基于宏轉錄組學技術解析濃香型酒醅活性微生物群落結構及功能變化特征
2022-06-02 08:42:46胡曉龍田瑞杰李保坤何培新朱文優
食品科學 2022年10期
關鍵詞:功能
胡曉龍,田瑞杰,李保坤,張 勇,遲 雷,何培新,魏 濤,朱文優
(1.宜賓學院農林與食品工程學部,四川 宜賓 644000;2鄭州輕工業大學食品與生物工程學院,食品生產與安全河南省協同創新中心,河南 鄭州 450000)
濃香型白酒作為我國優勢傳統發酵食品的典型代表,其以窖香濃郁、香味協調、尾凈余長等為特征的風格深受消費者喜愛。其生產主要采用復雜且獨特的半開放、固態發酵模式,包括原輔料處理、續醅拌和、配入曲藥、多微共酵及精心勾調等上百道工序。濃香型白酒釀造微生物主要來源于源釀造環境、器具、發酵劑、原料及窖泥,主要包括細菌、酵母、霉菌及古細菌,其在窖池內長期進行的物質和能量交換、轉化是形成濃香型白酒呈香物質及獨特風格的主要驅動力。
酒醅作為微生物發酵的主要載體和白酒呈香物質的直接來源,其微生物群落多樣性及其演替性一直是研究的熱點。近年來,免培養技術的廣泛應用(變性梯度凝膠電泳、克隆文庫、二代測序等),使得濃香型酒醅微生物群落多樣性、結構及其演替規律逐漸清晰。例如,在酒醅中觀察到的細菌和真菌屬數量分別高達496和155。其中,、、、和等是優勢細菌屬(相對豐度≥1%);、、和等是優勢真菌屬。整體上,發酵前期(前7 d左右)酒醅中細菌群落多樣性較高,隨后細菌群落多樣性急劇下降,且逐漸成為酒醅中絕對優勢屬。
盡管以上通過16S rRNA或ITS基因測序(DNA水平)方法得到的研究結果極大地豐富了人們對酒醅微生物群落物種多樣性(多樣性)及組成動態變化(多樣性)的認知,但基于DNA的方法可以檢測到死亡或休眠狀態細胞的DNA,卻不能準確反映濃香型白酒發酵過程中微生物的生理或轉錄活性(簡稱活性)及其功能。目前,基于RNA水平的宏轉錄組測序技術能夠準確揭示目標環境樣品中活性微生物群落結構。諸多研究表明,分別采用基于DNA和RNA方法分析的環境微生物群落結果存在較大差異。例如,中國臺灣烏龍茶中常見的細菌在DNA水平上為Bacteroidaceae(21.7%)、Microbacteriaceae(22%)和Fusobacteriaceae(12.3%),而在RNA水平上為Bacillaceae(42%)和Enterobacteriaceae(36%)。Duan Shan等利用宏轉錄組測序在蝦醬細菌群落中檢測到了、、和,而利用16S rRNA基因測序未檢測到。同時,宏轉錄組測序技術能更為準確地揭示環境微生物功能及代謝途徑,如醬香型白酒發酵過程中涉及乙醇和乳酸合成的主要途徑分別為糖酵解和丙酮酸代謝。
目前關于濃香型白酒酒醅活性微生物群落及其功能變化的研究鮮有報道。因此,本研究以濃香型白酒固態發酵主發酵期不同發酵節點(3、7 d和25 d)的酒醅樣品為研究對象,運用宏轉錄組學技術對濃香型白酒固態發酵過程中活性微生物群落結構進行解析并確定其優勢活性微生物組成,同時對發酵過程中功能基因的差異表達情況進行探究,以期為對濃香型白酒固態發酵機理解析及發酵過程控制提供理論基礎。
1 材料與方法
1.1 材料與試劑
白酒發酵過程中的酒醅樣品來自河南省某知名白酒企業(E111°33’~112°01’,N34°36’~35°05’)。
十二烷基硫酸鈉(sodium dodecyl sulfate,SDS) 上海百賽生物技術股份有限公司;-巰基乙醇、異戊醇、聚乙二醇、焦碳酸二乙酯 上海麥克林生化科技有限公司;水飽和酚(pH 4.5) 上海源葉生物科技有限公司;無水乙醇 天津市富宇精細化工有限公司;Tris、聚乙烯吡咯烷酮、RNase-Free Water、2×Master Mix、2000 Marker 北京索萊寶科技有限公司;氯化鎂、氯化鈉、氯化鈣、草酸鈉、磷酸二氫鈉、磷酸氫二鈉、EDTANa北京北化精細化學品有限責任公司;Ribo-ZeroMagnetic Gold Kit(Yeast)試劑盒 北京中北林格科技發展有限公司;分離用有機溶劑均為國產分析純。
1.2 儀器與設備
MP200A精密電子天平 上海良豐儀器儀表有限公司;TGL-20M高速離心機 上海盧湘儀離心機儀器有限公司;DYY-8C電泳儀 北京市六一儀器廠;Mixer4K微型渦旋混合儀 生工生物工程(上海)股份有限公司;85-2恒溫磁力攪拌器 江蘇金壇市中大儀器廠;IMS-40全自動雪花制冰機 常熟市雪科電器有限公司;LDZX-50KBS立式壓力蒸汽滅菌器 上海中安醫療器械廠;SW-CJ-1超凈工作臺 蘇州凈化設備有限公司;WD-9403C紫外分析儀 北京六一生物科技有限公司;NanoDrop2000微量分光光度計 賽默飛世爾科技公司;2100生物分析儀 北京愛立斯生物科技有限公司。
1.3 方法
1.3.1 樣品采集
分別對發酵第3、7、25天的窖池進行酒醅樣品采集,每個發酵節點隨機選取3 個平行窖池,對每個窖池中層位置的酒醅進行取樣并混合作為該發酵節點的代表樣,將代表樣放入無菌袋中并置于盛有干冰的泡沫箱中,然后立即帶回實驗室進行總RNA提取。
1.3.2 酒醅總RNA的提取
按照胡曉龍等SDS-苯酚法對酒醅進行總RNA提取,然后采用1%瓊脂糖凝膠電泳、NanoDrop2000微量分光光度計和2100生物分析儀評估總RNA的純度和完整性,最后選擇高質量的RNA樣本構建Illumina測序文庫。
1.3.3 宏轉錄組測序文庫準備和Illumina測序
RNA樣本檢測合格后,使用Ribo-ZerorRNA Removal Kits(Bacteria)和Ribo-ZeroMagnetic Gold Kit(Yeast)試劑盒分別去除細菌和真菌核糖體RNA。文庫的構建和測序在北京奧維森基因科技有限公司Illumina HiSeq 2500(Illumina,San Diego,CA,United States)平臺上完成。
1.4 數據分析
測序下機獲得原始數據,通過過濾帶有測序接頭、未知堿基含量大于1%和低質量堿基(≤20)含量大于50%的reads得到clean reads,利用diamond將其進行NR數據庫比對分析,并利用megan進行微生物分類學解析。以酒醅活性細菌屬和真菌屬水平數據為基礎計算酒醅中活性微生物群落的Shannon指數,以評估酒醅活性微生物物種多樣性。采用Trinity(v0.27)對clean reads進行拼接,然后使用prodigal軟件對拼接組裝得到的contig序列進行開放閱讀框預測,再通過CD-HIT軟件對預測的基因序列以0.95相似度去除冗余基因,從而得到非冗余基因集。通過FPKM(每百萬片段中來自某一基因每千堿基長度的片段數目)值對所有基因進行表達豐度水平計算,再使用統計軟件R中edgeR包和DESeq2包,分析不同樣本間表達豐度具有差異的基因(differentially expressed gene,DEG),并使用火山圖對DEG的整體分布情況進行分析,閾值設定為|logfold change|>1且value<0.05。最后對DEG進行基因本體論(Gene Ontology,GO)富集分析,通過把所有DEG向GO數據庫(http://www.geneontology.org/)的各個term映射,計算每個term的基因數目,然后找出與整個基因組背景相比,在DEG中顯著富集的term,且1 個DEG可以分布在多個條目中。選取每個比較組中對應Corrected_value值由小到大排序的前30 個功能條目進行繪柱狀圖,其中Corrected_value<0.05說明這些功能條目在DEG中顯著富集。
2 結果與分析
2.1 序列預處理結果
由表1可知,經高通量雙端測序及序列質量控制,每個樣品獲得的序列的和堿基分別高于97%和92%,表明序列數據可靠,可以用于下一步分析。不同發酵時間節點酒醅宏轉錄組序列中GC含量差異較大(41%~48%),表明不同發酵時間酒醅中活性生物群落組成或轉錄的基因序列組成存在較大的差異。

表1 樣品宏轉錄測序信息Table 1 Metatranscriptomic sequencing information of each sample
2.2 酒醅中活性物種注釋


圖1 濃香型酒醅中不同分類水平的優勢生物(含微生物)群落組成Fig.1 Composition of dominant microbial communities at different taxonomic levels in strong-flavor baijiu fermented grains
2.3 活性微生物群落α多樣性及β多樣性
在長期發酵過程中,酒醅微生物群落不斷演替,期間伴隨著物質和能量代謝,因此從RNA水平揭示不同發酵節點酒醅活性微生物群落多樣性(組內多樣性)及多樣性(組間多樣性)將更有助于進一步揭示酒醅微生物的功能、代謝途徑及濃香型白酒發酵機制。
2.3.1 酒醅活性微生物群落多樣性
在酒醅主發酵期,細菌群落多樣性呈現先升后降趨勢,發酵7 d,細菌多樣性最高,發酵25 d細菌多樣性最低(圖2a);活性真菌群落物種多樣性呈上升趨勢,且主要集中在3~7 d(圖2b),這些結果與之前大多數報道的結果相似。

圖2 不同發酵節點酒醅活性細菌(a)和真菌(b)群落Shannon指數Fig.2 Shannon index of active bacterial (a) and fungal (b) communities in fermented grains at different fermentation times
2.3.2 酒醅活性微生物群落多樣性
由圖1可知,真菌界和細菌界總含量占可鑒定界含量的93%以上,表明優勢活性生物群落的動態變化主要由微生物群落變化驅動。在酒醅發酵過程中,不同發酵節點活性優勢生物群落的變化存在明顯的規律性(圖3)。


圖3 界(a)、門(b)、綱(c)、屬(d)、種(e)分類水平下酒醅微生物動態變化規律Fig.3 Dynamic changes of microbial communities in fermented grains at kingdom (a),phylum (b),class (c),genus (d) and species (e) levels
在界水平(圖3a),優勢微生物為細菌界和真菌界,隨著發酵時間延長(≤25 d),細菌界含量呈增加趨勢,由17.7%增至56.3%,發酵7 d后成為第一優勢界;而真菌界含量呈下降趨勢,由42.8%降至0.01%,表明真菌是酒醅發酵前3 d的主要活性微生物,而隨發酵延長細菌界成為優勢活性微生物。這主要是由于大多數真菌為好氧性微生物,隨著發酵時間延長,氧氣含量不斷減少且養分不斷消耗,導致真菌含量減少,這也與之前相關可培養結果一致;而一些厭氧或兼性厭氧、耐酸的微生物在發酵過程中相對含量不斷增加,且主要為乳酸菌。
在門水平(圖3b),優勢微生物門主要為Firmicutes(3 個節點酒醅中平均相對含量38.2%)和Ascomycota(13.8%),還有少量的擔子菌門(Basidiomycota,2.9%)和藍細菌門(Cyanobacteria,1.4%)。在發酵過程中,Firmicutes和Ascomycota含量的變化趨勢分別與細菌界和真菌界的變化趨勢一致,且其分別占細菌界和真菌界含量的90.2%和71.2%,表明這兩個門是驅動酒醅活性微生物群落變化的主要微生物。
在綱水平(圖3 c),酒醅發酵3 d 優勢微生物Saccharomycetes含量最高(32.8%),其次為Bacilli(14.6%)、Agaricomycetes(5.5%)和Clostridia(0.01%)。隨著發酵時間延長,Saccharomycetes相對含量明顯下降,由32.8%降至0.04%,且含量下降主要發生在前7 d,由32.8%降至3.9%。而Bacilli含量在發酵過程中不斷增加,在發酵第7天,已成為酒醅第一優勢綱(42.7%),隨后又增加至53.7%。Agaricomycetes在發酵過程中呈現逐漸下降趨勢,由5.5%降至0.03%。Clostridia含量呈現先升后降的趨勢,在發酵7 d 達到最大值(2.5%)。在本實驗前期研究中,發現Clostridia是窖泥中的含量高且具有重要潛在功能微生物類群,有助于穩定窖泥微生態并能合成濃香型白酒重要呈香化合物及其前體(尤其是多種脂肪酸)。此外,采用基于DNA水平分子生物學方法也發現Clostridia是酒醅中的優勢微生物,并檢測到多種梭菌綱中的屬,如、和等。
在屬水平(圖3d),隨著發酵時間延長,酒醅優勢活性微生物、、和逐漸減少,其中含量下降主要發生在前7 d,由28.4%降至1.6%。、和含量呈先升后降趨勢,尤其是后兩個屬,其含量分別由第3天的0.1%和0.04%增加至第7天的6.8%和4.4%。在發酵過程中不斷上升,在發酵第25天成為唯一優勢活性微生物屬,而上述其他微生物在發酵25 d含量降至0.01%~0.46%之間。
圖3e展示了酒醅中含量前50的種(含unidentified)隨時間變化的特征,其中發酵7 d和發酵25 d聚為一簇,而3 d酒醅單獨聚為一簇。這表明在酒醅主發酵期,隨著發酵時間的延長前50活性物種組成逐漸趨于一致,并與發酵初期的組成存在明顯的差異。此外,根據其在不同發酵節點的酒醅含量差異可以分為3 類,其中第i類(共7 種)是隨發酵時間延長含量逐漸減少的物種,尤其是釀酒酵母()相對含量由27.8%逐漸下降至1.6%(7 d)和0.04%(25 d),以及樹干畢赤酵母()相對含量由4.0%下降至2.1%(7 d)和0.02%(25 d);第ii類是發酵7 d含量最高的物種(共14種),主要包括非屬中的物種,如變形鏈球菌()、蠟樣芽孢桿菌()和柔嫩梭菌()等,尤其是相對含量在7 d高達8.4%;第iii類生物(28 種和unidentified)主要是隨發酵時間延長而增加的物種,其主要為中的物種,且大多數物種是第一次檢測到,如和等。綜上,在主發酵期,隨著發酵時間延長,等真菌物種含量逐漸減少;大多數非屬(如及等)中的物種的含量先升后降,在發酵7 d達到最高;而大多數中的物種含量一直在增加,從而在發酵過程中得到了富集。
2.4 DEG分析
2.4.1 DEG篩選結果
圖4展示了不同發酵節點酒醅中顯著性DEG數量及變化情況,從3 d vs 7 d、7 d vs 25 d、3 d vs 25 d的酒醅樣品中測得的DEG數量分別為6 895、2 640 個和6 124 個。這說明3 d酒醅與7 d和與25 d酒醅均有較多的DEG數量(>6 100 個),而7 d酒醅與25 d酒醅的DEG數量相對較少,表明酒醅中活性微生物群落功能明顯變化主要發生在發酵前7 d。

圖4 3 d vs 7 d(a)、7 d vs 25 d(b)、3 d vs 25 d(c)酒醅之間的DEG火山圖Fig.4 Volcano plot of differentially expressed genes between 3-vs 7-day (a),7-vs 25-day (b),and 3-vs 25-days (c) fermented grains
此外,與7 d酒醅樣品相比,3 d酒醅樣品顯著下調4 284 個(占該組DEG數量的62.1%)和上調2 611 個(37.9%)DEG。同理,25 d酒醅樣品相比7 d酒醅樣品顯著下調2 023 個(占該組DEG數量的76.6%)和上調617 個(23.4%)DEG。綜上,隨著發酵時間延長,整個主發酵過程中下調的DEG數量明顯多于上調數量,表明酒醅微生物群落功能多樣性及代謝活性可能逐漸減小,同時也有少部分功能得到增強。
2.4.2 DEG的GO富集分析
由圖5a、b可知,發酵3 d酒醅較7 d酒醅下調和上調DEG富集的前30 個功能條目均具有顯著性,表明發酵前期微生物的上述代謝功能均存在顯著變化。其中7 d較3 d酒醅(圖5a)顯著上調DEG顯著富集的前30功能條目中與生物過程相關的條目20 個,DEG數量共為1 340 個,其中每個條目中DEG數量為9~103 個;與細胞組成相關的條目1 個,含14 個DEG;與分子功能相關的條目9 個,DEG數量共為1 194 個,其中每個條目中基因的數量為5~988 個unigenes。7 d較3 d酒醅顯著下調的DEG顯著富集的前30功能條目中與生物過程相關的條目6 個,DEG數量共為6 997 個;與細胞組分相關的條目24 個,DEG數量共為47 749 個(圖5b)。25 d較7 d酒醅(圖5c)中顯著上調DEG富集的前30功能條目中僅有催化活性功能條目顯著性富集,而顯著下調DEG(圖5d)富集的前30功能條目均呈顯著性富集。其中25 d較7 d酒醅(圖5c)中顯著上調DEG富集的前30功能條目中與生物過程相關的26 個條目,DEG數量共為1 194 個,與分子功能相關的條目4 個,DEG數量共為239 個;顯著下調DEG富集的前30功能條目中,與生物過程相關的條目13 個,DEG數量共為3 426 個;與分子功能相關的條目3 個,DEG數量共為333 個;與細胞組分相關的條目14 個,DEG數量共為6 629 個。綜上,不同發酵節點酒醅中顯著上調或上調DEG富集的前30功能條目隨著發酵時間延長存在一定的動態變化規律:1)發酵7 d相對3 d上調DEG富集的每個功能條目相關基因總數量(共2 548 個)遠高于25 d相對7 d上調DEG(共706 個),且富集的條目均集中在與生物過程和分子功能相關的條目,尤其是生物過程;2)發酵7 d相對3 d下調DEG富集的每個功能條目相關基因總數量(共54 746 個)遠高于25 d相對7 d下調DEG(共10 388 個),其中富集的條目均集中在與細胞組分和生物過程相關的條目,尤其是細胞組分。
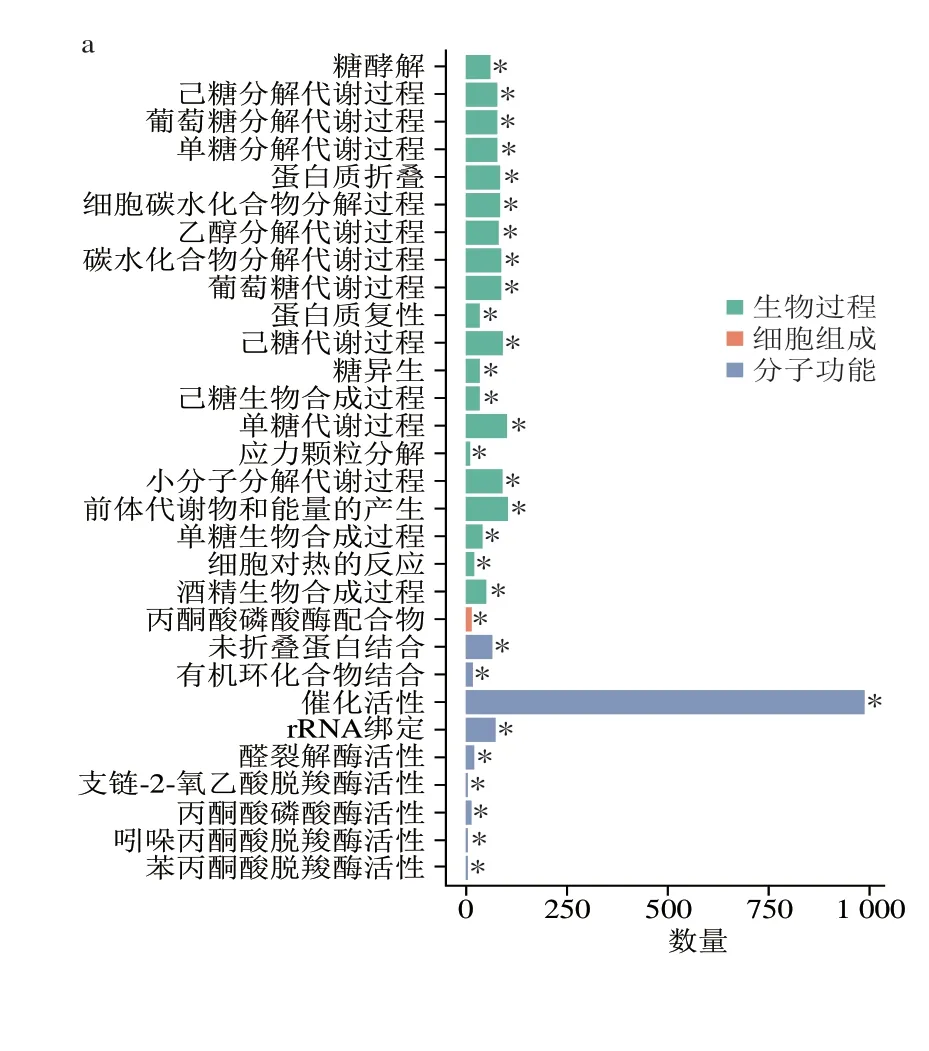

圖5 3 d vs 7 d酒醅下調(a)和上調(b)基因、7 d vs 25 d酒醅之間下調(c)和上調(d)基因的GO富集柱狀圖Fig.5 Significantly enriched GO terms of down-regulated (a) and up-regulated (b) genes between 3 d vs 7 d fermented grains,and downregulated (c) and up-regulated (d) genes between 7 d vs 25 d fermented grains
由圖5可知,隨著發酵時間延長,酒醅生物上調和下調DEG富集的前30功能條目均存在一定的差異。發酵到7 d,上調DEG顯著富集條目中包括糖酵解過程、葡萄糖分解代謝過程、醇類物質合成過程、細胞對熱反應等生物過程,以及催化活性和丙酮酸磷酸酶活性等分子功能等30 個條目均得到了顯著性富集,其中與糖和醇直接相關的生物過程有14 個條目共含996 個基因(圖5a),占所有生物過程條目中基因總數量的74.3%(996/1 340)。從發酵第7天到第25天,上調DEG在富集的前30 個條目中僅有催化活性功能條目具有顯著性,其中與糖和醇直接相關的生物過程有9 個條目共含172 個基因,與核酸及氨基酸代謝直接相關的生物過程有15 個條目共含188 個基因(圖5c)。發酵7 d,下調DEG富集的30 個條目均具有顯著性,且幾乎均與細胞組分或與其相關的生物過程相關。從發酵第7天到第25天,下調DEG富集的30 個條目均具有顯著性,且主要細胞組分生物過程,含14 個條目6 629 個基因,同時也發現與蛋白質代謝過程及醇類合成過程的條目得到了顯著富集。這些結果表明:1)發酵7 d后,酒醅微生物菌群參與糖和醇類化合物代謝的功能可能會顯著增強,這也與諸多文獻報道結果相似,即發酵1 周后還原糖含量明顯下降,而乙醇含量明顯上升;2)從發酵第3天至發酵第25天,酒醅微生物豐度可能會出現顯著降低,這也Wang Xueshan等研究結果一致,酒醅原核微生物豐度在發酵3 d后呈顯著下降趨勢。
3 結論
采用宏轉錄組技術從RNA水平對濃香型不同發酵節點酒醅中活性生物群落及其功能變化規律進行了解析,主要結論包括:酒醅主發酵期濃香型酒醅中可鑒定的活性生物群落主要由細菌和真菌組成,共檢測到2 034 個可鑒定的細菌和真菌屬,在很大程度上豐富了濃香型酒醅微生物群落多樣性內涵;揭示了主發酵期濃香型酒醅中可鑒定的不同分類水平下活性生物群落組成及其變化規律,如Firmicutes和Ascomycota是酒醅中主要優勢菌門,其中前者呈上升趨勢,而后者呈下降趨勢;主發酵期濃香型酒醅微生物群落功能多樣性及代謝活性可能逐漸減小,同時也有少部分功能得到增強,且其明顯變化主要發生在發酵前7 d;發酵7 d后,酒醅微生物菌群參與糖和醇類化合物代謝的功能可能會顯著增強;發酵3~25 d,酒醅活性微生物豐度可能會出現持續顯著降低。本研究從活性微生物群落結構和功能角度有助于進一步揭示濃香型白酒固態釀造機制,以期為濃香型白酒生產控制及定向群落組裝技術開發提供一定的理論支持。
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學生數理化(高中版.高考數學)(2020年1期)2020-02-20 13:23:44
經濟技術協作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
