低溫脅迫下甘藍型冬油菜BnYUCCA8基因差異表達及內源生長素含量變化
2022-05-24 07:29:14呼芳娣劉麗君牛早霞孫柏林楊雯昕蒲媛媛武軍艷李學才沈金雄孫萬倉
干旱地區農業研究 2022年3期
呼芳娣,劉麗君,馬 驪,牛早霞,孫柏林,徐 佳,楊雯昕,劉 博,蒲媛媛,武軍艷,方 彥,李學才,沈金雄,孫萬倉
(1.甘肅農業大學農學院,甘肅省油菜工程技術研究中心,省部共建干旱生境作物學國家重點實驗室(甘肅農業大學),甘肅省作物遺傳改良與種質創新重點實驗室,甘肅 蘭州 730070;2.華中農業大學作物遺傳改良國家重點實驗室/國家油菜工程技術研究中心,湖北 武漢 430070)
低溫是影響植物生長發育的主要環境因子,對植物的生長發育及地理分布起決定性作用[1]。甘藍型冬油菜是我國重要的油料作物,與白菜型冬油菜相比,具有產量好、品質優的特性,但由于其抗寒性較弱,嚴重影響了甘藍型冬油菜在我國北方地區的種植[2]。
下胚軸是連接兩個子葉和初生根(胚根)的幼莖[3]。植物下胚軸的伸長是植物對環境脅迫的重要適應機制[4],其伸長的程度主要受溫度、光照等環境因子和內源激素的雙重調控[5-6]。甘藍型冬油菜下胚軸長度與其抗寒性強弱密切相關,下胚軸越短,其抗寒性越強。相關研究表明,抗寒性強的品種將有機物運輸至地下部,給予越冬期植物抵御低溫的能量保障,而抗寒性弱的品種將光合產物運輸至地上部,易造成地上部的徒長,導致下胚軸伸長,降低其抗寒能力[7]。金姣姣[8]通過對不同抗寒性甘藍型冬油菜品種的下胚軸長度觀察發現,在甘藍型冬油菜的幼苗時期,抗寒性不同品種的下胚軸伸長長度不同,強抗寒品種下胚軸伸長較短,弱抗寒品種伸長較長。
生長素是植物發育的主要調節因子,參與協調植物生長的幾乎所有方面[9]。低溫脅迫下,植物體內激素水平會發生變化進而調控生理代謝活動,激活抗寒基因表達[10],增強植物抗寒性[11]。色氨酸轉氨酶(TAA)/YUCCA(YUC)途徑是生長素生物合成的主要途徑[12],YUCCA家族基因編碼的含黃素單加氧酶是該生物合成途徑的限速酶,能夠催化以色氨酸為前體的IPA轉化為IAA[13]。擬南芥藍光受體ZTL正向調節地上部分生長素合成的IPA途徑中YUCCA8基因的表達,從而促進下胚軸伸長。相對于野生型,擬南芥ztl突變體下胚軸較短,YUCCA8表達水平降低[14],表明YUCCA8基因在調控擬南芥下胚軸長短方面發揮重要作用。高溫誘導下,擬南芥生長素生物合成基因YUCCA8被光信號途徑關鍵轉錄因子PIF4激活從而調節高溫引起的下胚軸伸長[15]。且yuc8突變可以在很大程度上抑制 PIF4 過表達植物的長下胚軸的表型,也可以減少高溫誘導下下胚軸伸長[16]。雖然YUCCA8基因在生長素及下胚軸調控方面研究較多,但主要集中于高溫誘導下, 且YUCCA8通過調控下胚軸伸長適應低溫脅迫方面的研究鮮有報道。
蒲媛媛[2]將甘藍型油菜‘16VHNTS158’種植于蘭州上川(36°03′N、海拔2150 m),地膜覆蓋以后發現其越冬率為10.6%,‘16VHNPZ269’為11.7%,‘天油 2288’為0%,其半致死溫度和春化率見表1,由表1可知‘16VHNTS158’和‘天油 2288’抗寒性具有差異。本文從甘藍型冬油菜中克隆獲得BnYUCCA8基因,對其進行了生物信息學分析,并分析了其在不同組織及低溫脅迫下表達量的變化。檢測了低溫脅迫下根和葉中內源 IAA 含量的變化。對表達量、內源 IAA 含量和下胚軸長度進行了相關性分析,以期為探究甘藍型冬油菜抗寒性與下胚軸長度之間的分子機制提供理論依據。

表1 試驗材料抗寒特性及來源
1 材料和方法
1.1 材料
于2020年9月17日,將甘藍型油菜品系‘16VHNTS158’、‘16VHNPZ269’和‘天油 2288’播種于甘肅農業大學百草園試驗基地,待長至五葉期,即同年11月12日,選取各材料中完整性好的單株,取第5片真葉,3個生物學重復,于-80℃冰箱備用,用于基因克隆。待其越冬抽薹后,即2021年4月23日,取成熟根、莖、葉、下胚軸、花、花蕾、角果于-80℃冰箱備用,每個組織部位3個生物學重復,用于組織表達分析。2021年3月11日,將‘16VHNTS158’和‘天油2288’進行盆栽,待油菜長至5~6葉期時,各取3盆取其植株的根、莖、葉、下胚軸,液氮冷凍后放入-80℃冰箱備用,用于組織表達分析,將剩余盆栽材料全部置于22℃培養箱處理48 h后,分別放置于常溫(22℃)、4℃、0℃、-4℃培養箱,每隔0、3、6、24 h取樣,各處理條件下分別取葉和根的3個生物學重復,用于低溫脅迫下熒光定量PCR及內源IAA含量的測定。
1.2 RNA提取及反轉錄
根據北京天根生物公司RNAprep Pure Plant Kit 植物總RNA提取試劑盒提取試驗材料各組織總RNA,瓊脂糖凝膠電泳檢測RNA條帶,按PrimeScript RTrea-gent Kit with gDNA Eraser 反轉錄試劑盒(大連TaKaRa公司)進行反轉錄,得到單鏈cDNA,置于-20℃冰箱保存備用。
1.3 YUCCA8基因的克隆
根據NCBI甘藍型油菜數據庫中所預測的YUCCA8(ID:106359824)基因序列,利用Primer Premier 5.0軟件設計基因克隆引物,YUCCA8-F:5′-ATGGAGAGTATGTTTCGTTTGATG-3′,YUCCA8-R:5′-TTAGAATTGTTGGGAGATACATCTTCTAT-3′;以葉片cDNA為模板進行PCR擴增,擴增程序為:94℃預變性5 min;94℃變性30 s,53.6℃退火30 s,72℃延伸60 s,循環35次;72℃終延伸10 min;4℃保存。產物經1%瓊脂糖凝膠電泳,采用北京天根的膠回收試劑盒進行純化回收,回收產物與pMD19-T載體連接,轉化大腸桿菌Trans1-T1感受態細胞,結合藍白斑篩選和菌落PCR鑒定結果,挑選出陽性單菌落送上海生工生物工程公司測序。
1.4 YUCCA8基因的生物信息學分析
利用DNAMAN軟件進行克隆基因的多重序列比對和氨基酸同源性分析;Expasy在線分析蛋白質的理化性質;SignalP-5.0 Server預測蛋白信號肽;SOPMA在線預測蛋白的二級結構;SWISS-MODEL在線預測蛋白三級結構;MEGA7.0軟件分析系統進化;TMHMM Server v.2.0在線預測跨膜結構;Wolf-psort在線預測亞細胞定位;NCBI在線工具Conserved Domain Search Service進行保守結構域預測分析。
1.5 實時熒光定量PCR
實時熒光定量PCR(RT-qPCR)反應體系為20 μL:SYBR Premix Ex Taq TM Ⅱ 10 μL、正向引物/反向引物各0.8 μL、cDNA 2.0 μL、ddH2O 6.4 μL。內參基因為Actin[17]。采用2-ΔΔCT對基因的相對表達量進行計算,并用Duncan法進行差異顯著性分析。
1.6 IAA提取及含量測定
使用美國Waters Arc型高效液相色譜儀進行測定。色譜條件為:色譜柱:Symmetry C18色譜柱(4.6×250 mm、5 μm);流動相:甲醇+0.1%磷酸(百分數為體積比);流速:1.0 mL·min-1;進樣量:10 μL;檢測波長:254 nm;柱溫:30℃。
激素提取方法:參照趙楊靜等[18]的方法,略有改良。取油菜根部0.1 g,放入研缽,液氮中研磨成粉末加入6 ml提取液A(正丙醇∶水∶HCl=2∶1∶0.002,體積比)。4℃搖床100 r·min-1震動30 min,加入3 mL提取液B(二氯甲烷),4℃搖床100 r·min-1震動30 min;取出后4℃ 13 000 r·min-1離心5 min,此時溶液分層,用巴氏滴管吸取有機層溶液,氮氣吹干,加入50%甲醇水溶液1 ml;過0.22 μm有機針孔濾膜,裝入樣品瓶中置于4℃冰箱待測。
2 結果分析
2.1 BnYUCCA8基因的cDNA克隆
采用同源克隆技術,以甘藍型冬油菜cDNA為模板,利用擴增特異性引物,對目的基因進行PCR擴增,得到與預期大小相符的條帶(圖1),將目的片段純化回收,回收產物連接轉化后測序,得到全長片段為1281bp的CDS序列,與預期結果相吻合。命名為BnYUCCA8。

M: DNA marker;1: ‘16VHNTS158’;2: ‘16VHNPZ269’;3: ‘天油 2288’ ‘Tianyou 2288’
2.2 BnYUCCA8基因生物信息學分析
對從‘16VHNPZ269’中克隆得到的BnYUCCA8基因序列進行生物信息學分析表明,基因的起始密碼子ATG,終止密碼子TAA,編碼含426aa的蛋白質(圖2),相對分子質量約47.78 kD,理論等電點為9.02。基因由20種氨基酸組成,其中Val、Leu、Gly 3種氨基酸所占比例最高,分別為9.1%、8.7%、7.5%,負電荷殘基總數(Asp+Glu)為47,正電荷殘基總數(Arg+Lys)為57;分子式為C2134H3379N585O615S22,總親水性平均系數為-0.237,蛋白質不穩定指數為49.54,是一個不穩定的疏水性蛋白(圖3A),屬于CzcO超家族(圖3D)。跨膜結構預測結構表明BnYUCCA8蛋白存在一個跨膜結構(圖3F),亞細胞定位預測主要位于細胞質和內質網上,不含信號肽,推測其為非分泌蛋白(圖3E)。

注:(A) 圖中黃色方框為內含子區域。(B) 圖中上游起始密碼子ATG用方框表示;*為終止密碼子TAA的位置。
YUCCA8蛋白質二級結構顯示,無規則卷曲(Randomcoil)含量最豐富,為41.45%,其次是33.02%的α-螺旋(α-Helix),延伸鏈(Extended strand)和β-轉角(β-turn)含量最少,分別為17.56%和7.96%(圖3C)。以6sek.1為模板,對BnYUCCA8基因編碼蛋白的三級結構進行模型構建,模型序列一致性達到23.28%(圖3B)。
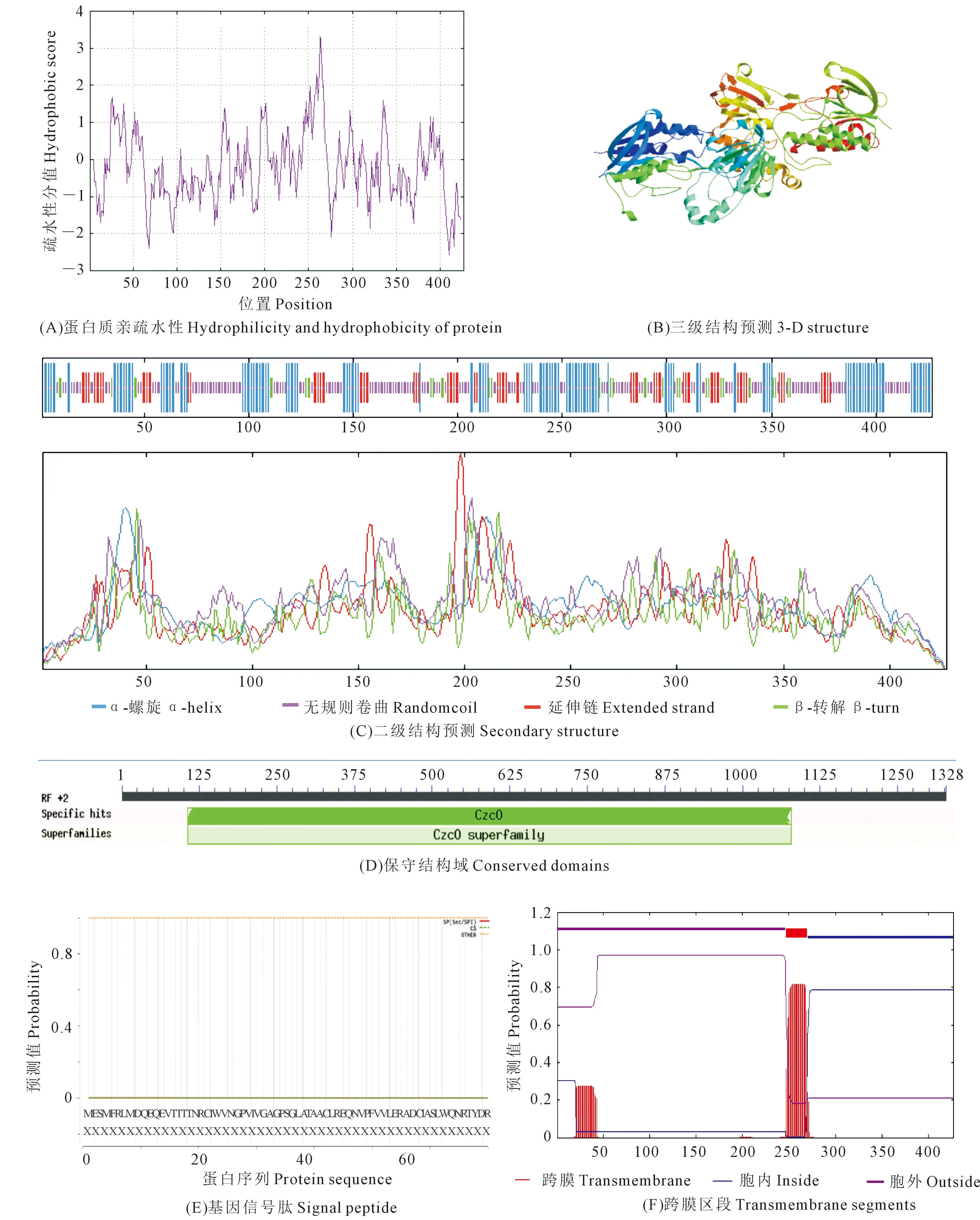
圖3 甘藍型冬油菜‘16VHNNPZ269’ BnYUCCA8蛋白質特性分析
甘藍型冬油菜‘16VHNPZ269’中BnYUCCA8基因對應的氨基酸序列與甘藍型油菜數據庫中YUCCA8基因對應的氨基酸序列相似性為98.13%、與擬南芥AtYUCCA8基因對應的氨基酸序列相似度為88.11%、與‘16VHNTS158’和‘天油2288’對應序列相似性分別為98.59%和89.72%,‘16VHNTS158’與‘天油2288’對應序列相似性為89.72%(圖4)。

圖4 不同材料氨基酸序列多重比對
2.3 BnYUCCA8的序列進化分析
利用MEGA 7.0軟件將克隆獲得的甘藍型油菜YUCCA8蛋白的氨基酸序列與從NCBI下載的11種植物的YUCCA8蛋白序列構建系統進化樹(圖5),結果表明,甘藍型油菜該蛋白與薺菜同源性最高,其次為擬南芥,與白芥和薺藍的親緣關系最遠。

圖5 不同植物YUCCA8蛋白的進化樹分析
2.4 甘藍型冬油菜BnYUCCA8基因的組織部位表達分析
組織表達分析結果顯示(圖6),BnYUCCA8基因在所有組織部位均有表達,且其表達量表現出組織部位差異和生長時期差異。BnYUCCA8基因在下胚軸伸長較短的‘16VHNTS158’的苗期階段(圖6A),葉和根中表達量最高,莖和下胚軸中表達量次之;花期階段,根和葉中表達量最高,其次為莖、下胚軸、花蕾,花和角果中表達量相對較低。下胚軸伸長較長的‘天油 2288’與下胚軸伸長較短的‘16VHNTS158’具有相同的表達模式(圖6B),即在苗期和花期兩個時期都表現出根和葉中的表達量高,其次依次為莖、下胚軸、花蕾、花、角果。

注:不同字母表示BnYUCCA8基因相對表達量分別在苗期和花期的不同組織部位間差異顯著(P<0.05)
兩個材料同一組織部位比較結果顯示,除花期莖中‘16VHNTS158’基因表達量高于‘天油 2288’,其他組織部位基因表達量皆為下胚軸較長的‘天油 2288’高于下胚軸較短的‘16VHNTS158’。在油菜苗期和花期的生育進程中,BnYUCCA8基因表達量表現為苗期高于花期。
2.5 低溫脅迫下BnYUCCA8基因在根和葉中的表達量變化
低溫脅迫下BnYUCCA8基因表達量分析結果見圖7,由圖7A可得,在‘16VHNTS158’的根中,隨著溫度降低,BnYUCCA8基因表達量總體呈降低趨勢,0℃較4℃時基因表達量略有升高,且隨著低溫脅迫處理時間的延長其基因表達量降低,低溫脅迫處理24 h時基因表達量在不同溫度梯度處理下的變化幅度較常溫(22℃)更為明顯,在4℃、0℃、-4℃的降幅分別為43%、17%、87%,且由圖7C可得,‘天油2288’根中,4℃、0℃、-4℃的降幅分別為45%、20%、54%。相同處理溫度及時間下,‘天油2288’的基因表達量高于‘16VHNTS158’。
對葉中BnYUCCA8基因表達量分析表明,在相同處理時間下,隨溫度降低其基因表達量變化模式與根中一致,處理時間越長,低溫脅迫下表達量變化趨勢越大,表達量越低。由圖7B可得,處理24 h時,‘16VHNTS158’葉中,4℃、0℃、-4℃下的表達量較常溫(22℃)分別降低了29%、28%、73%,由圖7D可得,‘天油2288’分別降低了43%、28%、64%。相同處理溫度及時間下,‘天油2288’較‘16VHNTS158’BnYUCCA8基因表達量高,由此表明BnYUCCA8基因可能通過調控下胚軸伸長進而參與甘藍型冬油菜抗寒性的調控。

注:不同小寫字母表示BnYUCCA8基因相對表達量在相同處理時間不同溫度間差異顯著(P<0.05)。
2.6 低溫脅迫對甘藍型冬油菜內源IAA含量的影響
內源IAA含量的測定結果表明(圖8),隨著溫度降低,內源IAA含量降低。在處理24 h時,相比于對照(22℃),‘16VHNTS158’中IAA含量在4℃、0℃、-4℃時分別降低了68%、92%、97%(圖8A)。‘天油2288’中分別降低了77%、89%、94%(圖8B)。相同處理時間及溫度下‘16VHNTS158’內源IAA含量較‘天油2288’低,其中在處理24 h時,‘16VHNTS158’較‘天油2288’IAA含量在常溫(22℃)、4℃、0℃、-4℃分別低19%、14%、40%、39%。

圖8 甘藍型冬油菜根中內源IAA含量對低溫的響應
2.7 甘藍型冬油菜BnYUCCA8基因表達量、生長素含量與下胚軸長度之間的相關關系
由表2可知,下胚軸伸長較短的強抗寒性品種‘16VHNTS158’中BnYUCCA8基因相對表達量和生長素含量之間存在顯著正相關關系,相關系數為0.682,下胚軸長度與BnYUCCA8基因相對表達量、生長素含量之間存在正相關關系,相關系數分別為0.540和0.427;下胚軸伸長較長的弱抗寒性品種‘天油 2288’中BnYUCCA8基因相對表達量和生長素含量之間存在極顯著正相關關系,相關系數為0.812,下胚軸長度與BnYUCCA8基因相對表達量、生長素含量之間存在正相關關系,相關系數分別為0.410和0.591。

表2 甘藍型冬油菜BnYUCCA8基因表達量、生長素含量與下胚軸長度的Pearson相關
3 討 論
YUCCA是植物IAA合成過程中的限速酶,其生物學功能在多種植物中得到了廣泛挖掘[19-20],YUCCA8基因屬于YUCCA家族成員,目前,在甘藍型冬油菜中未見有YUCCA8基因相關研究報道。本文克隆的YUCCA8基因CDS序列為1 281 bp,編碼426個氨基酸,利用生物信息學分析其是一個不穩定的疏水性蛋白,不同物種間具有高度保守性,袁美同等[21]通過對梨YUCCA基因家族鑒定得出PbYUCCA8蛋白編碼423個氨基酸,不穩定指數為48.17,總親水性平均系數為-0.167,也是一個不穩定的疏水性蛋白。
甘藍型冬油菜BnYUCCA8基因時空表達特征分析結果表明,其在葉和根中表達量最高,莖和下胚軸中次之,花、花蕾和角果中表達量相對較低。擬南芥中YUCCA8轉錄水平在幼葉中最高,在成熟的葉、根和花芽中較低,莖中幾乎檢測不到[22],與本文在莖中的表達量不同可能與物種間差異有關。YUCCA8過表達的擬南芥下胚軸較長[23],而yuc8突變可以抑制光信號途徑的關鍵轉錄因子 PIF4過表達,從而抑制其下胚軸伸長[16],本文研究發現,YUCCA8基因的表達量在‘天油2288’(下胚軸伸長較長)較‘16VHNTS158’(下胚軸伸長較短)中低,表明YUCCA8基因在調控下胚軸長短的過程中發揮重要作用。
目前,YUCCA8基因響應溫度的研究主要集中于高溫誘導下YUCCA8調控下胚軸長短[15],而對低溫條件下YUCCA8與生長素及下胚軸的相關關系研究甚少,因此,本文通過對甘藍型冬油菜進行低溫脅迫處理,測定BnYUCCA8基因的表達量變化,結果表明隨著溫度降低,BnYUCCA8基因表達量整體呈降低趨勢,在0℃較4℃時表達量略有升高,與內源IAA對低溫脅迫的適應趨勢基本一致。表明BnYUCCA8基因可能通過生長素的調控機制參與了對油菜低溫脅迫的響應。
低溫脅迫通常會引起植物體內激素水平發生變化,從而對植物生理代謝活動作出調節反應[10]。本文通過對低溫脅迫下內源IAA含量測定發現,隨著溫度降低,2個抗寒性不同的甘藍型冬油菜內源IAA含量不斷降低,且下胚軸伸長較長的弱抗寒品種‘天油2288’較下胚軸伸長較短的強抗寒品種‘16VHNTS158’內源IAA含量高,表明內源IAA含量與甘藍型冬油菜的抗寒性呈負相關關系。韋霞霞等[24]通過對葡萄抗寒性與生長素積累相關性分析發現,生長素(IAA)在抗寒性越強的品種中含量越低。許耀照等[25]研究表明,不同抗寒性白菜型冬油菜品種葉片IAA含量隨著生育進程中氣溫的降低均呈先升高后降低的趨勢,進入越冬階段之后,弱抗寒品種葉片和根中IAA含量均高于強抗寒品種。劉世紅等[26]采用ELISA技術測定自然降溫過程中油棕葉片內源激素含量變化,研究發現,從26℃降到18℃,IAA含量基本不變,低于18℃后,IAA含量顯著增加,抗寒性強的品種IAA含量高,表明IAA含量的增加利于提高抗寒力,這可能與10℃以上低溫處理過程有關。
通過對甘藍型冬油菜BnYUCCA8基因表達量、內源IAA含量和下胚軸長度進行相關性分析,結果表明2個抗寒性不同材料的甘藍型冬油菜3個指標間的相關性表現一致,BnYUCCA8的基因表達量和內源IAA含量呈顯著正相關關系,下胚軸長度與BnYUCCA8基因表達量和IAA含量呈正相關關系,證實BnYUCCA8是生長素調控關鍵基因,且其在下胚軸伸長程度方面發揮正向調節作用。但是關于該基因在甘藍型冬油菜低溫處理過程中的作用機制及相關功能還有待進一步研究。
4 結 論
本研究從甘藍型冬油菜中克隆到YUCCA8基因,命名為BnYUCCA8,CDS序列為1 281 bp,編碼426個氨基酸,是一個不穩定的疏水性蛋白,不同物種間具有高度保守性。時空表達分析表明,其在根和葉中表達量最高,花和角果中相對較低。低溫脅迫下BnYUCCA8表達量整體呈降低趨勢,內源IAA含量與其表達量變化趨勢基本一致,且‘天油2288’(下胚軸伸長較長且抗寒性較弱)較‘16VHNTS158’(下胚軸伸長較短且抗寒性較強)的BnYUCCA8表達量低,內源IAA含量也低。BnYUCCA8表達量與內源IAA含量呈顯著正相關關系,下胚軸長度與BnYUCCA8表達量和內源IAA含量呈正相關關系,表明甘藍型冬油菜BnYUCCA8基因可能在調控下胚軸長度和內源IAA含量中發揮重要調節作用。
