光照強度驅動典型陰生植物三七的生理生態響應特征
2022-05-26 10:02:04雙升普張金燕武洪敏孟珍貴李龍根陳軍文
生態學報 2022年9期
雙升普,張金燕,寸 竹,武洪敏,孟珍貴,李龍根,陳軍文,*
1 云南農業大學西南中藥材種質創新與利用國家地方聯合工程研究中心,昆明 650201 2 云南農業大學云南省藥用植物生物學重點實驗室,昆明 650201 3 云南農業大學農學與生物技術學院,昆明 650201
光作為光合作用能量的來源,其強度會影響植物的形態建成、光合生理及生物量的產生與分配等過程,從而影響植物生長發育進程[1—2]。而根據植物對光照強度的需求性不同,可以分為喜光植物與喜陰植物[3]。然而在自然界中,光照強度是高度異質化的,植物必須以固著的方式響應光照環境的異質性[4—5]。在沒有發生遺傳變異的情況下,植物主要通過表型可塑性適應異質化的光環境[6—7]。表型可塑性是植物在形態建成、生長特征和生理功能等方面發生變化從而響應異質環境的一種生態策略[8—10]。可塑性指標的大小可以反映植物對環境變化適應能力的強弱[11—12]。因此,植物表型的可塑性是植物光響應研究的主要指標之一。
光合生理是最先被光照強度驅動來響應變化的生理特征[13—16]。喜光植物通過較高的凈光合速率(Pn)、光飽和點(LSP)和光補償點(LCP)等來增強自身潛在的光合能力,因此能忍受較高的光照強度[17—19]。低光下,喜光植物風吹楠(Horsfieldiaglabra)Pn較低,此時其通過降低LCP和暗呼吸速率(Rd)來減少消耗從而適應遮陰環境;而在高光下可通過增加Pn來適應高光環境[13]。但是,較高的Pn意味著單位面積上需要投入更多的葉綠體、光合蛋白及有機物等物質來構建較厚的葉片[20—21]。喜陰植物由于光合能力較弱,導致用于構建葉片厚度的生物量較少,因此其高光適應能力較弱。另一方面,喜陰植物自身潛在的Pn、LSP、LCP以及Rd相對較低[17—19],當光照強度發生變化時,其光合生理如何在較小的變化幅度內實現最優調整是值得研究的問題。
植物對不同光照強度的響應最終表現在形態建成上。當環境光強減弱時,植物通過增大植株冠幅和增加單片葉面積等方式向著更有利于捕獲光能的方向發生改變;而在高光照條件下,植物通過減小單株葉面積和葉片數量等地上部分形態特征來減少光能的吸收[10,22—23]。此外,光照強度對植物地上部分的調節會影響植株碳的獲取,從而調節根系生物量的積累,使其更好的適應地上部光環境的變化[1]。前人研究發現喜陰植物可通過形態特征的變化來較快地響應不同的光照環境,但其形態可塑性與喜光植物相比變化幅度較小[10]。因此,了解喜陰植物形態建成如何在有限的變化范圍內較快響應異質光環境對于分析其響應策略是非常必要的。
生物量的積累和分配作為植物在異質光環境中內在和外在聯系的橋梁,其變化會影響相對生長速率(RGR),是植物響應光照環境的另一重要策略,尤其是不同光照環境下葉生物量比(LMR)、葉面積比(LAR)、比葉面積(SLA)等生物量分配的改變[24—26],其中SLA是與RGR變化相關的主要因子[27—28]。在低光環境下,較大的SLA使得有效葉面積增加,光合碳收獲增強,最終引起植物RGR增加[29]。然而SLA對光的適應性調整也是有限度的,當遮陰過度時喜陰植物總的生物量減少,此時分配到地上部分的生物量雖然比例有所增加但數量卻有所減少,使得植物捕獲的光能相對較少,導致RGR降低。生產實踐上,都會追求植物的最大RGR,以達到高產目的,因此,當環境光強發生變化時,陰生植物生物量如何優化分配調整來使RGR尤其是經濟器官的RGR保持較大是十分重要的。
三七(Panaxnotoginseng)為五加科人參屬多年生的典型陰生植物,以根和根狀莖入藥,是中國傳統的名貴中藥材,在種植過程中需要采用遮陰栽培[30—31]。傳統道地產區的三七種植中有“三分陽,七分陰”之說,認為三七在30%透光率(LT)下生長較好,但試驗證據相對缺乏。本課題組前期研究發現,當透光率為9.6%—11.5%時,三七具有較高的Pn,同時在1%透光率下三七也能生存,表現出較強適應低光的能力[31—33]。此外,有研究發現,一年生三七通過提高Pn、降低蒸騰速率(Tr)和氣孔導度(Gs)以及降低株高和葉面積來適應高光照環境[34—35],但實驗使用的材料是處于種苗階段的一年生三七。本課題組研究還發現,兩年生三七會通過降低根冠比(RSR)、根生物量比(RMR),增加莖生物量比(SMR)和LMR以獲取更多的光能,從而更好地適應低光照環境;而在高光照下則是通過葉片變厚變小來減少光能捕獲,并增加根重比來適應環境[36—38]。然而,以往的相關工作都是對不同生長光照強度下三七形態特征或生理生態特性中某一方面的研究,從形態、生理和生長角度綜合深入分析三七對長期光照環境變化的響應特征的研究相對較少,且以前相關研究生長環境光強設置的范圍較小。因此,本文以兩年生三七為實驗材料,設置一系列的生長環境光照強度(46.5% LT、21.8% LT、9.70% LT、5.10% LT和2.80% LT),對生長于不同光照強度下三七的形態特征、生物量積累及分配和光合生理特性進行分析,并進一步進行相關性、可塑性和主成分分析,進而闡明三七對不同光照環境的響應策略,為三七的優質高效種植提供理論依據。
1 材料和方法
1.1 試驗地概況
試驗設于文山州硯山縣盤龍鄉(104°32′E,23°53′N)苗鄉三七實業有限公司三七科技示范園內。硯山縣位于云南省東南部,103°35′—104°45′E、23°18′—23°59′N之間,海拔1539 m,年平均降雨量1200 mm,年平均氣溫約16 ℃,最冷月平均溫度9 ℃,最熱月平均溫度21 ℃,全年無霜期大于300 d,日均氣溫10 ℃以上,積溫為5000 ℃,屬于亞熱帶大陸性季風氣候[39]。硯山縣是三七的主要傳統產區和道地產區,同時也是三七最適宜生長的地區之一,被譽為“中國三七之鄉”。
1.2 試驗材料及設計
在空曠地使用三七專用遮陽網搭建5種不同透光率的蔭棚,蔭棚長4 m、寬4 m、高2 m,設有紗網,通風透氣。選擇晴天,使用Li-190R 量子傳感器(棚內和棚外各三個)和Li-1500 光量子記錄儀(Li-Cor,USA),于6:00—19:00每隔10 s采集一次每個蔭棚內的光照強度;同時測量全日照強度作為對照。以測量時間段內蔭棚光照強度(三個量子傳感器的平均值)占同一時刻透光率強度百分比的平均值作為三七生長環境光強的指標;經測量,5個處理的平均環境光照強度分別為:46.5% LT、21.8% LT、9.70% LT、5.10% LT、2.80% LT(圖1)。在前期研究發現,透光率為9.6%—11.5%時三七生長發育良好,透光率高于30%三七就會受到顯著的高光抑制,而透光率只有3%左右時三七將受到明顯的低光限制[36],因此,本實驗中9.70%透光率代表前期發現的適宜光照條件,5.10%、2.80%的透光率代表低光條件,46.5%和21.8%的透光率代表高光條件。實驗采取盆栽進行,花盆內徑30 cm、深25 cm,盆栽土壤為當地生土(紅壤),其土壤基本理化性狀如表1所示。于1月份選取生長良好、大小一致的三七種苗進行移栽,每個處理下劃分為15個重復小區,每個小區放置5盆,每盆3株。試驗期間三七常規田間管理措施統一按文山三七科技示范園的三七規范化種植技術規程進行。

圖1 不同透光率和全日照下的光量子通量密度Fig.1 Representative diurnal time courses of photosynthetic photon flux density in different light transmittance (LT) and the full sunlight
1.3 測定指標及方法
1.3.1形態指標的測量
于12月份三七(兩年生)收獲期,將不同透光率蔭棚下的三七進行取樣。取樣時,將三七整株取出,用水洗凈,每個處理選15株(每個小區1株)進行測量,每株重復3次。用電子數顯游標卡尺測量莖粗(mm);用直尺測量株高(cm)、每片小葉的長和寬(測葉片最寬的地方,cm),將葉片看作橢圓形,計算每個葉片的葉面積(葉面積=葉長×葉寬×0.75 cm2)[40],并統計出三七的單株葉面積(cm2);然后統計植株的葉片數及分枝數。
1.3.2生物量及分配的測量
每個處理選15株三七(每個小區1株),然后將根、莖、葉分開放在105 ℃下的烘箱中殺青1 h,然后在60 ℃下烘干至恒重。用電子天平稱量根生物量(根重,g)、莖生物量(莖重,g)、葉生物量(葉重,g);并統計出總生物量(Mt)、根生物量比(RMR,根重/植株總重)、莖生物量比(SMR,莖重/植株總重)、葉生物量比(LMR,葉重/植株總重)、葉面積比(LAR,總葉面積/植株總重cm2/g)和比葉面積(SLA,總葉面積/葉重cm2/g)。并且在移栽時也需要對三七種苗進行取樣,烘干后測量其總生物量(M0),用于計算相對生長速率(RGR)[41]:
RGR=(lnMt-lnM0)/t
其中,Mt表示兩年生三七的總生物量(g),M0表示移栽時測定的三七種苗的總生物量(g),t表示兩次測定的時間間隔(d)。
1.3.3氣體交換參數的測量
于生長旺盛期,使用Li-6400XT光合-熒光記錄儀(Li-Cor,USA)測定不同蔭棚下三七葉片的光響應曲線。使用CO2注入系統將CO2濃度控制為400 μmol/mol,夾上葉片后于500 μmol m-2s-1光強下穩定一段時間,待數據穩定后啟動自動測量程序,光照強度從大到小依次設為:800、500、400、300、200、150、100、80、60、40、20、10、0 μmol m-2s-1,每個光強下穩定2—3 min,采集氣體交換參數值。以光照強度(Photosynthetic photon flux density,PPFD)為橫軸,Pn為縱軸繪制光響應曲線(Pn-PPFD曲線)。據Bassman的方法擬合Pn-PPFD曲線方程[42]:
Pn=Pn-max-Pn-maxC0e-αPPFD/Pn-max
式中,Pn-max為最大凈光合速率,α為表觀量子效率(AQY),C0為度量弱光下凈光合速率趨近于0的指標。從中可得到單位面積最大凈光合速率(Pn-max/area)、單位面積氣孔導度(Gs/area)、單位面積暗呼吸速率(Rd/area)=Pn-max-Pn-maxC0、單位面積光補償點(LCP/area,μmol m-2s-1)=Pn-maxln(C0)/α。然后基于SLA計算單位質量最大凈光合速率(Pn-max/mass)=(Pn-max/area)×SLA×1000/10000、單位質量氣孔導度(Gs/mass)=(Gs/area)×SLA×1000/10000、單位質量暗呼吸速率(Rd/mass)=(Rd/area)×SLA×1000/10000、單位質量光補償點(LCP/mass,nmol g-1s-1)=(LCP/area)×SLA×1000/10000。實驗中使用統一校準后隨機分配的5臺Li-6400XT光合儀(Li-Cor,USA)同時測量5個處理,每個處理選取15株(每個小區1株)進行測量,每株選取1葉片,葉片空間取向和角度盡量一致。
1.4 數據處理
用SPSS 25.0軟件對數據進行單因素方差分析(One-way ANOVA)及差異顯著性分析(n=15),用Pearson法對三七RGR與各指標間的相關性進行分析;用Sigmaplot 14.0和Origin 2020b軟件作圖。
2 結果與分析
2.1 不同光照強度對三七生長及形態特征的影響
在高光和低光下,三七各形態特征參數都有所降低(圖2)。其中,株高、莖粗和單株葉面積在透光率為9.70% 時最高,且顯著高于46.5% 和2.80% (P<0.05);葉片數量和分枝數量在透光率為9.70% 的條件下最多,且顯著高于46.5% (P<0.05)。總的來看,在透光率為9.70% 時三七形態特征最好,高光或低光都會顯著降低其形態的生長發育。
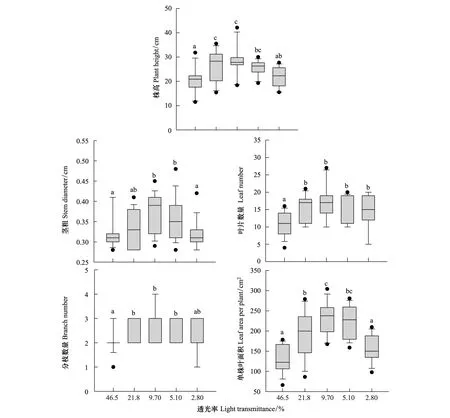
圖2 不同透光率對三七株高、莖粗、葉片數量、分枝數量和單株葉面積的影響Fig.2 Effects of different light transmittance on plant height,stem diameter,leaf number,branche number and leaf area per plant of P. notoginseng不同字母表示差異顯著(P<0.05)
2.2 不同光照強度對三七生物量分配的影響
高光和低光條件下三七各部分生物量都有所降低(圖3)。單株根生物量、單株莖生物量、單株葉生物量和單株總生物量在不同透光率下的變化趨勢一致,都在透光率為9.70%時最高,其次是21.8%,這兩種條件的各部分生物量都顯著高于其他透光率(P<0.05),且透光率為2.80%時各部分生物量最少。總的看來,透光率為9.70%條件下的各部分生物量和單株的總生物量都具有最大值,而透光率為2.80%時最低,并且顯著低于其他幾個透光率(P<0.05),表明低光下生物量積累較少。

圖3 不同透光率對三七單株根生物量、單株莖生物量、單株葉生物量和單株總生物量的影響Fig.3 Effects of different light transmittance on root biomass per plant,stem biomass per plant,leaf biomass per plant and total biomass per plant of P. notoginseng
由圖4可知,三七LAR和SLA都在透光率為2.80%下最大,且顯著高于其他幾個透光率(P<0.05),表明低光下植物會通過增加LAR和SLA來獲取更多的光能。另外,不同透光率下三七RMR、SMR和LMR是存在一定差異的。其中,RMR隨著透光率的降低而降低,而SMR和LMR則是隨著透光率的降低而逐漸增加,且三者在透光率為46.5%、21.8%和9.70%時差異都不顯著,但都顯著低于其他兩個透光率;并且RGR在透光率為9.70%時最大,并顯著高于其他幾個透光率(P<0.05)。表明低光和高光都會影響植物各部分生物量的分配比例,并抑制植物生長。

圖4 不同透光率對三七生物量分配的影響Fig.4 Effects of different light transmittance on biomass allocation of P. notoginseng
2.3 不同光照強度對三七光合能力的影響
由圖5可以看出,不同透光率下三七Pn-max/area、Gs/area、Rd/area、LCP/area和Pn-max/mass、Gs/mass、Rd/mass、LCP/mass是存在差異的。與透光率9.70%相比,高光(46.5% LT和21.8% LT)水平下的Pn-max/area、Gs/area、LCP/area和Pn-max/mass、Gs/mass、LCP/mass略高,但與對照差異不顯著;低光(5.10% LT和2.80% LT)水平下,上述參數則顯著低于9.70% LT下的(P<0.05),且隨著透光率的降低而減小。而Rd/area和Rd/mass都是隨著透光率的降低而逐漸上升,與透光率9.70%相比,除21.8% LT條件下的Rd/mass與其差異性不顯著外,其余參數在高光下都顯著下降(P<0.05);而低光下則都顯著較高(P<0.05)。這些結果表明9.70%的透光率下三七的光合能力較好,而高光下三七會制造和消耗較多的有機物,低光下有機物的制造和消耗較少。

圖5 不同透光率對三七光合生理的影響Fig.5 Effects of different light transmittance on photosynthetic physiology of P. notoginsengPn-max/area:單位面積最大凈光合速率Maximum net photosynthetic rate unit area;Gs/area:單位面積氣孔導度Stomatal conductance unit area;Rd/area:單位面積暗呼吸速率Dark respiration rate unit area;LCP/area:單位面積光補償點Light compensation point unit area;Pn-max /mass:單位質量最大凈光合速率Maximum net photosynthetic rate unit mass;Gs/mass:單位質量氣孔導度Stomatal conductance unit mass;Rd/mass:單位質量暗呼吸速率Dark respiration rate unit mass;LCP/mass:單位質量光補償點Light compensation point unit mass
2.4 不同光照強度下三七形態和生理特征與RGR的相關性分析
2.4.1形態特征與RGR的相關性分析
RGR與株高、莖粗以及單株葉面積呈正相關關系,相關性均達到極顯著水平(P<0.01),但相關性系數較低,分別為0.116、0.093和0.270,其中RGR與單株葉面積相關性最大,而與葉片數量的相關性則不顯著(圖6)。這些結果說明,在形態特征方面,單株葉面積是影響三七生長發育的主要因素,但形態特征各參數的相關性系數相對較小,表明與光合生理及生物量分配相比,形態特征對三七生長發育的影響較弱。

圖6 三七株高、莖粗、葉片數量、單株葉面積與相對生長速率的相關性Fig.6 Correlation between plant height,stem diameter,leaf number,leaf area per plant and relative growth rate (RGR) of P. notoginseng
2.4.2生物量分配與RGR的相關性分析
RGR與LAR、SLA、SMR、LMR呈負相關關系,相關性均達到極顯著水平(P<0.01),其相關系數分別為0.493、0.461、0.390、0.472,而與RMR則是極顯著的正相關關系(P<0.01)相關系數為0.553。說明RGR與RMR相關性最大,和LAR的相關性次之(圖7)。總的來看,生物量分配對三七的生長發育有著較大的影響,其中又以控制分配到根部生物量的RMR和調控有機物生產和積累的LAR的影響最大。說明生物量分配中三七主要通過RMR和LAR來響應不同光照強度。

圖7 三七葉面積比、比葉面積、根生物量比、莖生物量比、葉生物量比與相對生長速率的相關性Fig.7 Correlation between leaf area rate,specific leaf area,root mass ratio,stem mass ratio,leaf mass ratio and relative growth rate of P. notoginseng
2.4.3氣體交換參數與RGR的相關性分析
由圖8可知,RGR與Pn-max/area、Gs/area、LCP/area、Pn-max/mass、Gs/mass、LCP/mass呈正相關關系,相關性均達到極顯著水平(P<0.01),相關系數分別為0.581、0.558、0.574、0.350、0.274、0.300,而與Rd/area、Rd/mass則是極顯著的負相關關系(P<0.01)相關系數分別是0.472、0.333;說明RGR與Pn-max/area相關性最大,和Gs/area、LCP/area的相關性次之。總之,三七的RGR與Pn、Gs、LCP、Rd有很大的關聯,說明光合生理是植物響應光照環境的一個重要策略,是影響三七生長發育的主要因素。

圖8 三七單位面積最大凈光合速率(Pn-max/area)、單位面積氣孔導度(Gs/area)、單位面積暗呼吸速率(Rd/area)、單位面積光補償點(LCP/area)和單位質量最大凈光合速率(Pn-max/mass)、單位質量氣孔導度(Gs/mass)、單位質量暗呼吸速率(Rd/mass)、單位質量光補償點(LCP/mass)與相對生長速率的相關性Fig.8 Correlation between maximum net photosynthetic rate unit area (Pn-max/area),stomatal conductance unit area (Gs/area),dark respiration rate unit area (Rd/area),light compensation point unit area (LCP/area),maximum net photosynthetic rate unit mass (Pn-max /mass),stomatal conductance unit mass (Gs/mass),dark respiration rate unit mass (Rd/mass),light compensation point unit mass (LCP/mass) and relative growth rate of P. notoginseng
2.5 不同光照強度下三七形態和生理特征的可塑性
由圖9可知,在所有性狀指標中,莖粗的可塑性指數最小,單株根生物量可塑性指數最大;從形態特征、生長特征和生理特征三方面來看,形態特征中沒有一個參數的可塑性指數大于0.5,其可塑性最小,生理特征參數的可塑性指數大于0.5的最多,其可塑性最大;而從形態特征分組可以看出,各參數的可塑性指數均小于0.5,其中單株葉面積的可塑性最高;在生長特征分組中,單株根生物量及LAR的可塑性指數大于0.5,其中單株根生物量的可塑性最大,RGR的可塑性最小;生理特征中可塑性指數大于0.5的參數最多,有5項(Pn-max/area、Gs/area、Rd/area、LCP/area、Rd/mass)占比62.5%,其中Rd/area的可塑性最大,Gs/mass的可塑性最小。這些結果表明三七主要是通過改變光合生理、根的生物量以及LAR來響應光照強度的變化。
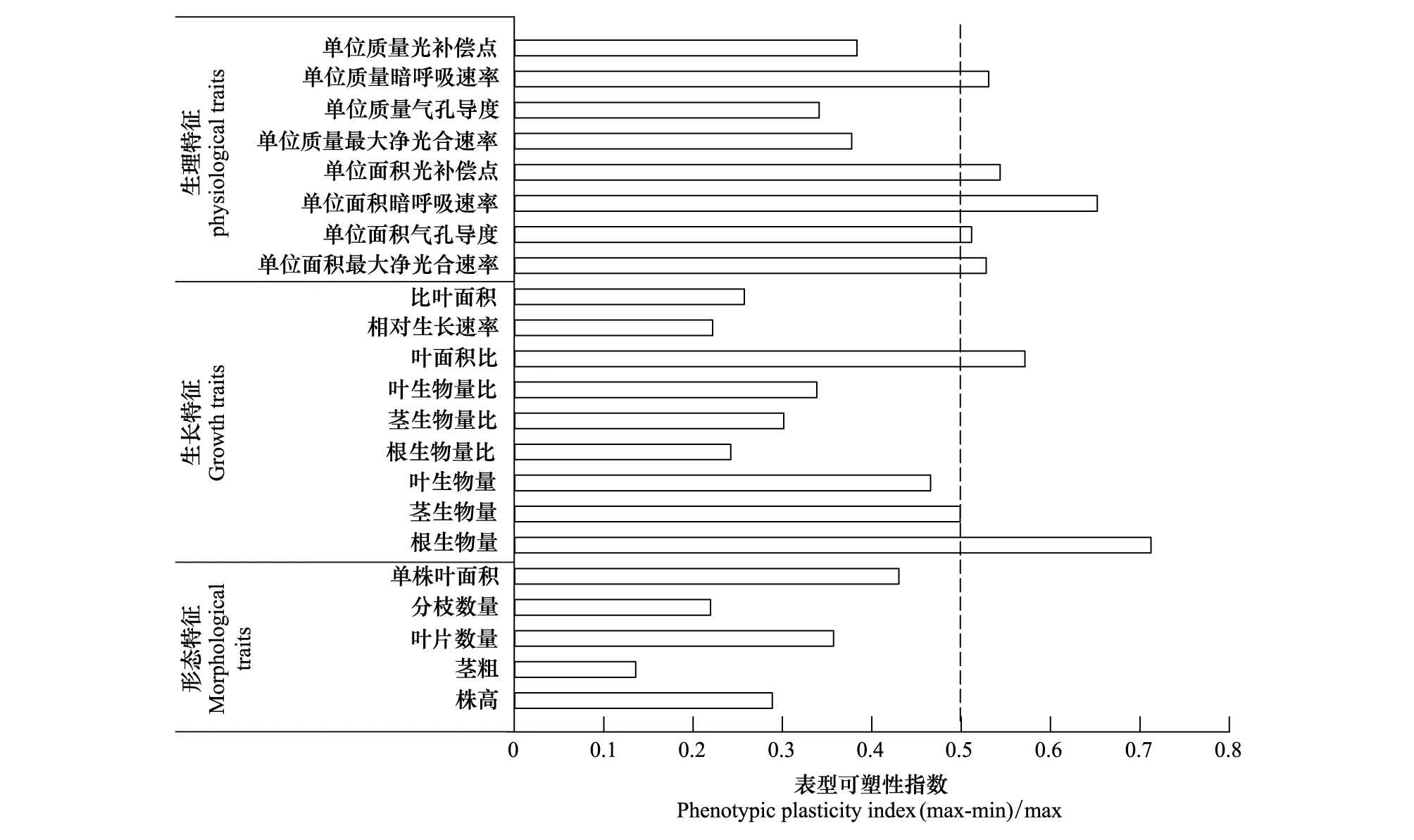
圖9 三七形態特征、生長特征和生理特征等各參數的表型可塑性指數Fig.9 Phenotypic plasticity index of parameters of morphological characteristics,growth characteristics,and physiological characteristics of P. notoginseng
2.6 不同光強響應特征的主成分分析
對不同光照強度下三七的23個形態特征、生長特征和生理特征參數進行主成分分析,得到各主成分對三七響應不同光強的特征值和貢獻率(表2)。從表中可以看出,PC1和PC2的累計貢獻度達70.61%,且特征

表2 各成分初始特征值及累積貢獻率Table 2 The initial eigenvalues and the accumulated variance contribution of each component
值都大于1,因此這兩個主成分能有效反映參數特征和有效解釋三七的響應特征。
由圖10可知,PC1中光合生理特征參數(Pn-max/area、Gs/area、Rd/area、LCP/area、Pn-max/mass、Gs/mass、Rd/mass、LCP/mass)和生長特征參數(單株根生物、單株總生物量、RMR、SMR、LMR、LAR、SLA、RGR)權重系數較大;PC2中生長特征參數(單株莖生物量、單株葉生物量)和形態特征參數(株高、莖粗、葉片數量、分枝數量、單株葉面積)權重系數較大。實驗點在PC1/PC2平面上較好的集聚為五簇,分別代表五個不同光強處理。同時PC1非常明顯的將透光率2.80%與其他四個透光率分離開,將透光率5.10%與透光率46.5%、21.8%、9.70%明顯分離開,而透光率46.5%、21.8%、9.70%三者間沒有明顯的區分,表明低光與其他幾個光強之間有著明顯的差異,而高光與透光率9.70%的差異較小。由載荷系數可以看出,Pn-max/area、Gs/area、LCP/area、RGR、根生物量、總生物量與PC1有著正相關關系,且對PC1的貢獻較大;而LAR、Rd/area對PC1也有著較大的貢獻,但為負相關關系;葉片數量、分枝數量、單株葉面積與PC2為負相關關系,并且對其貢獻較大。并且Pn-max/area、Gs/area、LCP/area、RMR、根部生物量及總生物量與RGR相關性較高,且為正相關關系,而Rd/area、LAR與RGR則有著較高的負相關關系,說明在不同光照強度下三七主要靠調整光合生理特征、LAR、根生物量以及RMR來響應變化。

圖10 不同透光率下三七形態和生理指標的主成分分析Fig.10 Principal component analysis of morphological and physiological indexes of P. notoginseng under different light transmittance LAR:葉面積比Leaf area rate;SLA:比葉面積Specific leaf area;RMR:根生物量比Root mass ratio;SMR:莖生物量比Stem mass ratio;LMR:葉生物量比Leaf mass ratio;圖中點的顏色代表不同透光率;橢圓為不同透光率下各參數的置信區間;箭頭代表各指標與主成分的關系
3 討論
3.1 高光環境下陰生植物三七采取快速碳獲得和碳消耗的冒險響應策略
形態特征作為判斷植物生長發育狀況的一項重要指標,可以直觀反映植株對不同強度光照環境的適應性[43]。植物形態特征的改變是通過生物量積累和局部特化的生物量分配變化來實現的,因此,生物量在各器官間的分配反映植物生長策略的調整[22,44]。高光環境下陰生植物三七LMR、SMR、SLA和LAR較低,RMR較高(圖4),這與魏巍等研究發現高光下陽生植物薇甘菊(Mikaniamicrantha)SLA 、LAR降低一致[24]。較低的SLA和LAR使葉片變厚變小,即單位生物量所承載的光合葉面積減少,使其捕光面積降低[45]。較厚的葉片在強光下單位面積葉片的光合能力較強[46],從而三七在高光下保持較高的Pn(圖5),消耗更多的光能,生產更多的有機物。同時高光下三七Rd較大(圖5),使得有機物消耗較多,從而維持Pn在一個較高水平來利用更多的光能,防止光合機構遭受破壞[42,47],這些特征有利于植物在強光下保持較高的生存能力,提高對強光的適應性。與此同時,SLA和LAR降低表明植物生物量分配發生變化,高光下較多的生物量分配到三七地下部分,抑制地上部分的生長,提高RMR,有利于吸收更多的水分和無機物,為植物蒸騰和光合作用提供更多的原料,這與葉片變小時三七蒸騰失水減少相互協調,成為陰生植物三七響應高光環境的重要策略[45,48]。總的來看,高光下三七較高的Pn可以制造更多的有機物,而Rd較大則會消耗大量的光合產物,并且此時生物量積累較少(圖3),這說明高光下陰生植物三七同時進行快速碳獲得和碳消耗的冒險策略[49]。
3.2 低光下陰生植物三七采取緩慢碳獲得和碳消耗的保守響應策略
低光下由于光能不足,導致碳獲得顯著下降(圖5),此時各部分的生物量和總生物量的積累都顯著降低(圖3),這與陽生植物大豆(Glycinemax)在遮陰處理下光合作用減弱,導致生物量積累減少的結果相一致[50]。遮陰過度時,三七吸收的光能無法滿足其正常光合作用所需,因此Pn降低,產生和積累的有機物減少,營養生長受到抑制,最終導致株高和葉面積降低(圖2),此時Rd降低,有機物的消耗減少,這與較低的光合速率相匹配。此外,三七通過減少根生物量的分配,將更多的資源分配到地上的莖和葉部分,增加SLA和LAR(圖4),植株葉面積增大,使其在低光下捕獲更多的光能,合成更多的光合產物來提高其耐陰性,以此來實現對弱光環境的適應[51—53]。另一方面,LCP的高低可以反映植物對弱光的利用能力,低光下三七具有較低的LCP,降低了對光強利用和凈碳獲得的閾值,這也是植物對低光環境的一種響應策略。因此,低光下陰生植物三七通過減少呼吸消耗來應對較少的光合有機物產量,從而維持碳平衡,此時,三七采取緩慢的碳獲得和碳消耗的保守策略來響應低光環境。
3.3 陰生植物三七光合生理活性及生物量分配的改變是其響應光強變化的主要策略
RGR與植物形態特征、生理特征及生物量的積累與分配等有著密切關系。前人研究發現,RGR受光合碳獲得和LAR的共同影響,較高的Pn和LAR均可提高RGR[54]。在本研究中,Pn、Gs和LCP與RGR呈正相關關系,Rd與RGR呈負相關關系(圖8),這與前人研究發現,陽生植物麻櫟(Quercusacutissima)有著較高的Pn和較低的Rd從而使得生長速率較快的結果相一致,這是因為高的Pn能夠使植物制造更多的光合產物,低的Rd則有利于減少對有機物的消耗,最終獲得最大的碳積累,使植物生物量積累和生長更快[19,55]。但在本研究中LAR與RGR呈極顯著的負相關關系(圖7),這與喜光植物榿木(Alnuscremastogyne)較大的LAR使其表現出較高的RGR有所差異[56],可能是因為陰生植物三七為了適應低光照環境而增大LAR,但此時光照強度過于偏低,植物光合效率較低,使得有機物的產生和積累減少,從而導致RGR降低。此外,本研究中RMR與RGR呈極顯著的正相關關系(圖7),較高RMR意味著植物將更多的生物量分配到根部,使得根部生長發育健壯,能吸取更多的水分和養分,而根部通常被認為是用于儲存有機碳的器官,增強RMR有利于有機物的積累[57],且三七是根莖類藥材,因此RMR與RGR有著較大的相關性。總之,陰生植物三七的Pn、Gs、LCP、Rd、LAR和RMR對RGR的影響較大。
植物依賴于自身的表型可塑性來調整對自然界中各種資源的獲取和消耗,以維持自身正常的生長和代謝,從而響應不同的環境條件[58]。可塑性指數可以直觀反映植物對環境變化適應能力的大小[8—10]。研究發現,陽生植物比陰生植物表現出較大的表型可塑性,且可塑性方式存在差異[59]。張常隆等研究表明,陽生植物紫莖澤蘭(Ageratinaadenophora)的葉面積、株高、冠寬、分枝數、LAR以及莖、葉生物量等指標的可塑性指數較大,這說明其主要通過調節植株形態特征以及生物量積累與分配來響應不同的環境[7]。本研究中,由可塑性分析(圖9)和主成分分析(圖10)發現,典型陰生植物三七與陽生植物有所差異,三七的形態特征可塑性相對較小,與RGR相關性不顯著,而其光合生理(Pn-max/area、Gs/area、Rd/area、LCP/area)、LAR和根生物量可塑性指數較高,在主成分上的載荷較大。這是因為三七葉片形態、光合生理及生物量分配對光強的變化是十分敏感的,尤其是光合生理(Pn-max/area、Gs/area、Rd/area、LCP/area)和LAR。而三七根部作為碳儲存的器官和經濟器官,其生物量的積累對環境的變化也較為敏感。總的來看,陰生植物主要依賴于光合生理及生物量分配的改變來響應光環境的變化。
3.4 10% LT左右的光照條件最有利于三七生長發育
三七作為一種典型的陰生植物,其對光照環境十分敏感[60],在有關三七栽培的資料中,有人認為“三分陽、七分陰”(即透光率在30%左右)的光照條件適宜三七的生長[35]。另外,早期也有研究指出三七生長適宜的透光率為40%—60%[62];而前期研究認為,從光合生理角度判斷,當透光率為9.6%—11.5%時,三七的光合生理表現較好[31—33]。本研究結果表明,三七在透光率為9.70%時,株高、莖粗和單株葉面積等形態特征明顯優于其他光照條件下的,同時各部分生物量和總生物量積累最高,高光或低光都會顯著影響其形態的生長發育和生物量的積累(圖2、圖3)。此外,與透光率9.70%相比,高光照水平下植株的Pn-max/area、Gs/area、LCP/area雖差異不顯著,但仍表現出一個較低的值;透光率低于9.7%時,隨著透光率降低,凈光合速率明顯減小。從形態特征、光合生理及生長特征綜合看來,陰生植物三七在透光率9.70%時各特征都表現優良。因此,建議在三七種植實踐中應該采取“一分陽,九分陰”(即透光率為10%左右)的光照強度管理措施。
4 結論
綜上所述,在光照強度變化的生長環境中,陰生植物三七形態可塑性較小,光合生理和生物量分配可塑性較大,且Pn-max、Gs、LCP、LAR與RGR相關性較強,表明三七光合生理特性、LAR和根生物量的變化是光照強度驅動的主要響應特征。綜合看來,9.70%左右的透光率下三七生長發育良好,因此在栽培管理中建議采取透光率為10%左右的光照環境,即“一分陽,九分陰”。對于不利的生長環境光照強度,陰生植物三七通過提升捕光能力和采取緩慢的碳獲得和碳消耗的保守策略適應低光環境;在高光條件下則會降低捕光面積、增加根部生物量,并采取快速的碳獲得和碳消耗的冒險策略。
猜你喜歡
數學小靈通·3-4年級(2024年2期)2024-05-15 02:02:28
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:36
瘋狂英語·新策略(2019年10期)2019-12-13 08:43:28
當代陜西(2019年10期)2019-06-03 10:12:04
數學小靈通·3-4年級(2017年9期)2017-10-13 08:10:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
