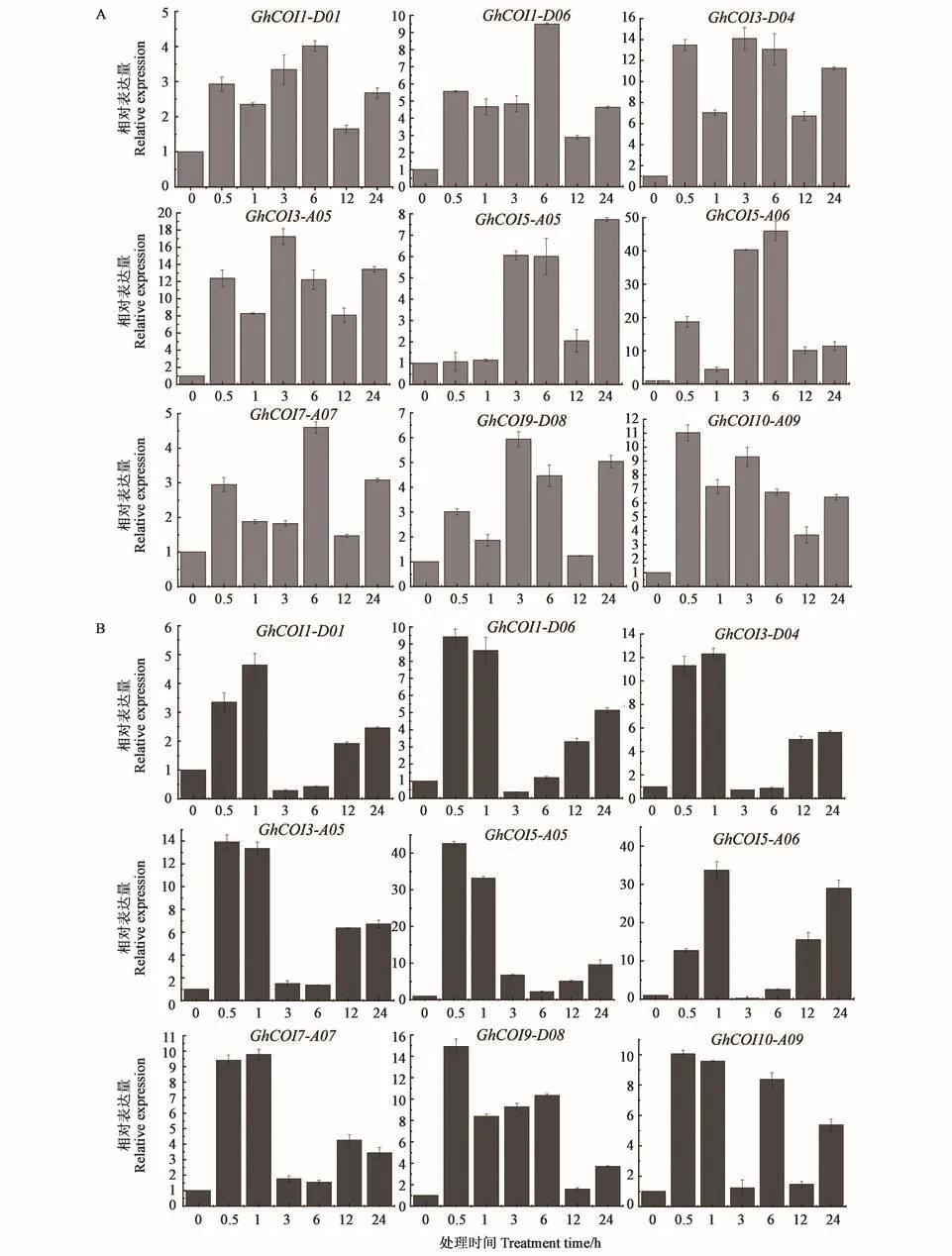
圖6 9個GhCOI基因在NaCl和PEG脅迫下的表達分析Fig.6 Expression analysis of 9 GhCOIs under NaCl and PEG stress
總體來看,GhCOI基因在2種非生物脅迫下的相對表達量都明顯上調(diào),與SRA數(shù)據(jù)庫中陸地棉TM-1在鹽和干旱脅迫下的轉(zhuǎn)錄組數(shù)據(jù)的表達趨勢相吻合。
3 討論
COI1是茉莉酸信號轉(zhuǎn)導途徑的關鍵調(diào)控因子,在植物調(diào)控生長發(fā)育和響應逆境脅迫方面起重要作用[21]。本研究共鑒定到陸地棉、海島棉、亞洲棉和雷蒙德氏棉COI基因各48、48、25和23個,前人在擬南芥中鑒定到7個COI基因[13]。擬南芥與棉花COI基因數(shù)量的差異表明,隨著植物倍性的增加,COI家族基因也在不斷的擴增和復制。系統(tǒng)發(fā)育分析發(fā)現(xiàn),151個GhCOI蛋白被劃分為3個亞族,且擬南芥COI基因主要聚集在Ⅰ亞族中,其中AtCOI1與其他6個成員有明顯的聚類分歧,具體表現(xiàn)為Ⅰ-1亞家族中只有AtCOI1,這與段龍飛[13]的研究結(jié)果一致。值得注意的是,AtCOI1是茉莉酸調(diào)節(jié)途徑的關鍵受體基因,GhCOI3-A05和GhCOI3-D04與其聚類進化關系較近,這說明在陸地棉中GhCOI3-A05和GhCOI3-D04可能是參與茉莉酸轉(zhuǎn)導途徑的關鍵候選基因。根據(jù)基因結(jié)構(gòu)分析發(fā)現(xiàn),陸地棉中有44%的COI基因無內(nèi)含子,其余基因含有1~7個不等,這種內(nèi)含子數(shù)量的差異可能是在進化過程中,由于內(nèi)含子的插入或丟失造成的變化;保守基序分析發(fā)現(xiàn)48個GhCOI基因均含有Motif 1和Motif 3,推測它們?yōu)樵摷易宓谋J匦曰颉?/p>
基因復制被認為是基因組和遺傳系統(tǒng)進化的主要驅(qū)動力之一[22],片段復制、串聯(lián)復制和轉(zhuǎn)位事件被認為是3種主要的進化模式。在這些模式中,片段復制和串聯(lián)復制被認為是植物基因家族擴大的兩個主要原因[23]。對陸地棉COI基因共線性分析發(fā)現(xiàn),48個GhCOI基因不均勻的分布于21條染色體上,且均為片段復制,其結(jié)果與小麥COI[24]家族基因共線性分析結(jié)果一致。基因在染色體上的這種不均勻分布表明基因在進化過程中存在遺傳變異[25]。此外,同義替換率和非同義替換率是評價重復事件純化選擇壓力的基礎,而純化選擇可能是重復基因功能差異的主要驅(qū)動力[26]。通過對復制基因?qū)Φ耐x替換率和非同義替換率分析發(fā)現(xiàn),GhCOI基因均在純化選擇壓下進行,這可能是促進COI基因能夠響應各種逆境脅迫反應的關鍵。
分析GhCOI基因在不同脅迫下的順式作用調(diào)控元件和表達譜。在GhCOI啟動子中發(fā)現(xiàn)了6種響應脅迫及防御反應調(diào)控元件(低溫、干旱、缺氧、防御和創(chuàng)傷),與Bai等[24]在小麥TaCOI家族基因的研究一致,這表明GhCOI和TaCOI基因一樣,對不同的脅迫處理和損傷有不同的調(diào)控機制。本文進一步研究了GhCOI基因在不同脅迫條件下的表達模式,結(jié)果顯示該家族基因可響應多種非生物脅迫。例如,在干旱和溫度誘導下,GhCOI3-D04、GhCOI3-A05和GhCOI7-A07表達上調(diào),且這些基因均含有1~3個MBS/LTR元件。COI同源基因在不同非生物脅迫下的表達可為該家族的生理生化和基因功能研究提供重要信息。
本研究利用qRT-PCR對部分基因進行表達驗證發(fā)現(xiàn),9個被檢測GhCOI基因在NaCl和PEG脅迫下均上調(diào)表達,其中GhCOI5-A06是能同時響應2種脅迫最強烈的基因,其表達量在NaCl脅迫下是對照的45.96倍,在PEG-6000脅迫下是對照的33.71倍。段龍飛[13]對擬南芥COI基因響應鹽和干旱脅迫的研究顯示:擬南芥COI基因表達模式不同于棉花COI基因,如AtCOI4在2種脅迫中表達量均量下調(diào)趨勢,而AtCOI5在鹽脅迫下表達量下調(diào),在干旱脅迫下表達量上調(diào)。這可能是不同植物在自然界所處生境不同,它們在各自進化過程中一些信號途徑的某些節(jié)點可能發(fā)生了分歧。由此推測,陸地棉GhCOI基因家族在進化過程中,為適應外界環(huán)境變化而分化出不同的家族成員[27]。
綜上所述,在基因進化過程中,親緣關系較近的基因結(jié)構(gòu)和保守基序數(shù)量、類型都極為相似,且均有共線性關系,而基因復制為基因家族的擴增提供了原動力,并對不同基因執(zhí)行同一功能提供了可能性。GhCOI基因的系統(tǒng)發(fā)育和表達模式將為基因功能提供更全面的理解,為陸地棉GhCOI家族基因響應非生物脅迫調(diào)控機制的研究奠定了基礎,為棉花抗逆育種提供了潛在的基因資源。今后還需進一步深入研究GhCOI基因如何發(fā)揮特殊功能。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
當代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
