抗食品腐敗真菌的乳酸菌篩選、復配及混合發酵工藝優化
2022-06-11 11:19:00杜瑩瑞李市場馮騰柱張亞娟龔明貴
食品工業科技 2022年12期
杜瑩瑞,李市場,馮騰柱,張亞娟,陳 妍,龔明貴
(河南科技大學食品與生物工程學院,河南洛陽 471023)
在食品工業中,真菌污染及其毒素是造成經濟損失、危害消費者健康的重要原因[1]。真菌及毒素大部分具有高毒及致癌性,極易在各類食品中存在,嚴重影響食品的安全和消費者健康[2]。食品腐敗真菌在市場上大多采用食品添加劑進行食品防腐,而添加劑屬于化學物品且大多都是國家禁止使用的[3]。因此,對抗食品腐敗真菌混合菌的篩選及發酵工藝的優化是迫切需要的。
乳酸菌和丙酸桿菌單獨發酵的發酵液具有一定的抗真菌活性[4-7],但活性較低。目前有許多報道乳酸菌和丙酸桿菌抗真菌活性的研究與應用,包括菌株篩選方法的優化與創新,發酵產物中抗真菌成分的前體物質的添加以及整個發酵過程的優化等。如Garnier等[8]通過模擬奶酪的方法對奶酪中具有抗真菌活性的菌株進行篩選,文中介紹了24孔板法對干酪中乳桿菌的篩選及測定其抗真菌活性,結果發現不同乳酸菌對真菌抑制的種類和效果有所差異。實驗所用的幾種干酪乳桿菌的含菌培養物均顯示出抗真菌活性,但抑制的真菌種類和抑制效果不盡相同。;并且Valerio等[9]通過添加乳酸菌抗真菌代謝物中苯乳酸的前體物質苯丙氨酸提高了乳酸菌的抗真菌活性,文中采用96孔板進行微孔稀釋試驗測定抗真菌活性,結果表明,乳酸菌顯示的抗真菌活性可以通過向生長培養基中加入苯丙酮酸來改善和優化,并且它是存在幾種以協同方式起作用的代謝物的結果;Zhang等[10]通過Lactobacillus buchneri和Lactobacillus diolivorans在酸面團中共同發酵產生丙酸來達到抗真菌效果,文中使用瓊脂孔擴散評估乳桿菌發酵的酸面團的抗真菌活性,以期可以通過添加酸面團來代替化學品的使用。因此,本文從多株乳酸菌和丙酸桿菌菌株中篩選出抗食品腐敗真菌活性較高菌株[11-12],通過一系列方法篩選抗食品腐敗真菌菌株最佳組合[13],從混合菌發酵角度出發,優化混合菌發酵條件以最大限度提升植物乳桿菌L9和費氏丙酸桿菌D5菌株組合的抗真菌效果,解決單菌抗真菌活性低、減少市場食品浪費問題[14-16]。
1 材料與方法
1.1 材料與儀器
費氏丙酸桿菌(Propionibacterium freudenreichii)D4、D5和乳酸乳球菌(Lactococcus lactis) L1~L6、L8、L9菌株、沃納利葡萄球菌(Staphylococcus warneri)L7 由上海市華東師范大學微生物實驗室保藏;植物乳桿菌(Lactobacillus plantarum) MRE3013、MRE3014、MRE3035菌株 從羅非魚腸道中分離得到,由中國海洋大學食品學院贈予;費氏丙酸桿菌(Propionibacterium freudenreichiiATCC 9614) D1、D2 美國ATCC菌種保藏中心;指示菌 面包腐敗真菌(黑曲霉、黃曲霉、桔青霉、枝孢霉、交鏈孢霉);MRS培養基、SLB培養基、ME培養基、MESP培養基、TGYE培養基 北京奧博星生物技術有限責任公司;PDA培養基 國藥集團化學試劑有限公司;M17培養基 北京陸橋技術股份有限公司;WFH培養基 20%高筋小麥粉(w/v)在室溫下于磁力攪拌器中1200 r/min攪拌4 h,于冷凍離心機中離心(4000 r/min,10 min,4 ℃)獲得上清液,每升上清液中加入葡萄糖、麥芽糖、蔗糖和果糖各15 g,以及10 g酵母浸粉和7 g瓊脂,pH6.8±0.1,106 ℃,滅菌20 min;脫脂乳粉培養基 40 g/L脫脂乳粉,pH7.0±0.1,115 ℃,滅菌30 min。
HCB-900V超凈工作臺 青島海爾生物醫療股份有限公司;CR22N大容量高速冷凍離心機 Hitachi Koki Holdings Co., Ltd;9203A電熱恒溫鼓風干燥箱上海躍進醫療器械廠;Synergy HT酶標儀BioTek Instruments,Inc.;T18 ULTRA-URRAX均質機 艾卡(廣州)儀器設備有限公司;IKA lab dancer漩渦振蕩器 艾卡(廣州)儀器設備有限公司;DELTA 320 pH計 梅特勒-托利多國際有限公司。
1.2 實驗方法
1.2.1 單一抗真菌性強的菌株篩選 首先從L1-L9、D1、D2、D4、D5、MRE3013、MRE3014和MRE303516株菌株中篩選出抗真菌性較強的單一菌株,再進行篩選較強抗真菌性的菌株組合。
將各株乳酸菌和丙酸桿菌分別接種到 MRS和SLB液體培養基中,置于30 ℃的恒溫搖床中,以180 r/min轉速進行培養2 d,制備種子液。配制MRS瓊脂培養基于24孔板中,用10 μL規格的一次性接種環沾取各菌株的種子液,在瓊脂平板中進行接種,在無菌層流空氣將其吹干后,放置在厭氧產氣袋中,在30 ℃培養箱中培養48 h[17-19]。制備指示霉菌孢子懸液,并用血球計數板計數[20]孢子懸液的孢子濃度,并將孢子懸液用無菌水稀釋到1×105個孢子/mL,4 ℃冰箱保藏備用。配制軟瓊脂,高壓蒸汽滅菌后在55 ℃水浴鍋中保溫備用。將1×105個孢子/mL的5種霉菌孢子懸液分別用軟瓊脂10倍稀釋,振蕩搖勻,制備含1×104個孢子/mL霉菌孢子的軟瓊脂[21]。將5 mL 1×104個孢子/mL的5種軟瓊脂分別加入到培養48 h的各菌株MRS瓊脂平板中進行覆蓋。待軟瓊脂凝固后,將平板放入28 ℃的培養箱中有氧培養,觀察記錄各孔霉菌的生長情況。
1.2.2 24孔板法篩選復合菌株協同抗真菌組合 將各株抗真菌乳酸菌和丙酸桿菌分別接種到 MRS 肉湯和SLB培養基中,在30 ℃條件下分別培養24和48 h制備種子培養液。將種子培養液離心(5000×g,10 min,4 ℃),并用無菌生理鹽水重懸振蕩進行洗滌,洗滌2次后重懸于十分之一體積的生理鹽水中,將菌懸液稀釋,取樣測定OD600,緩慢加入生理鹽水稀釋,直到稀釋到OD600=0.5,兩個菌大小形態相似,所以對應的OD600也幾乎一樣,以此為比例進行稀釋。記錄這個稀釋濃度,在超凈臺里以同樣濃度稀釋,兩個菌都是稀釋到OD600=0.5制備菌懸液。
將制備好的菌懸液各50 μL加入到含乳酸菌和丙酸桿菌混菌瓊脂培養基的24孔板中,設計陰性對照(加入菌懸液100 μL)和陽性對照組(加入無菌生理鹽水100 μL),然后加入用 1 mL融化的小麥粉水解物(WFH)瓊脂培養基(45 ℃水浴鍋保溫),與之前加入的菌液搖晃混合混勻,待瓊脂凝固后置于 30 ℃培養箱中培養18 h[22]。 培養18 h后,在24孔板各孔中覆蓋100 μL 含104個指示真菌孢子/mL 的軟瓊脂,28 ℃培養,每24 h觀察乳酸菌/丙酸桿菌組合對指示霉菌的抑制情況并記錄[23]。
1.2.3 分級抑菌濃度(Fractional inhibitory concentration,FIC)法進一步篩選復合菌協同抗真菌組合 制備1.2.2步驟中篩選得到的菌株的OD600=0.5的菌懸液進一步篩選復合菌協同抗真菌作用。將菌液用無菌的生理鹽水進行10倍梯度稀釋,乳酸菌稀釋為10、10-1、10-2、10-3倍原濃度的菌懸液,10、10-1、10-2、10-3、10-4倍原濃度的菌懸液。24孔板為4×6排列的小孔,除了最后一縱列作為空白對照(100μL無菌生理鹽水)外,剩下4×5格。將4種梯度濃度的乳酸菌菌懸液從上到下每行濃度依次遞減加入,每孔50 μL;將5種梯度濃度的丙酸桿菌菌懸液從左到右丙酸菌每行濃度依次遞減加入,每孔50 μL,然后加入用1 mL融化的小麥粉水解物(WFH)瓊脂培養基(45℃水浴鍋保溫),與之前加入的菌液搖晃混合混勻,待瓊脂凝固后置于30 ℃培養箱中培養18 h。然后在24孔板各孔中覆蓋100 μL 含104個指示真菌孢子/mL的軟瓊脂,28 ℃培養,每24 h觀察乳酸菌/丙酸桿菌組合對指示霉菌的抑制情況并記錄。
1.2.4 混合菌株發酵工藝優化 在上述的試驗中,篩選出了植物乳桿菌L9和費氏丙酸桿菌D5作為后續混合發酵中的抗真菌菌株組合。因此,我們將對其發酵工藝進行優化,確定最佳發酵培養基和發酵條件,以提高協同抗真菌能力[24]。
1.2.4.1 Plackett-Burman試驗 研究發現有12個因素對植物乳桿菌L9和費氏丙酸桿菌D5有影響,它們分別是葡萄糖(A)、碳酸鈣(B)、酵母浸粉(C)、酪蛋白胨(D)、磷酸氫二鉀(E)、硫酸鎂(F)、硫酸錳(G)、氯化鈉(H)、接種比(D5:L9)(J)、培養溫度(K)、乙酸鈉(L)、檸檬酸銨(M)[25]。通過Plackett-Burman試驗對以上12個因素進行研究,每個因素分別選取兩個水平:低水平“-1”和高水平“+1”,響應值為發酵液上清液的抗真菌活性。表1列出了因素的編碼及水平,X1~X12分別編碼了12個因素。
Plackett-Burman試驗設計表如表1所示,按照設計表中的順序配制培養基、調整接種量和發酵溫度,發酵4d后做96孔板試驗,用酶標儀測定[26]并計算發酵液上清液的抗真菌活性,在表中最右列列出,96孔板操作和抗真菌活性計算公式如下:

表1 Plackett-Burmen實驗設計水平范圍Table 1 Range of different factors investigated of Plackett-Burman design
將制得的無細胞上清液加入到96孔板的孔中,每孔添加量為100 μL;將無菌的ME肉湯以每孔100 μL的量加入到后續的孔中,對無細胞上清液進行連續的2倍濃度梯度稀釋,獲得一系列的無細胞上清液稀釋液(至少3個平行對照),以不含無細胞上清液的ME肉湯作為陰性對照;然后在稀釋液中加入100 μL 含1×104個孢子/mL的霉菌孢子ME肉湯(陽性對照加入無霉菌孢子肉湯),放入28 ℃的恒溫恒濕(80% RH)培養箱中培養48 h。
霉菌生長受無細胞上清液抑制的程度通過測定96孔板中霉菌在620 nm波長處的吸光度確定。細胞上清液的抗真菌活性以AU為單位定量,其表示為霉菌生長抑制率為50%的情況下,對應的無細胞上清液的濃度的倒數。其中生長抑制率(Ifungal)的計算公式為:

其中,Ifungal為對霉菌的生長抑制率;At為實驗組孔中霉菌的吸光度;Apc為陽性對照(無霉菌生長)孔的吸光度;Anc為陰性對照孔中霉菌的吸光度。
通過線性插值法確定Ifungal=50%時無細胞上清液的濃度(IC50),公式如下:
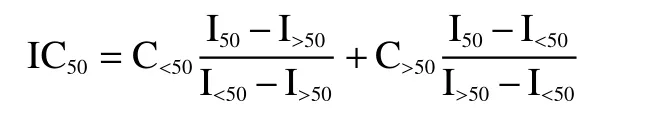
其中IC50為無細胞上清液在Ifungal=50%時的濃度;C<50為無細胞上清液在Ifungal<50%孔中的濃度;C>50為無細胞上清液在Ifungal>50%孔中的濃度;I>50為無細胞上清液在Ifungal>50%孔中的生長抑制率;I<50為無細胞上清液在Ifungal<50%孔中的生長抑制率;I50=50%。
于是抗真菌活性(Aantifungal)的計算公式為:

1.2.4.2 最陡爬坡試驗 影響植物乳桿菌L9和費氏丙酸桿菌D5菌株發酵液抗真菌活性的三個關鍵因素為酵母浸粉(C)、葡萄糖(A)、碳酸鈣(B);最終得到三個主要因素的回歸方程:抗真菌活性(AU)=-26.7+0.834A-0.518B+0.268C,從回歸方程中可以看出,葡萄糖(A)和酵母浸粉(C)對抗真菌活性有正效應,碳酸鈣(B)則對其有負效應。以發酵液上清液的抗真菌活性作為響應值,對3個關鍵因素進行最陡爬坡試驗,試驗設計如表2所示。
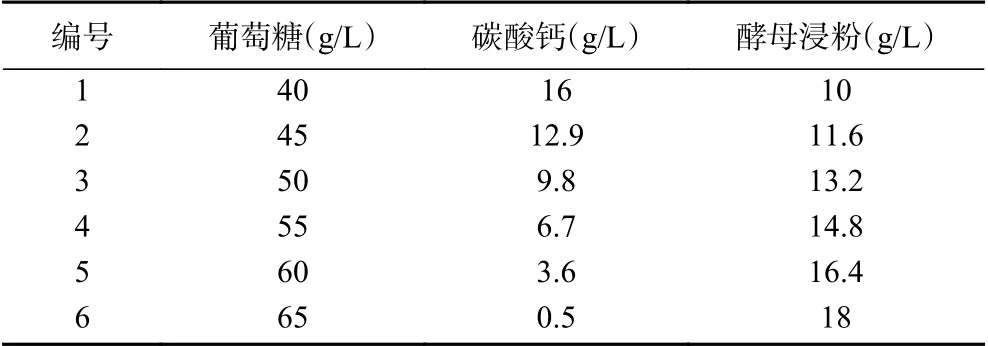
表2 最陡爬坡試驗設計Table 2 Design of the steepest climbing test
1.3 數據處理
各項數據均平行三次,用Word和Graphpadprism8作圖,通過Minitab 18軟件進行分析,得到試驗方差分析表比較組間差異的顯著性,通過Design Expert 12軟件對數據進行多元回歸擬合,對得到的回歸模型進行方差分析。
2 結果與分析
2.1 單菌菌株對5種霉菌的抗真菌效果
測定單菌菌株對5種霉菌的抗真菌效果,在24孔板中用雙層瓊脂覆蓋法評估了單菌菌株對5種霉菌指示菌的抗真菌效果,而桔青霉、黃曲霉是前期所試驗面包中污染霉菌出現頻率最高的兩種霉菌,因此篩選主要對這兩種菌抑制效果較強的抗真菌菌株。經研究發現有6株菌株對桔青霉、黃曲霉的抑制效果比較明顯,其中L7為沃納利葡萄球菌,L8、L9為乳酸乳球菌,3103、3014、3035為植物乳桿菌。從圖1可以看出,L9菌株和3103菌株對桔青霉、黃曲霉的抑制效果最強,其次為3035菌株、L8菌株、L7菌株和3014菌株。所有菌株對5種霉菌的抗真菌效果如表1所示。鑒于L7是不屬于《可用于食品的菌種名單》的菌種,所以后續試驗中不再對它的抗真菌活性進行進一步研究。
從圖1和表3中可以看出,在15株抗真菌菌株中,編號為L8、L9的乳酸乳球菌和編號為3103、3014、3035的植物乳桿菌這5株菌株對面包中污染霉菌出現頻率最高的兩種霉菌桔青霉和黃曲霉有較強的抗真菌效果;丙酸桿菌屬菌株對霉菌的抑制效果要整體強于乳酸菌組。但乳酸菌中也有個別對5種霉菌抑制能力較強的菌株,如L1、L9、3013和3035菌株。而且霉菌對于抗真菌菌株的敏感性也有差異,在PDA平板中加入不同添加量的發酵液后,枝孢霉和鏈格孢霉在較低的發酵液添加量下被抑制,在96孔板試驗下,也是在較低的濃度下被抑制,而黑曲霉、黃曲霉和桔青霉在稍高的發酵液添加量下才被抑制。因此得出黑曲霉、黃曲霉和桔青霉的生長不容易被抗真菌菌株所抑制,而枝孢霉和交鏈孢霉的生長較易被抑制。

圖 1 6株菌株對桔青霉Q05(A)和黃曲霉Y03(B)的抑制效果Fig.1 Inhibitory effects of 6 strains on Penicillium citrinum Q05 (A) and Aspergillus flavus Y03 (B)

表3 各抗真菌菌株對5種霉菌的抑制效果Table 3 Inhibitory activity of antifungal strains against 5 kinds of molds
2.2 復合菌協同作用對真菌的影響
由前期實驗表明,選擇黃曲霉作為指示菌。對所有乳酸菌和丙酸桿菌懸液進行處理,取樣測定OD600值,加生理鹽水稀釋,直到稀釋到OD600=0.5,兩個菌大小形態相似,所以對應的OD600也幾乎一樣,以此為比例進行組合,在所有的組合中有7組結果相對較好,如圖2所示。所有乳酸菌和丙酸桿菌菌株組合的測定結果如表4所示。
從圖2和表4中可以看出,乳酸菌中植物乳桿菌L9、3014、3035在與丙酸桿菌中D1和D5組合培養后表現出抗真菌活性的增加,表現出了抗真菌活性的協同作用。而植物乳桿菌L9、3014、3035在第一個結果中已被篩選出為單一抗真菌較強菌株,又與丙酸桿菌進行組合也被篩選出為抗真菌性較強組合,證明猜想正確。

圖 2 乳酸菌和丙酸桿菌組合對黃曲霉Y03的抑制效果Fig.2 Inhibitory effect of Lactobacillus and Propionibacterium combination on Aspergillus flavus Y03

表4 乳酸菌和丙酸桿菌的組合對黃曲霉Y03的抑制效果Table 4 Inhibitory effect of Lactobacillus and Propionibacterium combination on Aspergillus flavus Y03

圖 3 FIC法測定抗真菌活性Fig.3 Antifungal activity assayed by FIC method
2.3 FIC法探究復合菌抗真菌組合的協同作用
從上個結果中發現植物乳桿菌L9、3014和3035菌株與費氏丙酸桿菌D1和D5有較強的抗真菌活性,于是在本實驗中用FIC方法對這3×2種組合的協同抗真菌活性進行了進一步的探究,其結果如圖3所示。
從圖3中可以發現,在植物乳桿菌L9與費氏丙酸桿菌D1和D5的組合中,其抗真菌活性與兩種菌的濃度表現出正相關關系。而在植物乳桿菌3014和3035與費氏丙酸桿菌D1的組合中,10-2濃度的植物乳桿菌有著最佳的抗真菌效果,而過高的植物乳桿菌細胞濃度減弱了其對黃曲霉Y03的抑制作用,這可能是費氏丙酸桿菌D1與植物乳桿菌之間在過高濃度有相互抗衡作用, 但在費氏丙酸桿菌D1的抗真菌效果上看,最高的細胞濃度取得了最佳的抗真菌效果。費氏丙酸桿菌D5與植物乳桿菌3014和3035的組合的抗真菌效果并不理想,所有細胞濃度的抗真菌效果大致相當。
因此,在對比這3×2種植物乳桿菌和費氏丙酸桿菌混合培養所展現的協同抗真菌作用后,最終確定植物乳桿菌L9和費氏丙酸桿菌D5作為后續混合發酵中的抗真菌菌株組合。
2.4 Plackett-Burman試驗結果
本文通過用Plackett-Burmen試驗設計從12個考察因素中快速、有效地篩選出顯著因子,各因素的編碼和水平見表1,試驗設計及結果見表5,表中的1~23分別代表23組試驗,按照試驗設計進行試驗,每組三次平行樣[27]。
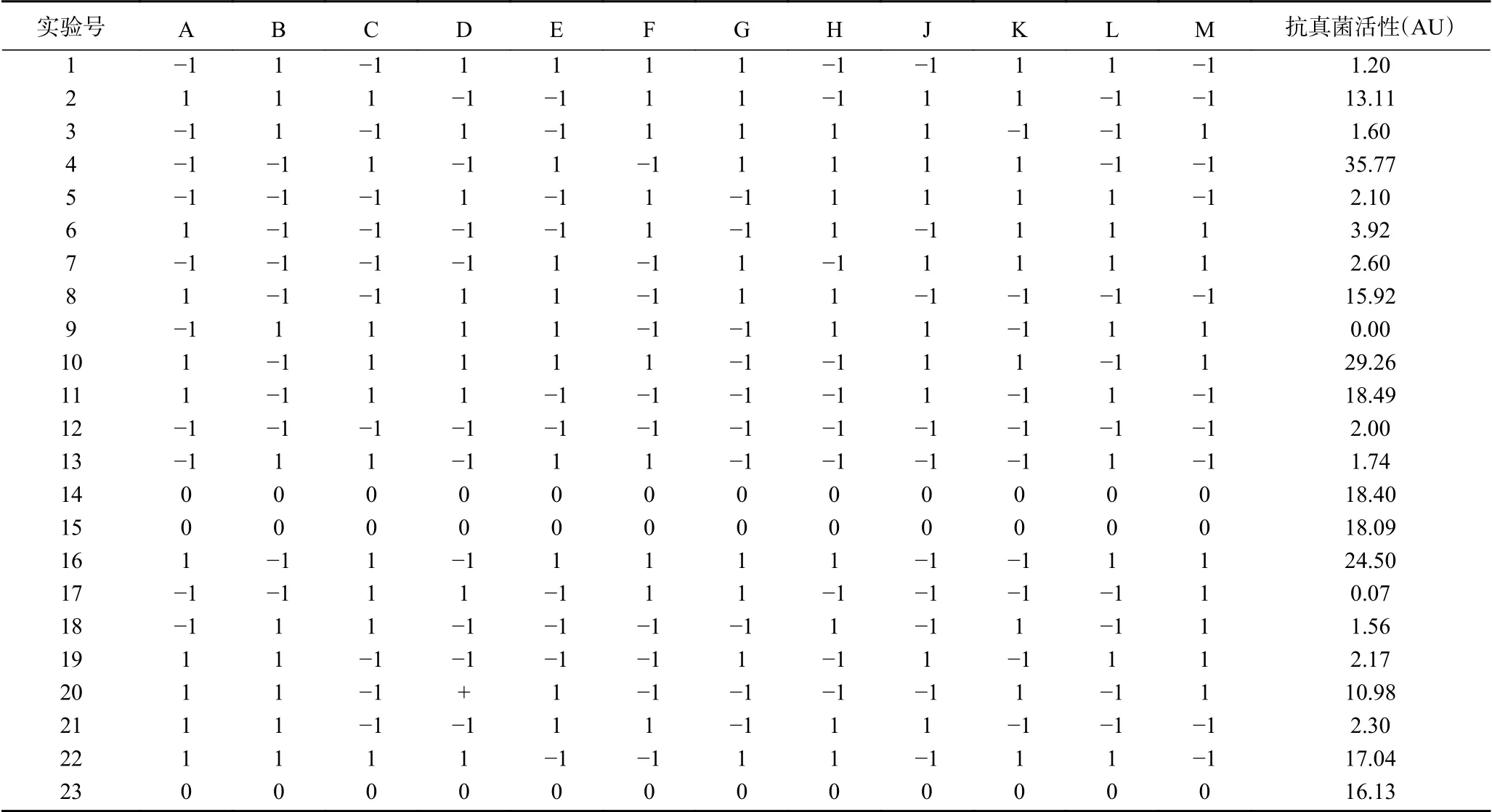
表5 Plackett-Burman試驗設計和結果Table 5 Plackett-Burman experimental design and results
Plackett-Burman試驗的方差分析表如表6所示,從表6中可以看出模型P值為0.03<0.05,表明多元回歸模型對結果的影響顯著。葡萄糖在模型中P值為0.014<0.05、碳酸鈣在模型中P值為0.02<0.05,酵母浸粉的P值為0.009<0.01,表示這三個因素的水平變化對植物乳桿菌L9和費氏丙酸桿菌D5菌株發酵液抗真菌活性有顯著的影響,其他因素不顯著。

表6 Plackett-Burman試驗的方差分析Table 6 Analysis of variance in Plackett-Burman design
12個因素的帕累托圖如圖4所示,12個因素的標準化效應值按照大小依次從上到下排列,當標準化效應值大于2.262時即為顯著[28]。從圖4中可以看出,標準化效應值大于2.26的三個因素依次分別為酵母浸粉(C)、葡萄糖(A)和碳酸鈣(B),表明這三個因素對植物乳桿菌L9和費氏丙酸桿菌D5菌株發酵液抗真菌活性的影響最大。對標準化效應值最小的酪蛋白胨、氯化鈉和硫酸鎂3個存在與否對發酵液抗真菌活性影響較小的培養基成分進行舍棄,最終得到的用于進行下一步爬坡試驗的發酵培養基配方為:葡萄糖40 g/L,碳酸鈣16 g/L,酵母浸粉 10 g/L,磷酸氫二鉀 0.25 g/L,硫酸錳 0.1 g/L,乙酸鈉 5 g/L,檸檬酸銨 2 g/L。接種比例為5:1(D5:L9)、發酵溫度為37 ℃。
2.5 最陡爬坡試驗結果
如表2和圖5所示最陡爬坡試驗的6個梯度條件及其發酵液的抗真菌活性結果。從中可以看出,隨著葡萄糖和酵母浸粉含量的增加,碳酸鈣含量的減少,發酵液抗真菌活性呈現先增后減趨勢,在編號為4的實驗組條件下(葡萄糖55 g/L、碳酸鈣6.7 g/L和酵母浸粉14.8 g/L)出現最大值,其發酵液抗真菌活性達到最大,為47.07 AU。因此,得到優化后的三個關鍵影響因素的最佳比例為葡萄糖55 g/L、碳酸鈣6.7 g/L和酵母浸粉14.8 g/L。
3 結論
本文以導致面包發霉的典型真菌(黑曲霉、黃曲霉、桔青霉、枝孢霉、交鏈孢霉)為指示菌[29],篩選出了5株對面包中污染霉菌出現頻率最高的兩種霉菌桔青霉和黃曲霉抑制效果比較明顯的單菌菌株,分別是編號為L8、L9的乳酸乳球菌,編號為3103、3014、3035的植物乳桿菌,并篩選出一組發酵液具有明顯抗真菌作用的復合菌株組合植物乳桿菌 L9 和費氏丙酸桿菌 D5,對其發酵工藝進行優化,選出對最終抗真菌活性影響最大的三個因素依次分別為酵母浸粉、葡萄糖和碳酸鈣,發酵培養基優化配方為葡萄糖55 g/L,碳酸鈣6.7 g/L,酵母浸粉14.8 g/L,磷酸氫二鉀 0.25 g/L,硫酸錳 0.1 g/L,乙酸鈉 5.0 g/L,檸檬酸銨 2.0 g/L,接種比例為5:1(D5:L9),發酵溫度為37 ℃,抗真菌活性可達47.07 AU。本論文首次對乳酸菌和丙酸桿菌協同抗真菌效果進行研究,結果表明復合菌發酵液抗真菌效果明顯,可在食品制作中加入復合菌發酵液替代部分水,抑制食品中腐敗霉菌的生長,延長食品保質期。
