蘋果樹腐爛病拮抗菌株的分離、篩選和鑒定*
2022-06-09 01:08:52宿靜瑤付崇毅杜美娥李正男孫平平
中國果樹 2022年5期
劉 欣,馬 強,宿靜瑤,付崇毅,杜美娥,李正男,孫平平
(1 內蒙古農業大學園藝與植物保護學院,呼和浩特010018)(2 內蒙古農牧業科學院蔬菜研究所)(3 鄂爾多斯生態環境職業學院)
蘋果樹腐爛病(Apple valsa canker)是由殼囊孢屬真菌(Cytosporasp.)侵染蘋果樹后引起的枝干病害。病害發病初期呈紅褐色,水漬狀,手壓發病部位產生凹陷,流出黃褐色帶酒糟味汁液,發病后期病部失水并干縮下陷,在發病部位散生黑色小粒點[1]。目前,蘋果樹腐爛病在我國所有蘋果種植區域均有發生,具有分布廣、危害重、防治難的特性,被果農稱為蘋果樹的“癌癥”[2]。
目前,對蘋果樹腐爛病的防治方法中采用最多、效果最顯著的就是刮除法,即刮除病斑及周圍無癥狀組織中潛伏侵染的病原菌,配合戊唑醇、烯唑醇、菌毒清、瑞拉菌素等藥劑涂抹或者噴施患病部位[3-4],雖然該方法對蘋果樹腐爛病的防治起到了一定的作用,但是,刮除法費時費力,而且化學藥劑具有易產生抗藥性和環境污染等缺點。因此,尋找持效、低毒、環境友好型的防治方法成為蘋果樹腐爛病防治的發展趨勢。生物防治具有持效性和環境友好型的優點,目前已經成為植物病害防治的熱點。在蘋果樹腐爛病的生物防治中,已經有大量研究者開展了相關工作,如翟世玉[5]、鄧振山等[6]、Li等[7]和Zhang 等[8]分別證明枯草芽孢桿菌LF17、銀杏內生真菌、楊凌糖絲菌Hhs.015 和解淀粉芽孢桿菌GB1 對蘋果樹腐爛病具有較好的防效。
我國幅員遼闊,土壤類型多樣,微生物資源豐富。在先前報道獲得的優質生防菌資源中,很大一部分來自于秦嶺地區及新疆、西藏、青海等省(自治區)少被污染或極限環境下[9-14]。內蒙古草原面積廣闊,這些草原土壤從未被開墾過,因此其土壤中一定存在大量抗逆性強,且生防效果良好的生防菌資源。本研究從內蒙古達茂草原中采集根際土壤,以蘋果樹腐爛病病原菌Cytospora maliQH2 為供試病原菌,利用皿內拮抗結合枝條測定方法對分離的菌株進行篩選,并對其抑菌譜、耐鹽、耐紫外等抗逆特性進行測定,以期為蘋果樹腐爛病的有效防控提供更多的資源。
1 材料與方法
1.1 材料
1.1.1 供試病原菌
蘋果樹腐爛病菌(C.mali)QH2 由馬強等分離自內蒙古呼和浩特市謙和果園感染了腐爛病的金紅樹[15];梨輪紋病菌(Botryosphaeria berengerianade)、梨黑斑病菌(Alternaria alternata)、蒂腐病菌(Botryodiplodia theobromae)、梨炭疽病菌(Glomerella cingulate)、梨褐腐病菌(Monilinia fructigena)由內蒙古農業大學園藝病害病原生物學及綜合防控研究室保存;變紅鐮刀病菌(Fusariumincarnatum)、尖孢鐮刀病菌(Fusarium oxysporum)、茄鐮刀病菌(Fusarium solani)由寧夏大學顧沛文教授提供;辣椒疫霉病菌(Phytophthora capsici)由西南大學樊光進博士提供。
1.1.2 儀器設備
離心機(HICO21)、恒溫智能光照培養箱(HPX-9162MBE,上海博訊實業有限公司醫療設備廠)、恒溫振蕩器(THZ-82)、PCR 儀(T100,Bio-Rad Laboratories)、紫外分光光度計(UV-2800,北京普析通用儀器有限責任公司)、顯微鏡(LEICA ICC50W,德國萊卡儀器有限公司)等。
1.1.3 培養基
馬鈴薯葡萄糖培養基(PDA):200 g 馬鈴薯,20 g 葡萄糖,20 g 瓊脂粉,1 000 mL 蒸餾水,pH 值7.2~7.4。
改良高氏1 號培養基:20 g 可溶性淀粉,0.5 g NaCl,1 g KNO3,0.5 g K2HPO4,0.5 g MgSO4·7H2O,0.01 g FeSO4,20 g 瓊脂粉,1 000 mL 蒸餾水,pH值 7.2~7.4,培養基滅菌后,待溫度降到50~55 ℃,加入3.3 mL 的3%重鉻酸鉀。
LB 培養基:10 g 胰蛋白胨,5 g 酵母提取物,10 g NaCl,15 g 瓊脂粉,1 000 mL 去離子水。
小米培養基:10 g 小米加蒸餾水煮20 min,過濾后的上清液用蒸餾水定容至1 000 mL,加入1 g葡萄糖、0.3 g 蛋白胨、0.25 g NaCl、0.02 g CaCO3。
V-P 反應:5 g 葡萄糖,2 g K2HPO4,5 g 蛋白胨,1 000 mL 去離子水。
硝酸鹽還原培養基:5 g 蛋白胨,1 g KNO3,1 000 mL 去離子水。
明膠液化培養基:5 g 蛋白胨,3 g 牛肉浸粉,120 g 明膠,2 g KNO3,1 000 mL 蒸餾水。
淀粉水解培養基:10 g 蛋白胨,3 g 牛肉浸粉,30 g 可溶性淀粉,15 g 瓊脂粉,1 000 mL 去離子水,pH 值7.6±0.1。
H2S 培養基:3 g 牛肉膏,3 g 酵母浸膏,10 g多胨,0.2 g FeSO4,5 g NaCl,0.3 g 硫代硫酸鈉,12 g 瓊脂,1 000 mL 蒸餾水。
1.2 土壤樣品的采集、菌株的分離與純化
2019 年9月從內蒙古包頭市達茂草原上采集土壤樣品,采集時去掉表層5 cm 的表土,采集25 cm深的根際土壤,用牛皮紙袋包好后運至實驗室4 ℃保存。采用稀釋平板涂布法[16]分離土壤中細菌菌株。稱10 g 干燥處理的土壤樣品研磨,放入100 mL 無菌水中,4 ℃180 r/min 振蕩30 min,利用無菌水將土樣逐級稀釋成100、1 000 倍,吸取100 μL 的稀釋液均勻涂布于LB(細菌)與改良高氏1 號(放線菌)培養基上,28 ℃培養2 d。挑取形態、顏色不同的單個細菌菌落劃線純化,純化后的菌株轉入20%甘油中-20 ℃保存備用。
1.3 活性菌株的皿內篩選
采用平板對峙法[17]對分離得到的細菌和放線菌進行活性篩選。細菌拮抗方法:用直徑6 mm 的無菌打孔器在PDA 平板的兩側打孔,挑去瓊脂塊后,將在28 ℃培養2 d 的細菌菌餅(5 mm 直徑)倒置放入孔中,在PDA 平板中央接種28 ℃培養5 d 的C.maliQH2,28 ℃恒溫培養。放線菌的平板對峙法:在PDA 平板兩側接種直徑6 mm 的放線菌菌餅,28 ℃恒溫培養2 d 后在平板中央接種C.maliQH2,以只接種C.maliQH2 作平板對照,28 ℃恒溫培養。待對照菌落長滿平板時,觀察并記錄抑菌圈直徑,計算抑菌率。
抑菌率(%)=(1-拮抗處理病斑半徑/對照病斑半徑)×100
1.4 活性菌株的離體枝條拮抗活性測定
參照翟世玉[5]的方法測定活性菌株對蘋果樹腐爛病在金紅和黃太平枝條上的拮抗活性。挑取篩選的菌株接種到50 mL LB 液體培養基(細菌)或小米液體培養基(放線菌),28 ℃180 r/min 培養48 h用于離體枝條拮抗活性測定。選2 年生健康蘋果枝條,截成10 cm 左右,于2%次氯酸鈉中浸泡5 min,75%酒精浸泡3 min,無菌水沖洗3 次后晾干,石蠟封住枝條兩端,利用無菌打孔器在蘋果枝條中間部位致6 mm 的傷口。將拮抗菌菌液均勻噴施在枝條上,0.5 h 后將C.maliQH2 貼在傷口上,用無菌水浸濕的無菌脫脂棉包裹,保鮮膜封口。以致傷后噴施LB 液體培養基再接種腐爛病菌菌餅的枝條為陽性對照,不接種蘋果樹腐爛病菌菌餅的枝條為陰性對照,所有枝條25 ℃16 h 光照/8 h 黑暗保濕培養。接菌5 d 后揭掉棉花,14 d 后測量病斑直徑,計算抑菌率。
抑菌率(%)=(1-處理平均值/CK 平均值)×100
1.5 活性菌株對蘋果樹腐爛病菌菌絲生長的影響
挑選出對蘋果樹腐爛病菌皿內和離體枝條上均具有顯著拮抗活性的菌株,參照1.3 平板對峙方法接種生防菌株和病原菌,待對照長滿皿后挑取抑菌圈邊緣處菌絲并在光學顯微鏡下觀察菌絲形態,以不接生防菌的蘋果樹腐爛病菌平板為對照,比較拮抗菌株對蘋果樹腐爛病菌菌絲形態的影響。
1.6 活性菌株的抑菌譜測定
將供試的9種病原菌轉入PDA 培養基中,28 ℃培養5 d。參照1.3 方法,利用平板對峙法[17]測定篩選活性菌株對9 種病原菌的抑菌活性。
1.7 穩定性測定
耐鹽性:將不同量的NaCl 加入到LB 培養液中[18],配制成含0.1%、0.5%、1%、2%、5%、10%、12%、15%NaCl(W/V)的LB 培養液,接種1 mL活性菌株的種子液,28 ℃180 r/min 培養48 h,測定OD600。
紫外線穩定性:用0.22 μm 的細菌濾膜將28 ℃180 r/min 培養48 h 的活性菌發酵液過濾,得到無菌發酵液,置于30 W 紫外燈下[18],垂直距離45.5 cm 處,照射10、20、30、40、50、60、90、120 min。采用牛津杯法[19]檢測細菌活性,置于28 ℃恒溫培養,每個處理重復3 次,以不經紫外線照射的無菌發酵液為對照,觀察并記錄抑菌圈直徑,測定紫外線對其活性的影響。
1.8 活性菌株鑒定
1.8.1 活性菌株的形態學特征觀察
將拮抗菌株在LB 固體培養基上劃線培養,28 ℃培養24 h,觀察記錄菌落形狀、透明度及菌落顏色等菌落特征。
1.8.2 分子鑒定
將拮抗菌株接種于LB 液體培養基,28 ℃180 r/min 振蕩培養24 h,取1 mL 菌液10 000 r/min 離心1 min,使用Ezup 柱式細菌基因組DNA 抽提試劑盒對拮抗細菌的DNA 進行提取[20]。使用通用引物7F/1492R(5′-AGAGTTTGATCCTGGCTCAG-3′/5′-GGT TACCTTGTTACGACTT-3′),對拮抗菌的16 S rRNA序列進行PCR 擴增,反應體系如下:2×M5 Hipper超光速Mix 10 μL,ddH2O 7 μL,DNA 模板1 μL,上下游引物各1 μL;反應條件:94 ℃5 min;94 ℃45 s,55 ℃45 s,72 ℃90 s,30 個循環;72 ℃10 min。反應結束后用1%瓊脂糖凝膠電泳檢測,并用Sanprep 柱式DNA 膠回收試劑盒(SK8131)回收目的條帶,連接到pMD19-T 載體,轉化到Escherichia coliDH5α 感受態細胞,涂布于含Amp 的LB 平板上,37 ℃培養過夜,挑選單菌落,將PCR 擴增鑒定陽性的單克隆送交上海生工測序。使用Vector NTI Advance 11 軟件將測序結果進行校正和拼接,用BLASTn 對拼接的序列進行同源序列檢索,確定其分類地位,應用MEGA7.0 軟件,采用Clustal W算法對序列進行比對,用比鄰法(NJ)構建系統發育進化樹。
1.8.3 生理生化特征的鑒定
參照《常見細菌系統鑒定手冊》[21]的方法測定生理生化特征反應,包括對V-P 反應、硝酸鹽還原反應、明膠液化反應、淀粉水解、H2S 的利用能力。
V-P 反應:將菌株接種在V-P 液體培養基中,以空白培養基為對照,28 ℃下培養7~14 d。將少許培養7、14 d 的培養液加入試管中,加入6 滴A液(α-萘酚6 g,無水乙醇溶解,定容至100 mL)與培養物充分混勻,再加入2 滴B 液(氫氧化鉀40 g,蒸餾水溶解,定容至100 mL)混勻。振搖試管后靜置0.5~2 h 后,如變紅則為陽性,不變色為陰性。
硝酸鹽還原:將菌株接種在硝酸鹽還原液體培養基中,以空白培養基為對照,28 ℃下培養7~14 d。將少許培養7~14 d 的培養液加入試管中,加入2 滴A 液(對氨基苯磺酸8.0 g,溶解于1 000 mL 的5 mol/L 乙酸中)和2 滴B 液(α-萘酚2.5 g,溶解于1 000 mL 的5 mol/L 乙酸中)混勻。如變紅則為陽性,不變色為陰性。
明膠液化:將菌株接種到明膠液化培養基上,空白培養基為對照,28 ℃恒溫培養7 d,觀察前先將試管放在低溫環境中冷卻20~30 min,待對照培養基凝固后,觀察接種細菌的試管是否液化以及液化的程度。
淀粉水解:將待測菌株以點接種在淀粉水解瓊脂培養基上,直徑<5 mm,28 ℃恒溫培養7 d。加碘液到沒有生長菌落的培養基上,若碘液沒有使培養基變藍,而是形成了透明圈,表示菌株產生了淀粉酶,透明圈越大,淀粉酶活性越強;若培養基遇碘變藍,表示菌株沒有產生淀粉酶。
H2S:將菌株點接在H2S 培養基上,以空白培養基為對照,28 ℃下培養7 d,觀察培養基菌落周圍的顏色。若培養基菌落周圍變成黑色,說明有H2S 產生。若培養基不變色,則無H2S 產生。
2 結果與分析
2.1 拮抗菌的分離和皿內篩選
從6 份土壤樣品中分離純化得到細菌288 株,放線菌54 株。經皿內拮抗篩選得到具有拮抗作用的細菌18 株,放線菌4 株(表1)。在18 株細菌中,有7 株細菌的抑菌圈直徑在1 cm 以上,其中菌株5-29 的抑菌帶寬度最大,達到1.35±0.19 cm,抑菌率為45%;在4 株放線菌中,3 株放線菌5-2、2-1和4-8 對C.maliQH2 具有明顯的抑制活性,抑菌帶寬度均在0.70 cm 以上。

表1 拮抗菌平板對峙法篩選結果
2.2 活性菌株的離體枝條拮抗活性
用燙傷接種法測定皿內篩選出的18株細菌和4株放線菌先后在金紅和黃太平枝條上對蘋果樹腐爛病的拮抗活性。結果顯示,接種細菌5-29、3-18、1-28 與3-10 的金紅蘋果枝條上的病斑長度分別為2.12±2.09 cm、2.26±1.73 cm、2.39±2.68 cm 和2.69±2.22 cm,顯著低于對照,抑菌率均在57%以上,有良好的抑菌效果。其余細菌的抑菌率均在50%以下,效果相對較差。接種待測4 株放線菌中,5-2的金紅蘋果枝條上的病斑長度為2.63±1.96 cm,對蘋果樹腐爛病的抑制率為58.91%,其余3 株病斑長度均大于3 cm,抑制率均在55%以下。后續利用黃太平繼續測定篩選出的4 株細菌與放線菌5-2 對蘋果樹腐爛病的防治效果,結果顯示4 株細菌在黃太平枝條上對C.maliQH2 的抑制率在60%以上,病斑長度顯著低于對照,其中菌株5-29 對蘋果樹腐爛病的抑制率達到77.13%,菌株3-18 對蘋果樹腐爛病的抑制率高達81.98%,放線菌5-2 的抑菌率只有40.89%,抑菌效果較弱(表2)。故而,選擇4株細菌進行后續的穩定性測驗。
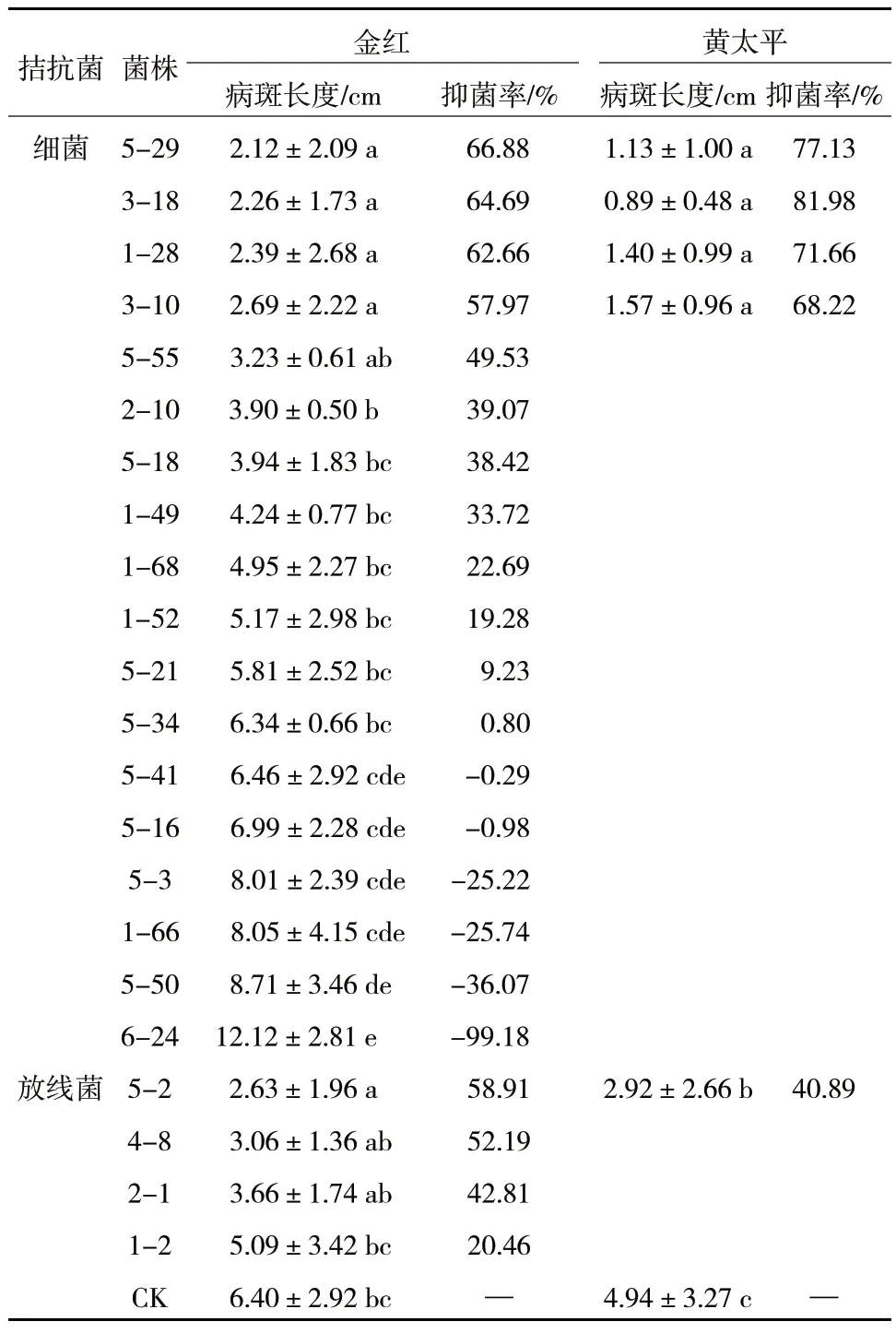
表2 拮抗菌離體枝條檢測結果
2.3 拮抗菌對病原菌菌絲的影響
蘋果樹腐爛病菌菌株與拮抗菌對峙培養3~5 d后,病原菌菌落前沿的顏色變為深褐色,菌絲顏色明顯加深,挑取菌落邊緣深褐色菌絲在顯微鏡下觀察,發現菌絲與正常菌絲相比頂端顯著膨大,明顯畸形,菌絲粗細不均勻,出現菌絲的分支增多或細胞質外滲的現象,對菌絲的生長有一定的影響。
2.4 拮抗菌的抑菌譜
利用皿內拮抗方法測定4 株活性菌株對9 種常見病原菌的抑菌效果,結果顯示,菌株5-29、3-18和3-10 具有廣泛的抑菌譜,對供試的9 種病原菌均具有一定的拮抗效果,而菌株1-28 對變紅鐮刀、尖孢鐮刀、茄鐮刀和辣椒疫霉4 種病原菌無顯著的抑制活性(表3)。
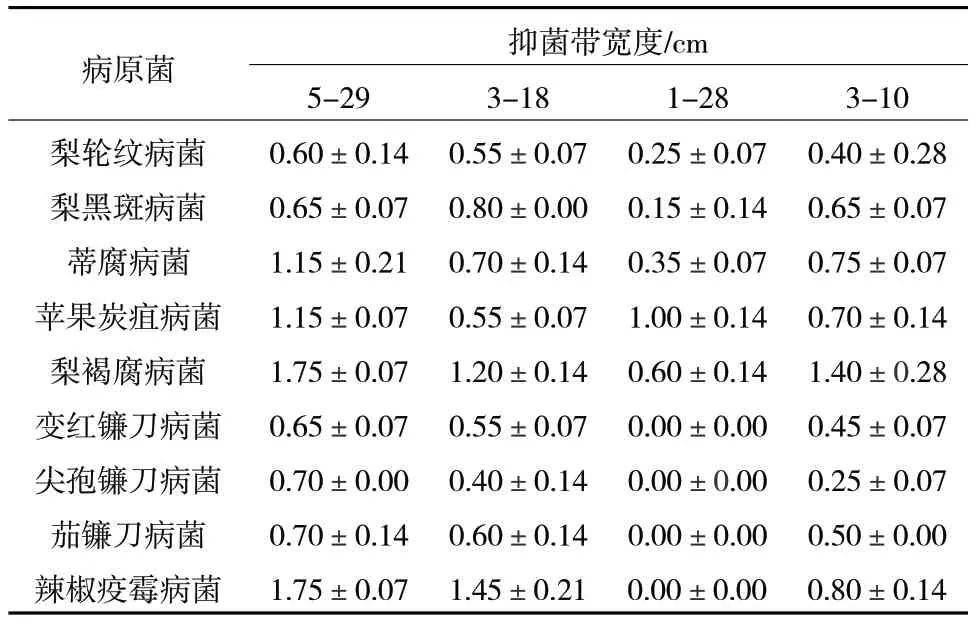
表3 拮抗菌對9 種常見病原菌的抑菌效果
2.5 菌株穩定性測定
2.5.1 拮抗菌的耐鹽性
4 株活性菌株在不同NaCl 濃度下生長狀況測定結果顯示,當NaCl 濃度≤2%時,4 株菌株的OD600值均在2.0 以上,當NaCl 濃度達到5%時,OD600值1.4~1.9,而當濃度達到10%時,除菌株3-18 外,其余3 株菌株5-29、1-28、3-10 均不能生長,當NaCl 濃度達15%時,4 株菌株均不生長,因此菌株3-18 耐鹽性最強,可耐12%的NaCl,其他3 株細菌可耐5%的NaCl(表4)。
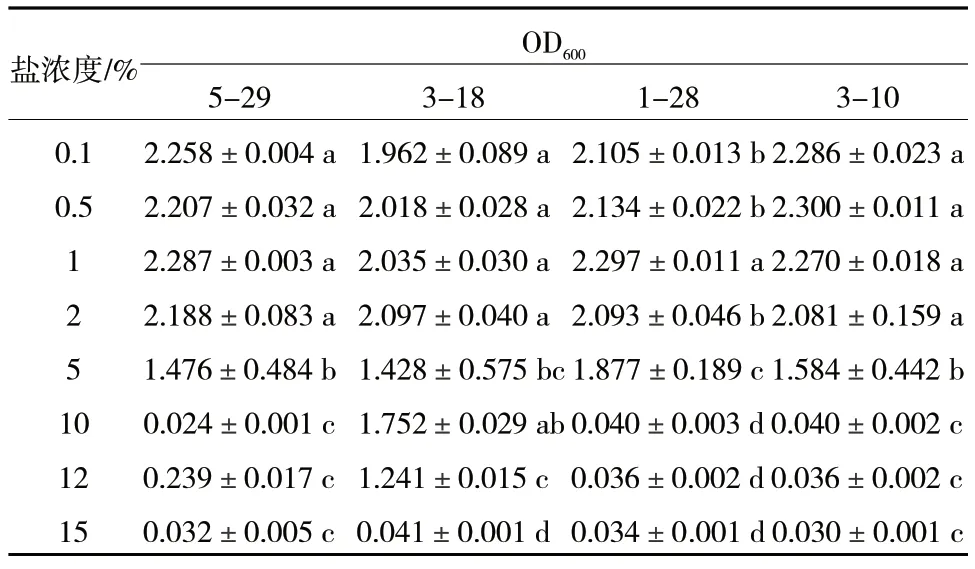
表4 菌株耐鹽性測定
2.5.2 紫外線穩定性
將4 株活性菌株無孢子發酵濾液經紫外線照射不同時間后測定其對C.maliQH2 的拮抗活性。結果見表5,紫外線照射越久,菌株對C.maliQH2 的拮抗性越弱,紫外線照射120 min 后,菌株5-29 和3-18對C.maliQH2 的抑菌圈直徑分別為0.63±0.08 cm和0.48±0.03 cm,具有拮抗作用,而菌株1-28 和3-10 均已失活。因此,菌株5-29 與3-18 具有較強的耐紫外活性。鑒于皿內試驗、離體防效、抑菌譜和穩定性試驗結果,我們對5-29 和3-18 進行鑒定。
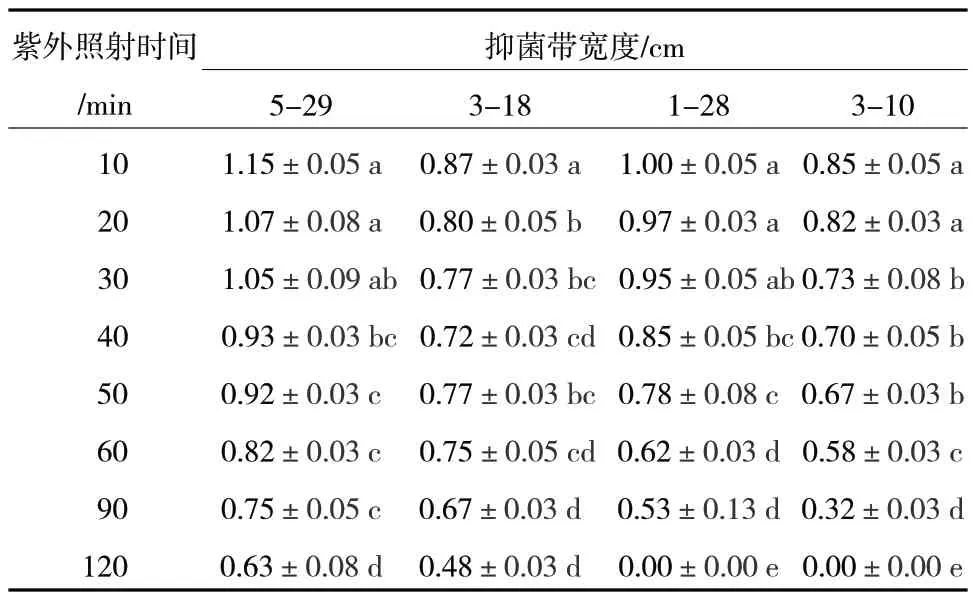
表5 4 株菌經紫外處理的穩定性結果
2.6 活性菌株的種類鑒定
2.6.1 形態學及分子鑒定
菌株5-29 與3-18 具有典型細菌的培養特征,在LB 平板上呈乳白色和灰白色,質地黏稠不易挑起,對其16 S RNA 序列進行擴增,分別得到1 427 bp(Genebank:MZ411444)與1 430 bp(Genebank:MZ411445)的擴增產物。將其序列與數據庫中的序列進行BLAST 分析,并構建進化樹,結果顯示,5-29與B.velezensisFZB42 菌株聚到一支,3-18 與B.atrophaeus聚到一支(圖1)。綜合形態學及基于16 S rRNA 的系統發育分析,確定分離到的菌株5-29為貝萊斯芽孢桿菌(B.velezensis),菌株3-18 鑒定為萎縮芽孢桿菌(B.atrophaeus)。

圖1 活性菌株DM5-29 與DM3-18 的系統發育進化樹
2.6.2 生理生化特性
菌株5-29 與3-18 的V-P 和硝酸鹽還原反應均呈陽性,有明膠液化現象,可以水解淀粉,不能產生H2S,符合貝萊斯芽孢桿菌與萎縮芽孢桿菌的生理生化特征[22-23],進一步確定了菌株5-29 為貝萊斯芽孢(B.velezensis),菌株3-18 鑒定為萎縮芽孢桿菌(B.atrophaeus)(表6)。
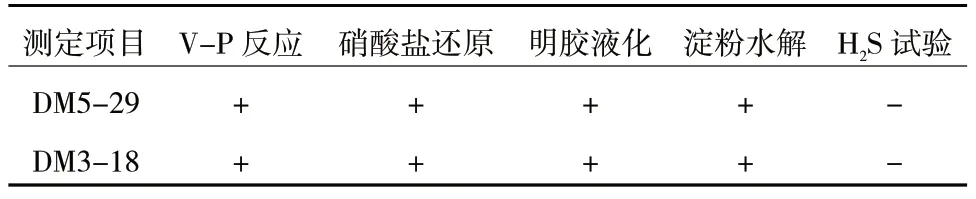
表6 生理生化特征
3 討論
本研究從采集自內蒙古自治區達茂旗從未被開墾過的6 份草原土壤樣品中獲得了細菌288 株、放線菌54 株,并且獲得的B.velezensisDM5-29 和B.atrophaeusDM3-18 對蘋果樹腐爛病防效顯著,對環境適應性強。說明草原的復雜環境賦予了菌株極強的適應能力,該研究也呼應了從其他極端環境下獲得的菌株具有好的生防效果并且適應性強的研究結果[9-14]。
貝萊斯芽孢桿菌(B.velezensis)在生物防治、藥物研發、食品發酵和工業應用等方面具有重要作用[22],近些年用于生物防治的研究在國內外的報道越來越多,研究主要集中在其促進植物生長、拮抗病原菌、誘導系統抗性和抑菌物質等方面。例如陳爽等[24]從土壤中分離得到的B.velezensisHM3-3,對盆栽大豆根腐病的防效可達46%。孫平平等[18]發現B.velezensisL-1 對梨灰霉病菌等保護和防控的活體抑制率分別為92.88%和77.47%,使病原菌的菌絲膨大、畸形,具有穩定的拮抗效果。萎縮芽孢桿菌(B.atrophaeus)可以在80 ℃以上的溫度生長,抗性強,無致病性,廣泛應用于醫療、工業、農業和衛生等領域[23]。馬榮等[25]發現,B.atrophaeusXW2發酵濾液對蘋果樹腐爛病菌菌絲生長抑制率為52.44%,孢子萌發抑制率為69.26%,離體防效試驗表明,B.atrophaeusXW2 發酵液對5 個不同品種蘋果樹腐爛病的預防效果在97.95%以上,治療效果在70.76%以上;王璐[26]發現,B.atrophaeusCP297-AE對桃黃曲霉的抑菌效果明顯,可以引起菌絲的嚴重變形。本研究從內蒙古達茂草原分離篩選出的B.velezensisDM5-29 和B.atrophaeusDM3-18 均能有效防治蘋果樹腐爛病,在金紅和黃太平離體枝條上的防效分別在64%和77%以上,同時DM5-29 及DM3-18 對梨輪紋病菌、梨黑斑病菌、蒂腐病菌、梨炭疽病菌、梨褐腐病菌、變紅鐮刀病菌、尖孢鐮刀病菌、茄鐮刀病菌、疫霉病菌9 種重要園藝作物病原菌均具有一定的拮抗效果,抑菌譜范圍廣,有較好的應用前景。該結果為貝萊斯芽孢桿菌和萎縮芽孢桿菌的功能以及蘋果樹腐爛病的生防菌資源提供了更多的信息。
本研究通過皿內篩選得到具有良好防效的細菌18 株、放線菌4 株,但在活體枝條測定中有14株細菌和4 株放線菌活性很低或失活。生防菌在應用中受到環境、病原菌以及寄主等多方面影響防效不穩定[27]。因此,利用皿內拮抗結合活體枝條以及田間試驗對病害進行防控是篩選菌株的必要措施。苑琳等[28]利用B.atrophaeusSF1 對尖孢鐮刀菌胡麻專化型進行抑菌研究發現,B.atrophaeusSF1 在不同地區之間的防效出現了差異,其中內蒙古室外的防效最差。內蒙古地區土壤多屬于鹽堿地,紫外照射強,篩選出耐鹽、耐紫外菌株是保證田間防效的關鍵。前期報道的B.velezensisL-1 在10%的NaCl 培養液中可正常生長,但在12%的NaCl 培養液中活性顯著下降[18],解淀粉芽孢桿菌GM-1 在紫外線照射40 min 抑菌率明顯下降[29]。本研究中的菌株3-18可以在12%的NaCl 培養液中生長,5-29 可耐5%的NaCl;紫外線照射2 h 內,菌株5-29 和3-18 的無菌發酵液對C.maliQH2 仍具有一定的拮抗活性。說明5-29 和3-18 不但生防效果顯著,而且具有很強的抗逆性,這為2 個菌株的施用奠定了基礎。
4 結論
本研究從內蒙古達茂草原土壤中分離細菌和放線菌,采用平板對峙和離體枝條法,結合耐鹽、耐紫外的穩定性篩選,最終獲得B.velezensis5-29和B.atrophaeus3-18,2 個株菌對蘋果樹腐爛病防效顯著、抗逆性強,有應用潛力,后續將對其田間應用進行研究。
