我國草莓膠孢炭疽菌的多基因聯合鑒定與致病性分析
2022-05-26 02:53:02侯圣凡華戰迎劉峻杰董振飛馮澤坤閆佳琪王紅清
中國農業大學學報 2022年4期
侯圣凡 華戰迎 劉峻杰 董振飛 馮澤坤 閆佳琪 王紅清
(中國農業大學 園藝學院,北京 100193)
草莓(Fragaria×ananassaDuch)是一種重要的經濟作物,因其種植周期短、結果早且經濟效益高,所以在地方特色經濟中占有重要的地位[1]。我國草莓主產區有山東、安徽、遼寧、河北和江蘇等地。近年來,山西和云南等地的草莓產業也得到迅速發展。我國草莓主要是依靠設施栽培,由于多年連續種植,導致草莓根部病害近年來頻繁發生,其中以真菌性病害對草莓的影響最為顯著,對我國草莓產業的健康發展造成了巨大的危害[2]。
炭疽病是一種寄主廣泛且種類繁多的常見植物病害,而且此類真菌存在的地域范圍廣,可侵染多種農作物。1931年,Brooks等[3]最先報道了侵染草莓匍匐莖和葉柄的病原菌為草莓炭疽菌,早期草莓炭疽病只是零星分布,后蔓延至整個佛羅里達州,經鑒定引起炭疽病的病原為草莓炭疽菌(Colletotrichumfragariae)。1965年,澳大利亞等地公布了草莓炭疽菌和尖孢炭疽菌(C.acutatum)的存在[4]。1978年Mass等[5]根據對馬里蘭的草莓病害研究調查認為草莓炭疽病病原主要為膠孢炭疽菌(C.gloeosporioides)、草莓炭疽菌和尖孢炭疽菌。近年來,炭疽病在草莓生長中危害日趨嚴重,已經成為草莓苗期甚至整個生長期的主要病害。設施環境下,草莓炭疽病大多不在地上部表現癥狀,而是在定植后的緩苗期危害根頸,引起根部腐爛,導致植株萎蔫和枯死。其致死率一般在10%~20%,有的可達到80%以上[6-7]。葉正文等[8]于1997年首次在我國報道草莓炭疽病并初步鑒定草莓炭疽病病原菌有尖孢炭疽菌和草莓炭疽菌2種,且以尖孢炭疽菌為主。2007年張海英等[9]經過鑒定發現引起撫順草莓炭疽病的病原菌為膠孢炭疽菌和草莓炭疽菌。
尖孢炭疽菌與草莓炭疽菌通常對草莓地上部葉片及果實產生危害,地上部癥狀容易發現,可以及時防治。但膠孢炭疽菌引起的根部病害難以及時發現與控制,對草莓危害較重。本研究通過對草莓炭疽根腐病病原菌膠孢炭疽菌分離純化發現,菌株在PDA平板上呈現不同的形態特征,因此對膠孢炭疽菌展開進一步的分析研究。膠孢炭疽菌分布地理范圍廣泛,寄主眾多,具有復合種和種群集合的特征[10],傳統的使用單一ITS序列鑒定真菌有時會出現混淆錯誤,結果不可靠。Crouch等[11]通過對炭疽菌ITS序列比對分析發現錯誤率可高達86%。因此本研究將通過菌落形態鑒定、分離純化與分子鑒定、多基因序列(ITS-ACT-GAPDH-CAL-TUB2-CHS)聯合鑒定以及致病性鑒定的方法,對我國草莓主產區的草莓炭疽病病原菌膠孢炭疽菌復合種進行系統發育與致病性分析,以期為病害防控提供科學依據。
1 材料與方法
1.1 材料
本研究于2017—2019年進行。樣品采自草莓主產區山東臨沂、山東聊城、河北懷來、遼寧東港、江蘇徐州、安徽合肥、北京昌平、北京順義、四川成都、甘肅定西、云南玉溪和山西晉城12個地區,采集草莓根腐病樣品480株。經過對草莓病樣的歸類整理得到109株菌株進行后續研究(表1)。供試草莓品種有‘紅顏’‘哈尼’‘寧玉’‘香野’‘妙香’‘甜查理’。

表1 草莓根腐病病樣菌株采集信息Table 1 Collection information of strawberry root rot disease strains

表1(續)

表1(續)
1.2 試驗方法
1.2.1病原菌分離與純化
取草莓根頸部病健交界處的組織(0.5~1 cm2左右),用75%酒精滅菌30 s,無菌水洗滌3次;之后用2%的次氯酸鈉消毒6~8 min,無菌水洗滌3~4次;晾干置于PDA培養基,在25 ℃的恒溫恒濕培養箱生長5~7 d。從菌落邊緣處挑取菌落菌絲置于PDA液體培養基中,在25 ℃搖床中培養1 d。采用單孢劃線分離法將單孢菌轉移到PDA平板上用于鑒定和后續保存。
1.2.2DNA提取及多基因序列擴增
根據BufferA和BufferB溶液進行DNA的快速提取[12]。挑取少量菌絲放于裝有50 μL BufferA溶液的PCR管中,之后置于PCR儀內,95 ℃裂解10 min;再向每個PCR管內加入50 μL Buffer B溶液,震蕩混勻,12 000 r/min離心20 s,取上清液50 μL于新PCR管中,即獲得所需DNA并測定其濃度。基于多基因序列各引物(表2)進行PCR擴增,反應體系(25 μL)包括:PCR mix 12.5 μL、上下游引物(10 μmol)各1 μL、DNA模板1 μL,加dd H2O 9.5 μL。PCR反應程序:94 ℃預變性3 min;94 ℃變性30 s,55 ℃退火45 s,72 ℃延伸1 min,30個循環;72 ℃延伸3 min。放于4 ℃保存。擴增產物使用1.5%瓊脂糖凝膠電泳進行PCR檢測,PCR產物送擎科公司測序。把測序成功的目的序列登錄到NCBI數據庫進行比對,下載同源性序列(表3)。用MEGA 7.0軟件進行多重比對并修正[13],利用鄰近法構建系統發育樹[14]。
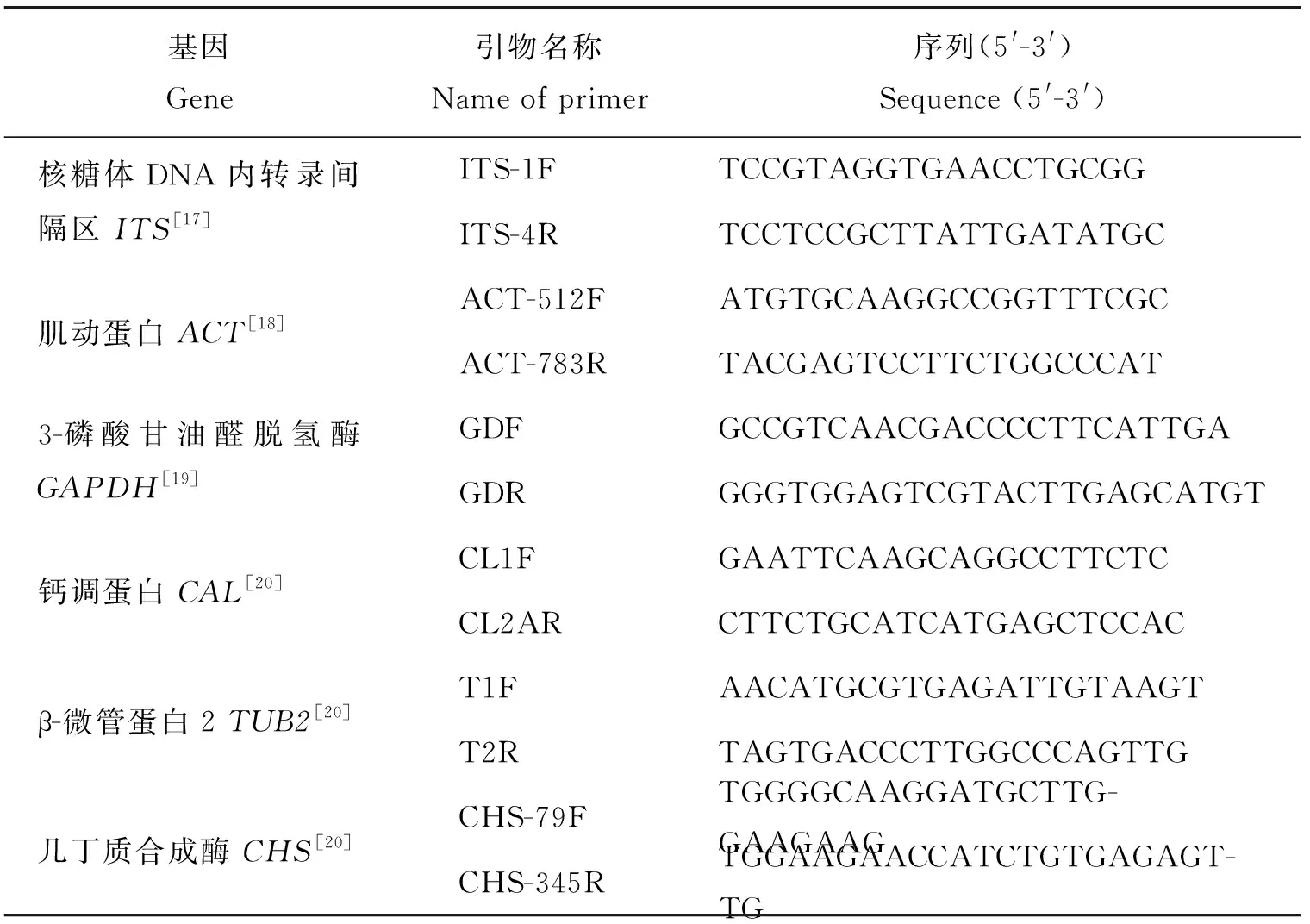
表2 各引物及PCR循環條件Table 2 Primers and PCR cycle conditions

1.2.3真菌致病性測定
選取生長狀況一致的草莓苗‘紅顏’進行噴霧法接種。將不同菌株的分生孢子制成1.0×106個/mL孢子懸浮液,均勻噴灑于草莓葉片上(以有水滴流下為度),以噴灑無菌水為對照,每處理噴灑5株,重復3次。接種后草莓植株置于培養箱(25 ℃)中黑暗覆膜保濕培養24 h。之后揭膜,在25 ℃,光周期為12 h/12 h下培養。
參考王豐等[15]方法,并稍加改動,對接種草莓苗病情分級與菌株致病性進行等級劃分。0級:無病;1級:葉片枯萎面積占葉面積5%以下;3級:葉片枯萎面積占葉面積5.1%~15%;5級:葉片枯萎面積占葉面積15.1%~30%;7級:葉片枯萎面積占葉面積30.1%~50%;9級:葉片枯萎面積占葉面積50%以上或葉片枯死。發病率(DR)=發病株數/調查株數×100%,病情指數(DI)=[(各級發病株數×病級數)/(最高發病級數×調查株數)]×100。利用SPSS 26.0軟件進行致病性分析,采用最小顯著差數法(LSD)進行差異顯著性檢測,并完成柯赫氏法則驗證[16]。
2 結果與分析
2.1 草莓炭疽根腐病田間癥狀
草莓定植初期,炭疽根腐病病原菌通過侵染草莓根部,分泌相應毒素。其孢子在維管系統中定植,破壞水分傳導的木質部導管,導致植物萎蔫、干枯直至死亡(圖1(a));切開病株短縮莖會發現根頸褐變、腐爛(圖1(b))。

(a)感病植株萎蔫癥狀;(b)感病植株根頸橫切面癥狀。(a) Symptoms of wilting of infected plants; (b) Transverse section of the Crown of infected plants.圖1 草莓炭疽根腐病田間癥狀Fig.1 Field symptoms of strawberry anthracnose crown rot
2.2 草莓炭疽根腐病病原菌鑒定
2.2.1病原菌形態觀察
暹羅炭疽菌菌株:在培養過程中菌落正面呈現白色,后期白色變暗,略微出現灰色;菌絲稀疏呈絮狀,后期會產生孢子泥和分生孢子盤;菌落背面中間區域為黑褐色,從中間到邊緣顏色逐漸變淡(圖2(a))。分生孢子柱狀、兩端鈍圓,尺寸為(11.73~13.99) μm×(3.00~5.02) μm(圖2(b))。隱秘炭疽菌菌株:菌落正面呈現乳白色,菌絲較為致密;菌落背面呈白色,其中間區域出現較小的褐色圓圈(圖3(a))。分生孢子柱狀、兩端鈍圓,大小較為均一,尺寸為(16.56~17.90) μm×(4.32~5.50) μm(圖3(b))。果生刺盤孢菌菌株:初始菌落正面邊緣為白色,中間略微顯現灰色,生長到第8天時,菌落正面出現大面積灰褐色,菌絲稀疏,氣生菌絲較為茂盛,菌落背面出現大面積灰黑色(圖4(a))。分生孢子柱狀、兩端鈍圓,尺寸為(9.96~13.03) μm×(3.30~3.81) μm(圖4(b))。暹羅炭疽菌附著胞(圖2(c))、隱秘炭疽菌附著胞(圖3(c))和果生刺盤孢菌附著胞(圖4(c))分別由是相應的3種分生孢子在水中萌發芽管后形成的梨形和卵形等暗褐色結構,其形態穩定且大小不均一。其中暹羅炭疽菌和隱秘炭疽菌菌株中有一定數量的分生孢子會產生多個附著胞,隱秘炭疽菌的附著胞芽管偏短。

(a)菌落形態特征;(b)孢子形態特征;(c)附著胞形態特征。(a) Morphological characteristics of colony; (b) Morphological characteristics of conidia; (c) Morphological characteristics appressorium.圖2 暹羅炭疽菌形態特征Fig.2 Morphological characteristics of C.siamense

(a)菌落形態特征;(b)孢子形態特征;(c)附著胞形態特征。(a) Morphological characteristics of colony; (b) Morphological characteristics of conidia; (c) Morphological characteristics appressorium.圖3 隱秘炭疽菌形態特征Fig.3 Morphological characteristics of C.aenigma

(a)菌落形態特征;(b)孢子形態特征;(c)附著胞形態特征。(a) Morphological characteristics of colony; (b) Morphological characteristics of conidia; (c) Morphological characteristics appressorium.圖4 果生刺盤孢菌形態特征Fig.4 Morphological characteristics of C.fructicola
2.2.2病原菌多基因序列聯合鑒定
提取真菌基因組DNA后,采用真菌ITS1和ITS4通用引物序列,成功從不同菌株DNA樣品中獲得DNA擴增產物。以rDNA-ITS序列構建系統發育樹,結果表明,分離純化的109個菌株中有30個菌株歸類于膠孢炭疽菌復合種。30個膠孢炭疽菌復合種可根據不同菌株形態特征分為3類。從3類菌株中各選1株代表菌株SCD26、LND07和SCD15進行多基因序列鑒定,構建膠孢炭疽菌的多基因聯合系統發育樹(圖5)。結果發現膠孢炭疽菌代表菌株聚類為3個分支,其中SCD26病原菌聚類于暹羅炭疽菌;LND07病原菌聚類于隱秘炭疽菌;SCD15病原菌聚類于果生刺盤孢菌。

SCD15、SCD26和LND07為膠孢炭疽菌復合種代表菌株。SCD15, SCD26 and LND07 are representative strains of C.gloeosporioides complex.圖5 膠孢炭疽菌復合種多基因聯合系統發育樹Fig.5 Phylogenetic tree of the C.gloeosporioides complex based on multi-gene combination
2.3 草莓炭疽根腐病病原菌致病性試驗
通過對代表性菌株隱秘炭疽菌、果生刺盤孢菌和暹羅炭疽菌進行接種實驗,發現在接種隱秘炭疽菌第9天、接種果生刺盤孢菌和暹羅炭疽菌第12和13天時草莓葉片開始發黃萎蔫,3種炭疽菌引起的病害程度差異顯著。其中隱秘炭疽菌病情指數與致死率分別為82.2和66.7%,顯著高于果生刺盤孢菌和暹羅炭疽菌。隱秘炭疽菌發病較快,引起病害較重。本研究采用葉片噴霧法接種病原菌,但在感染病原菌早期,草莓葉片并沒有出現病斑,只出現葉片發黃萎蔫直到枯死,切開草莓短縮莖,其橫截面出現大面積壞死褐變。隱秘炭疽菌感染的草莓,從短縮莖一側向髓感染(圖7(c)),而果生刺盤孢菌(圖8(c))和暹羅炭疽菌(圖6(c))感染的草莓,短縮莖髓處優先發病。按柯赫氏法則,將上述感病草莓進行組織分離,病原菌純化,鑒定其病原菌為暹羅炭疽菌、隱秘炭疽菌與果生刺盤孢菌,與最初各接種的病原菌相同,即完成了柯赫氏法則。

(a)接種第1天;(b)接種第18天;(c)患病植株根頸橫截面。(a) Day 1 of inoculation; (b) Day 18 of inoculation; (c) Cross section of rhizome of diseased plants.圖6 暹羅炭疽菌致病性測定Fig.6 Determination of pathogenicity of C.siamense

(a)接種第1天;(b)接種第18天;(c)患病植株根頸橫截面。(a) Day 1 of inoculation; (b) Day 18 of inoculation; (c) Cross section of rhizome of diseased plants.圖7 隱秘炭疽菌致病性測定Fig.7 Determination of pathogenicity of C.aenigma

(a)接種第1天;(b)接種第18天;(c)患病植株根頸橫截面。(a) Day 1 of inoculation; (b) Day 18 of inoculation; (c) Cross section of rhizome of diseased plants.圖8 果生刺盤孢炭疽菌致病性測定Fig.8 Determination of pathogenicity of C.fructicola
3 討論與結論
3.1 多基因聯合鑒定的必要性
草莓整個生育期都可發生炭疽根腐病。在草莓育苗期和定植初期,發病率可高達90%以上,給健康草莓苗的繁育造成巨大破壞[21]。草莓炭疽病的鑒定對該病害的防治有重要的參考價值。由于病原菌菌落形態特征穩定性差且易受培養條件影響,單獨依靠形態特征進行系統分析是不可靠的[22]。隨著分子生物學的發展,基因序列鑒定成為一種常用的手段。目前草莓炭疽病的分類依據主要結合于前人的分類系統,以分生孢子和附著胞的形態特征、大小、純培養特征以及ITS基因序列進行構建系統發育樹來確定其歸屬。但是僅用1種基因的測序結果,一些親緣關系較近的種仍不能進行有效區分,所以需要使用多基因序列進行物種進化的研究[23]。目前用于炭疽病鑒定分類的常用基因有ITS(核糖體DNA內轉錄間隔區)、TUB2(β-微管蛋白2)、ACT(肌動蛋白)、GAPDH(3-磷酸甘油醛脫氫酶)、CHS(幾丁質合成酶)和CAL(鈣調蛋白)等。Weir等[20]通過多基因聯合建樹的方法將膠孢炭疽菌復合種劃分為22個種和1個亞種;宋麗麗等[24]利用多基因聯合建樹法對分離自上海和安徽的6個草莓炭疽菌菌株進行分子鑒定,發現這6個炭疽菌菌株均為果生刺盤孢菌;韓永超等[25]通過多基因聯合分析明確了引起武漢地區草莓根頸腐病的病原為膠孢炭疽菌復合種內的暹羅炭疽菌;張方博[26]通過對草莓根腐病研究也發現,暹羅炭疽菌是草莓根腐病病原之一;Zhang等[27]對草莓炭疽病進行多基因聯合鑒定發現隱秘炭疽菌也屬于膠孢炭疽菌復合種。本研究發現草莓膠孢炭疽復合種目前包含3個小種即暹羅炭疽菌、隱秘炭疽菌和果生刺盤孢菌,與前人研究結果一致。
3.2 草莓炭疽根腐病優勢菌種分析
同一寄主的炭疽病病原菌菌株之間,致病力存在差異。馮樂樂等[23]對浙江省鮮食大豆炭疽病病原平頭炭疽菌進行分離鑒定,發現分離出的9株病原菌株雖同屬于平頭炭疽菌,但致病力卻不同。郭艷春等[28]認為不同地區引起黃麻炭疽病的病原菌致病能力也不同。草莓膠孢炭疽菌復合種下的暹羅炭疽菌、隱秘炭疽菌和果生刺盤孢菌可在不同程度上引起草莓根部腐爛、地上部萎蔫和干枯等癥狀。Chen等[29]認為果生刺盤孢菌為浙江地區草莓炭疽菌的優勢菌株,其致病力是膠孢炭疽菌復合種中最強的;Zhang等[27]研究結果表明在中國東部草莓栽培區中暹羅炭疽菌才是草莓炭疽病的優勢菌株,致病力最強。本研究發現隱秘炭疽菌與果生刺盤孢菌和暹羅炭疽菌相比致病力最強。不同地區氣候、土壤條件、栽培品種以及栽培措施不同可能造成草莓炭疽根腐病病原菌優勢種不同,具體是由于何種原因導致優勢菌株不同,還有待進一步研究。
本研究通過廣泛采集全國不同草莓主產區根部腐爛樣品發現,有27.5%的樣品是由膠孢炭疽菌復合種引起。膠孢炭疽菌復合種可分為3種生理小種,分別為隱秘炭疽菌、果生刺盤孢菌和暹羅炭疽菌,其中隱秘炭疽菌發病較快,引起的病癥較為嚴重。本研究還分離出43.4%的尖孢鐮刀菌專化型(Fusariumoxysporumf. sp.fragariae)、11.1%的擬盤多毛孢(Neopestalotiopsisclavispora)以及其他病原菌,這些病原菌都可以加重草莓根腐病的發生。此外,關于草莓炭疽根腐病病原菌如何與其他病原菌復合侵染引起草莓病害也需進一步研究。
