水分脅迫下發草(Deschampsia caespitosa)葉片脯氨酸及其代謝產物變化
2022-05-26 08:32:18羅巧玉馬永貴王彥龍拉毛葉馬玉壽
生態學報 2022年9期
羅巧玉,陳 志,馬永貴,王彥龍,拉毛葉,馬玉壽,*
1 青海師范大學生命科學學院,高原科學與可持續發展研究院,青海省青藏高原藥用動植物資源重點實驗室,西寧 810008 2 青海大學農牧學院,西寧 810008
水分是限制植物生長重要的非生物因子之一,影響植物的生存、生理代謝、生長發育乃至地理分布[1—2]。同時,水分脅迫是植物生長發育過程中經歷的最常見、最普遍的脅迫因子[3—4]。干旱、淹水、高溫、冰凍或鹽漬等都能引起水分脅迫[5—8]。水分脅迫對植物的影響主要體現在細胞活性、組織和器官功能上[9]。水分脅迫下植物體內積累大量活性氧,水分代謝平衡被打破,細胞原生質脫水、水勢下降,導致植物在形態和功能上發生重大變化[10]。為保護自身免受傷害,植物進化出有效的抗氧化系統以清除過多的活性氧[11—12]。其中脯氨酸(proline,Pro)作為一類分布廣泛的重要滲透調節物質,在防止水分脅迫對植物造成傷害中起重要作用[13]。Pro的生物合成途徑有2條,包括以谷氨酸(glutamate,Glu)為底物的Glu途徑和以鳥氨酸(ornithine,Orn)為底物的Orn途徑[12]。Glu途徑中Glu生成谷氨酸半醛(glutamic-γ-semialdehyde,GSA)是可逆反應,Δ1-吡咯琳-5-羧酸合成酶(Δ1-pyrroline-5-carboxylate synthetase,P5CS)是限速酶、Δ1-吡咯琳-5-羧酸脫氫酶(Δ1-pyrroline-5-carboxylate dehydrogenase,P5CDH)是逆反應的催化酶[7]。Orn途徑中鳥氨酸轉氨酶(ornithine aminotransferase,δ-OAT)是關鍵酶[14]。GSA和Δ1-吡咯琳-5-羧酸(Δ1-pyrroline-5-carboxylate,P5C)是Pro代謝途徑的中間產物,可以自發地相互轉化。Pro降解過程是合成過程的逆過程,Δ1-吡咯琳-5-羧酸還原酶(Δ1-pyrroline-5-carboxylate reductase,P5CR)是催化酶、脯氨酸脫氫酶(proline dehydrogenase,ProDH)是限速酶[7]。目前,關于Pro對干旱[5,15]、水澇[6,16]、鹽[15,17]、凍融[12,18]等脅迫中的響應規律已有較多報道,但不同植物對干旱和水澇脅迫的耐受、抵御機制不盡相同[12—13,16],而且有關從干旱到水澇的梯度水分脅迫下Pro積累以及Pro合成途徑中酶活性、底物及中間產物動態變化等的研究較少。
發草(Deschampsiacaespitosa)別名無芒發草、小穗發草,是禾本科(Gramineae)發草屬(Deschampsia)多年生草本植物,具有耐寒、耐旱、耐水淹、耐鹽堿、耐重金屬等優良特性[19—21]。發草適口性好、營養物質含量高,是值得大力推廣的優良牧草[22]。同時,發草莖葉柔軟、耐修剪,是良好的地被草坪植物[23]。研究表明,發草適生范圍廣,不僅能夠生長于草原等旱生環境,而且能夠生長于河灘、沼澤等濕生生境[24—26]。沼澤濕地及邊緣過渡帶具有周期性淹水和出露交替的特征,水分條件經常發生極端干旱或淹水的變化[27],發草在長期進化中已適應極端干旱或淹水交替變化的特殊土壤水分條件。目前關于發草的研究僅見于其作為藏嵩草(Kobresiaschoenoides)、青藏苔草(Carexmoorcroftii)群落的伴生種的調查、發草的分布與起源以及形態學特征[24—26],水分脅迫下發草抗逆性特征的研究尚少。因此,研究干旱、水澇兩種截然相反的水分逆境下發草Pro生理應答機制,對該物種抗旱/澇育種、植被修復選種和資源利用等具有重要意義。
本研究以發草為供試植物,利用盆栽模擬水分脅迫試驗方法,研究從干旱到水澇的梯度水分脅迫下發草Pro積累狀況及其代謝途徑中底物和關鍵酶的動態變化規律,分析發草Pro代謝對干旱和水澇脅迫的響應特點,為進一步研究發草及發草屬植物的耐受/抵御水分脅迫機制奠定基礎,也為高寒沼澤濕地旱澇“共耐性”植物的深入研究及利用發草開展退化高寒沼澤濕地植被恢復提供理論基礎。
1 材料與方法
1.1 栽培基質
土壤基質為河沙和壤土的混合物,壤土取自青海省果洛藏族自治州瑪沁縣大武鎮高寒草甸,沙和土以1∶1(體積分數)混合均勻。其理化性質為:全氮0.31%、全磷0.26 mg/g、全鉀19.58 mg/g、有機質1.45%、pH 7.63(水土比為1∶1)、電導率225.52 μS/cm(水土比為5∶1)。
1.2 供試植物
供試植物發草種子由青海大學畜牧獸醫科學院草原研究所提供,是經過多年野生栽培馴化的新品系。挑選飽滿一致、無病害種子,用2% NaClO3浸泡消毒5—10 min,用蒸餾水漂洗3—5次后備用。
1.3 實驗設計
本研究于青海師范大學城北校區(36.742°N,101.749°E)進行。該試驗點海拔2390.6 m,夏季平均氣溫16.4 ℃。2018年9月將發草種子直接播種于裝有3 kg供試土壤的底部帶孔盆缽(直徑20 mm,高25 mm)內,3—5 d出苗,待幼苗穩定后進行定苗,每盆定苗10株。期間對幼苗進行正常水分管理。冬季將植物轉移至溫室內過冬,2019年4月中下旬天氣轉暖時將植物轉移至室外露天培養。7月25日發草植株長至25 cm時進行水分脅迫處理,具體水分處理[28—29]如表1所示。采用完全隨機設計,每個處理設10個重復。干旱處理及植物正常需水量處理的盆缽底放置盆托,水澇處理的盆缽底放置水桶以免水分流出。田間持水量采用環刀法測定[30],用環刀采集具代表性原狀盆土進行土樣吸水使土壤水分達到飽和,排除重力水后烘干稱重。水分處理過程中,原地搭建遮雨棚,雨棚兩側通風,不影響溫度和濕度。遮雨棚內放置便攜式氣象儀(霍爾德HED-SQ,中國),監測實時氣象數據。水分處理期間大氣溫度和濕度狀況如圖1所示。用土壤水分傳感器(ProCheck,USA)監測土壤含水量進行水分控制[28—31],每兩天補充損失水分以控制土壤水分達到處理條件,并設置1個無植物盆土作為對照,估計土壤表面蒸發水分量。水分處理期間各處理的土壤含水量如圖2所示。每次澆水時間為18:00—19:00。澆水過程中觀察并記錄植株性狀變化。水分脅迫處理共持續28 d,分別在試驗處理前及處理后7、14、21、28 d取植物葉片。用蒸餾水漂洗后拭干表面水分,裝入凍存管經液氮速凍,置于-80 ℃冰箱保存備用。

表1 發草水分脅迫處理設計Table 1 Treatment design of D. caespitosa about water stress

圖1 水分處理期間的大氣溫度和濕度狀況Fig.1 Atmospheric temperature and humidity conditions during water treatments

圖2 水分處理期間不同水分處理下土壤含水量狀況Fig.2 Soil water content under different water treatmentsHW:重度水澇脅迫Heavy waterlogging stress;MW:中度水澇脅迫Medium waterlogging stress;LW:輕度水澇脅迫light waterlogging stress;CK:對照Control check;LD:輕度干旱脅迫Light dry stress;MD:中度干旱脅迫Medium dry stress;HD:重度干旱脅迫Heavy dry stress
1.4 測定指標與方法
Pro含量測定采用酸性茚三酮顯色法[32],酶液提取參照Lutts 等方法[33]。Glu、Orn、GSA、P5C含量及P5CDH、P5CR活性測定采用上海江萊生物科技有限公司生產的酶聯免疫分析試劑盒。P5CS活性測定采用Garci-Ríos等[34]方法:100 mmol/L Tris-HCl(pH 7.2)緩沖液(包含25 mmol/L MgCl2、75 mmol/L Glu、5 mmol/L ATP、0.4 mmol/L NADPH)中加入粗酶液啟動反應,340 nm下測定吸光值的減少量。δ-OAT活性測定采用Charest等[35]的方法:在0.2 mol/L Tris-KOH(pH 8.0)緩沖液(包含5 mmol/L Orn、10 mmol/L α-酮戊二酸、0.25 mmol/L NADH)加入粗酶液啟動反應,340 nm下測定吸光值的減少量。ProDH活性測定采用Lutts等[33]方法:0.15 mol/L Na2CO3-HCl(pH 10.3)緩沖液(包含15 mmol/L L-脯氨酸、1.5 mmol/L NAD+)中加入粗酶液啟動反應,340 nm下測定吸光值的增加量。以上指標每處理重復測定3次。
1.5 數據分析
采用SPSS 22軟件對試驗數據進行單因素方差分析(One-way ANOVA),采用LSD法在顯著水平為5%條件下進行比較,當P<0.05時,差異顯著。數據結果均用“平均值±標準誤”表示,并用利用Origin 2017軟件作圖。
2 結果與分析
2.1 水分脅迫下發草生長以及葉片Pro含量的變化
水分脅迫下發草的生長受到抑制,隨著水分脅迫時間增長,植物受到的抑制效果越明顯。干旱脅迫下發草葉片由鮮綠色變成墨綠泛白狀態,隨著干旱脅迫時間增長,葉片不斷萎蔫,直至21 d后重度干旱處理下發草死亡。水澇脅迫下發草葉片顏色變化不明顯,但是隨著水澇脅迫時間增長,葉片及枯落物腐爛發臭,28 d后重度水澇處理下發草地上部分全部腐爛。
根據不同處理對發草Pro含量的方差分析結果(表2)可知,水分處理、處理時間及水分處理和處理時間的互作對發草Pro含量的影響極顯著(P<0.01)。由圖3發現,未進行水分處理前,所有植物Pro含量均無顯著性差異。水分處理下發草Pro含量均顯著高于對照。隨著處理時間增長,Pro含量呈現增加趨勢。其中中度干旱處理下Pro含量最大,達到142.10 μg/g鮮重。除了重度水澇和重度干旱處理下發草Pro含量一直升高外,其他水分處理21 d前后Pro含量逐漸下降或者趨于穩定不再繼續升高。上述結果表明,干旱到水澇的梯度水分脅迫下發草Pro含量均顯著增加,從而參與滲透調節來抵制逆境。隨著處理時間增長,Pro含量增加,但除了重度干旱和重度水澇脅迫外,其他水分脅迫處理21 d前后Pro含量趨于穩定。
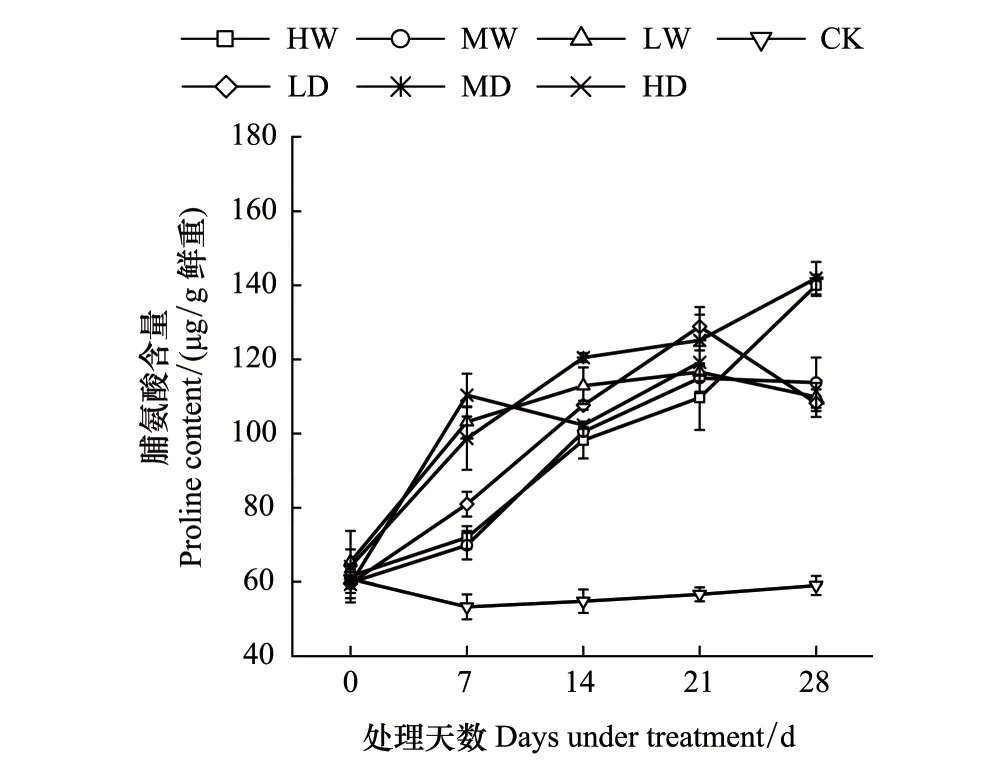
圖3 不同水分處理下發草葉片脯氨酸含量的動態變化 Fig.3 Dynamic changes of proline content in D. caespitosa leaves under different water treatments
2.2 水分脅迫下發草葉片Pro代謝底物和中間產物含量的變化
植物Pro代謝途徑包括Glu途徑和Orn途徑,Glu和Orn分別是這兩種代謝途徑中的底物。根據不同處理對發草Pro代謝途徑中底物、中間產物含量的方差分析結果(表2)可知,水分處理及水分處理和處理時間的互作對發草Glu和Orn含量的具有極顯著影響(P<0.01);處理時間對發草Glu含量也具有極顯著影響(P<0.01)。從不同水分處理下發草Pro代謝底物Glu和Orn含量的動態變化(表3)發現,未進行水分處理前,所有植物間Glu和Orn含量均無顯著性差異。重度干旱和水澇處理在7 d時Glu含量顯著下降(P<0.05),其中重度干旱處理下Glu含量為9.56 μg/g鮮重。輕度干旱處理下Glu含量無顯著變化,直到21 d時迅速降低。隨著處理時間增長,Glu含量上升,28 d時趨于穩定。其中輕度水澇處理下Glu含量最高為20.43 μg/g鮮重。28 d時重度水澇、中度水澇和輕度干旱處理下發草Glu含量顯著低于對照(P<0.05)。重度干旱處理7 d Orn含量顯著降低(P<0.05)。中度和輕度水分處理下Orn含量有升高趨勢。水澇處理后Orn含量無變化,14 d后發草Orn含量顯著降低(P<0.05)。綜上所述,干旱到水澇的梯度水分脅迫下發草Glu和Orn含量均顯著減少,Glu和Orn共同參與Pro代謝途徑以生成更多Pro。

表2 不同處理對發草葉片脯氨酸及其代謝途徑中底物、中間產物含量的方差分析結果Table 2 Results of ANOVA for the effects of water treatment and treatment time on the content of proline and its substrates and intermediates of the metabolic pathway in D. caespitosa leaves
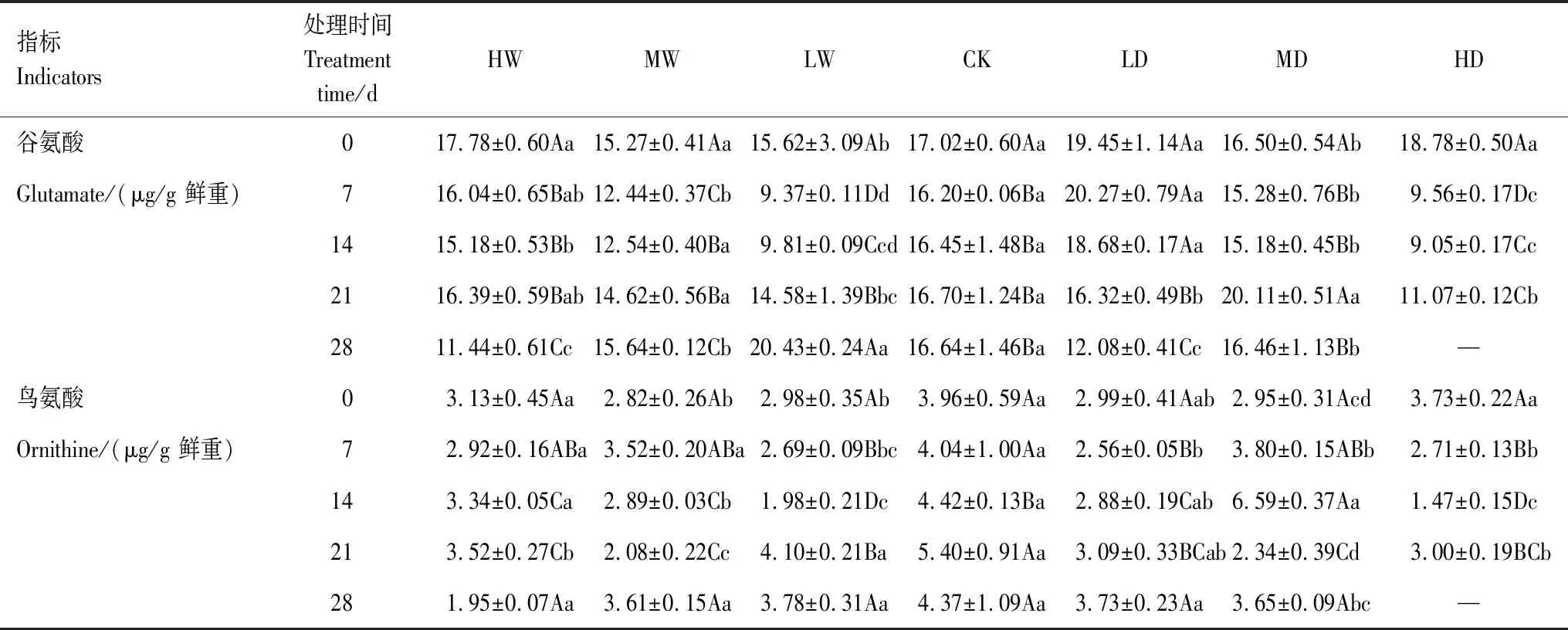
表3 不同水分處理下發草葉片脯氨酸代謝底物谷氨酸和鳥氨酸含量的動態變化Table 3 Dynamic changes of glutamate and ornithine content in D. caespitosa leaves under different water treatments
GSA和P5C是植物Pro合成途徑中的中間代謝產物。根據不同處理對發草Pro及其代謝途徑中底物、中間產物含量的方差分析結果(表2)可知,水分處理、處理時間及水分處理和處理時間的互作均對發草GSA和P5C含量具有極顯著影響(P<0.01)。從不同水分處理下發草Pro代謝中間產物GSA和P5C含量的動態變化(表4)發現,未進行水分處理前,發草葉片的GSA和P5C含量均無顯著性差異。水分處理后發草GSA和P5C的含量隨著處理時間增加,但未呈現明顯規律。重度水澇、輕度水澇、輕度干旱、中度干旱、重度干旱處理下發草GSA含量顯著升高(P<0.05)。干旱到水澇的梯度水分脅迫下發草P5C與GSA含量變化趨勢基本一致。

表4 不同水分處理下發草葉片脯氨酸代謝中間產物谷氨酸半醛和Δ1-吡咯啉-5-羧酸含量的動態變化Table 4 Dynamic changes of Glutamine hemialdehyde and Δ1-pyrroline-5-carboxylate content in D. caespitosa leaves under different water treatments
2.3 水分脅迫下發草葉片Pro代謝關鍵酶活性的變化
P5CS、P5CDH、δ-OAT、P5CR和ProDH是植物Pro合成途徑中的關鍵酶。根據不同處理對發草Pro代謝關鍵酶活性的方差分析結果(表5)可知,水分處理及水分處理和處理時間的互作均對發草葉片中影響Pro代謝的關鍵酶P5CS、P5CDH、δ-OAT、P5CR和ProDH的活性具有極顯著影響(P<0.01),處理時間對發草葉片δ-OAT的活性也具有極顯著影響(P<0.01)。
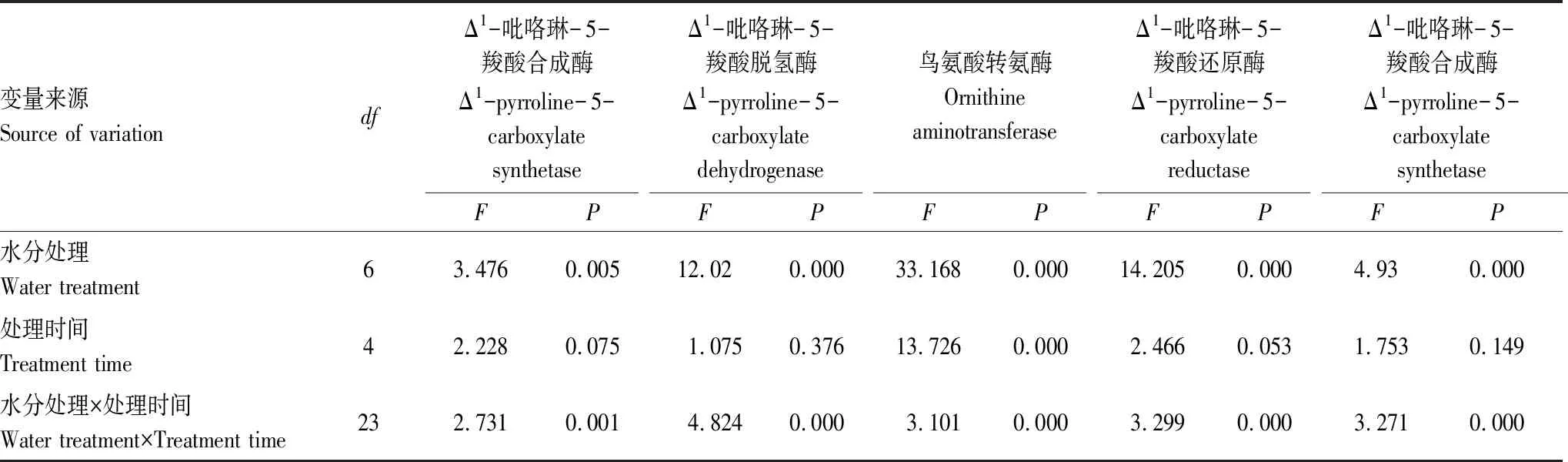
表5 不同處理對發草葉片Pro代謝關鍵酶活性的方差分析結果Table 5 Results of ANOVA for the effects of water treatment and treatment time on the key enzymes activity of Pro in D. caespitosa leaves
P5CS是Glu途徑中的限速酶,Glu在P5CS的催化作用下生成GSA,進而生成Pro。從不同水分處理下發草Pro代謝關鍵酶活性的動態變化(表6)表明,水分處理后P5CS活性呈現增強趨勢。重度水澇和中度水澇處理下不同處理時間P5CS的活性無顯著變化。輕度水澇處理下P5CS的活性于28 d時最強。輕度干旱處理P5CS的活性呈先增強后降低趨勢。處理14 d,重度干旱處理P5CS的活性顯著增強為9.49 U/g鮮重。處理21 d時降低到7.93 U/g鮮重。Glu途徑中Glu生成GSA是可逆反應,P5CDH是逆反應的催化酶。水分處理后P5CDH活性下降,變化趨勢與P5CS活性的變化相反。δ-OAT是Orn途徑中的關鍵酶,可將Orn轉化為GSA,進而合成Pro。除重度水澇外,水分處理后δ-OAT活性均呈增強趨勢。不同水分處理下δ-OAT活性增強的時間不一致,處理7 d時,輕度水澇處理和重度干旱處理下發草葉片δ-OAT的活性顯著增強(P<0.05);處理14 d時,輕度干旱處理和中度干旱處理下發草葉片δ-OAT的活性顯著增強(P<0.05)。P5CR是Pro生物合成通路中的關鍵酶,將P5C還原成Pro。水分處理前期P5CR的活性無顯著變化,但隨著處理時間增加,輕度干旱、重度干旱、輕度水澇和重度水澇處理下活性增強顯著。ProDH是Pro降解反應的限速酶,將Pro降解為P5C。隨著水分處理時間增加,發草ProDH活性顯著下降。21 d時水分脅迫下發草ProDH活性顯著低于對照植物,尤其在輕度水澇和重度水澇處理下ProDH活性降低顯著。

表6 不同水分處理下發草葉片脯氨酸代謝關鍵酶活性的動態變化Table 6 Dynamic changes of the key enzymes activity of proline in D. caespitosa leaves under different water treatments
2.4 水分脅迫下發草葉片Pro代謝中各代謝物、關鍵酶之間的相關性
對水分脅迫下發草Pro代謝途徑中Pro、Glu、Orn等10個指標進行相關性分析(表7),結果表明:發草Pro含量與Glu和Orn含量、ProDH活性具有顯著負相關(P<0.05),與GSA和P5C含量及δ-OAT活性具有極顯著正相關(P<0.01);Glu含量與GSA含量、δ-OAT活性間存在極顯著負相關(P<0.01),與P5CS和P5CR活性間存在極顯著正相關(P<0.01);Orn含量與P5C含量、P5CR活性具有極顯著正相關(P<0.01),與GSA含量具有顯著正相關(P<0.05),與P5CDH活性存在顯著負(P<0.05);GSA含量與P5C含量、δ-OAT活性間具有極顯著正相關(P<0.01),與ProDH活性具有顯著負相關(P<0.05);P5C含量與P5CR活性間存在極顯著正相關(P<0.01),與δ-OAT活性間具有顯著正相關(P<0.05);P5CS活性與P5CR活性間具有極顯著正相關(P<0.01);P5CDH活性與δ-OAT活性間具有顯著正相關(P<0.05)。

表7 發草葉片Pro代謝中各代謝物、關鍵酶之間的相關性Table 7 Correlation analysis of metabolites and key enzymes of Pro metabolism in D. caespitosa leaves
3 討論
3.1 水分脅迫對發草葉片Pro含量變化的影響
Pro是分子透性最大、極易溶于水的相容性滲透劑和抗氧化劑,在植物體內分布廣泛并以游離狀態存在。Pro的水合能力很強,在抵抗水分脅迫中發揮重要作用[5,17]。Pro的積累可提高細胞液濃度,有效保持滲透平衡,使水分進入細胞或降低水分從細胞中流出,防止細胞過度缺水而變形[7];Pro和酶相互作用可以穩定蛋白
質的結構,保持細胞膜完整性,防止膜蛋白變性[6];Pro具有保護光復合物Ⅱ的作用[36],維持逆境脅迫下植株正常的光合作用[37]。因此,逆境中植物體內Pro含量升高既可能是植物適應性的表現,也可能是植物細胞受損的表征[5,15—16]。大量研究將Pro含量高低作為衡量植物抵抗逆境能力強弱的指標[38—39]。因此,發草體內Pro含量高低可作為衡量在水分脅迫環境下發草抗逆性強弱的指標。本研究結果表明,干旱到水澇的梯度水分脅迫下,發草葉片Pro含量均顯著升高(P<0.05),說明發草植株內Pro含量的升高是主動積累的過程,通過積累Pro來緩解水分脅迫損傷。本研究結果證實植物受到干旱、水澇等水分脅迫時會通過積累Pro來增強對逆境的抵抗能力[38—39]。隨著處理時間延長,水分脅迫下發草植株內Pro含量升高,但21 d前后Pro含量不再增加甚至有下降趨勢。前期和中期,生長于水分逆境下的發草葉片內Pro含量逐漸積累,調節發草細胞質滲透壓,穩定了生物大分子的結構,提高了發草抗逆性,保證生理代謝正常進行。后期,發草逐漸對水分逆境產生適應性,體內Pro含量保持穩定。目前,其他植物與發草對水分逆境抗逆性強弱的比較研究尚無報道,但本研究中重度干旱和重度水澇脅迫下發草分別于處理21 d和28 d后死亡,表明發草對水澇和干旱具有較強的耐受性及旱澇“共耐性”特征。任青吉等[40]對高寒沼澤化草甸上的華扁穗草(Blysmussinocompressus)、草地早熟禾(Poapratensis)、發草等51種植物的葉片形態特征和光合生理進行了比較研究,發現發草的水分利用效率(water use efficiency,WUE)最高為3.76 μmolCO2/mmolH2O,而蒸騰速率(transpiration rate,Tr)最低為4 mmol H2O m-2s-1。高的水分利用效率和低的蒸騰速率保證了發草可以在水分狀況較差的土壤上正常生長。
3.2 水分脅迫對發草葉片Pro代謝中各代謝物含量及關鍵酶活性變化的影響
不同植物、甚至同一植物在不同環境條件下Pro累積的代謝途徑不相同[6,15—16]。總體來說,逆境脅迫下Pro的積累一方面是Pro合成代謝加快,另一方面是Pro氧化分解速率降低[11]。水分脅迫下,植物體內Pro代謝關鍵酶活性變化直接決定Pro的積累狀況。本研究發現,干旱到水澇梯度水分脅迫下,發草葉片內Glu途徑中底物Glu的含量和Orn途徑中底物Orn的含量均顯著下降(P<0.05),同時P5CS活性、δ-OAT活性、P5CR活性均顯著增強(P<0.05),表明Pro合成代謝中Glu途徑和Orn途徑加強。P5CS活性、δ-OAT活性、P5CR活性增強可能是水分脅迫時植物體內誘導一些具有脫氨、同分異構或氧化蛋白質中氨基酸殘酶的活性增強,使P5CS、甜菜堿醛脫氫酶(BADH)、δ-OAT、P5CR的轉錄水平上調[41]。而ProDH活性顯著降低(P<0.05),表明Pro分解代謝受到抑制,說明水分脅迫條件下發草Pro含量升高是通過合成代謝的加強和分解代謝的抑制而主動積累的結果,與Das等[6]的研究結果相似。通過Pro含量與Pro代謝中各代謝物、關鍵酶之間的相關性分析發現Pro含量與δ-OAT活性具有顯著性正相關(P<0.01),而與P5CS無顯著相關性,可能與P5CS受Pro的負反饋調節有關,當Pro含量增加時負反饋調節抑制了P5CS活性,而δ-OAT活性不受Pro的反饋調節[42]。
4 結論
干旱到水澇的梯度水分脅迫下,發草葉片通過提高其Pro含量來緩解滲透脅迫造成的傷害。發草葉片內Pro含量的升高是主動積累的過程,是Glu途徑和Orn途徑共同作用的結果。持續21 d的重度干旱處理和持續28 d的重度水澇處理時發草死亡,共同表明發草對水澇和干旱具有較強的耐受性。
猜你喜歡
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國衛生(2016年3期)2016-11-12 13:23:26
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
