直播稻金粳818成熟胚遺傳轉化體系的建立
2022-05-17 00:42:45李軍玲張紅磊張融雪蔡昱萌汪穎佟卉劉燕清蘇京平
安徽農業科學 2022年9期
李軍玲 張紅磊 張融雪 蔡昱萌 汪穎 佟卉 劉燕清 蘇京平
摘要 [目的]建立高效的金粳818成熟胚遺傳轉化體系。[方法]以直播稻品種金粳818成熟胚為外植體,設計4種誘導培養基,篩選適合金粳818的誘導條件;設計5種分化培養基,篩選適合金粳818的分化條件;獲得的愈傷組織用農桿菌介導法進行轉基因再生。[結果]最佳誘導培養基為4.10 g/L NB+2 mg/L 2,4-D,誘導率為89.3%;最佳分化培養基為MS+5.0 mg/L KT+0.5 mg/L NAA,分化率為81.1%,成苗率為52.1%;經PCR分子檢測,再生幼苗100%為轉基因陽性植株。[結論]該研究建立了金粳818成熟胚遺傳轉化體系,為品種定向改良奠定了技術基礎。
關鍵詞 金粳818;成熟胚;愈傷組織;遺傳轉化
中圖分類號 S 511? 文獻標識碼 A? 文章編號 0517-6611(2022)09-0100-04
doi:10.3969/j.issn.0517-6611.2022.09.024
開放科學(資源服務)標識碼(OSID):
Establishment of Genetic Transformation System of Mature Embryo of Direct Seeding Japonica Rice Variety Jingeng 818
LI Jun-ling, ZHANG Hong-lei, ZHANG Rong-xue et al
(Crop Research Institute, Tianjin Academy of Agricultural Sciences, Tianjin Key Laboratory of Crop Genetics and Breeding, Tianjin 300384)
Abstract [Objective]To establish an efficient genetic transformation system of Jingeng 818. [Method]Using the mature embryo of direct seeding rice variety Jingeng 818 as explant, four induction medium schemes were designed to obtain the suitable induction conditions of Jingeng 818;five kinds of differentiation medium were designed to obtain the suitable differentiation condition for Jingeng 818; and the obtained callus was used to establish the transgenic regeneration system by agrobacterium mediated method. [Result]The results showed that the best induction medium was 4.10 g/L NB+2 mg/L 2,4-D with the induction rate 89.3%. The best differentiation medium was MS+5.0 mg/L KT + 0.5 mg/L NAA with the differentiation rate 82.1% and the seedling rate 52.1%. By PCR molecular detection, 100% of the regenerated seedlings were transgenic positive plants.[Conclusion] In this study, the genetic transformation system of mature embryo of Jingeng 818 was established, which laid a technical foundation for directional improvement of varieties.
Key words Jingeng 818;Mature embryo;Callus;Genetic transformation
水稻是我國第一大口糧作物,水稻生產事關國家口糧安全。稻田雜草防除是生產中的棘手問題,由于禾本科雜草、雜草稻等與水稻親緣關系較近,常規除草劑對其無效,培育具有除草劑抗性水稻品種能較好地解決稻田草害問題。金粳818是天津市水稻研究所選育的具有除草劑抗性的粳型常規水稻品種,為采用生物技術手段對金粳818進行改良而培育的新種質,可為除草劑抗性水稻提供新材料,其遺傳轉化體系的建立尤為重要。
利用愈傷組織再生體系進行水稻遺傳轉化是最廣泛應用的方法之一,具有明顯優點,如外植體來源廣泛、周期短、易于轉化、轉化效率高等[1];農桿菌介導法是水稻遺傳轉化首選方法,具有易操作、低拷貝、基因沉默少、遺傳穩定等諸多優勢[2]。相比秈稻而言,粳稻愈傷誘導率和分化再生率較高,不少粳稻品種的組織培養再生體系已經建立且轉基因體系成熟,如日本晴、中花11、秀水134等[3-7]。影響水稻愈傷組織誘導率、分化率和再生率的主要因素是基因型[8],而品種間基因型的差異導致尚缺乏統一的配方適合所有品種。筆者以成熟胚為外植體,設計了4種不同誘導培養基摸索適宜誘導條件,設計5種分化培養基,利用獲得的愈傷組織以農桿菌介導法進行轉基因再生,建立高效的金粳818成熟胚遺傳轉化體系,為利用生物技術定向改良奠定基礎。
1 材料與方法
1.1 材料
以天津市水稻研究所自主育成的金粳818成熟胚為外植體。含有pCAMBIA1301質粒的EHA105農桿菌由天津市農作物遺傳育種重點實驗室保存。培養基NB(貨號N492)和MS(貨號M519)及激素均購自Phytotechnology。
1.2 培養基
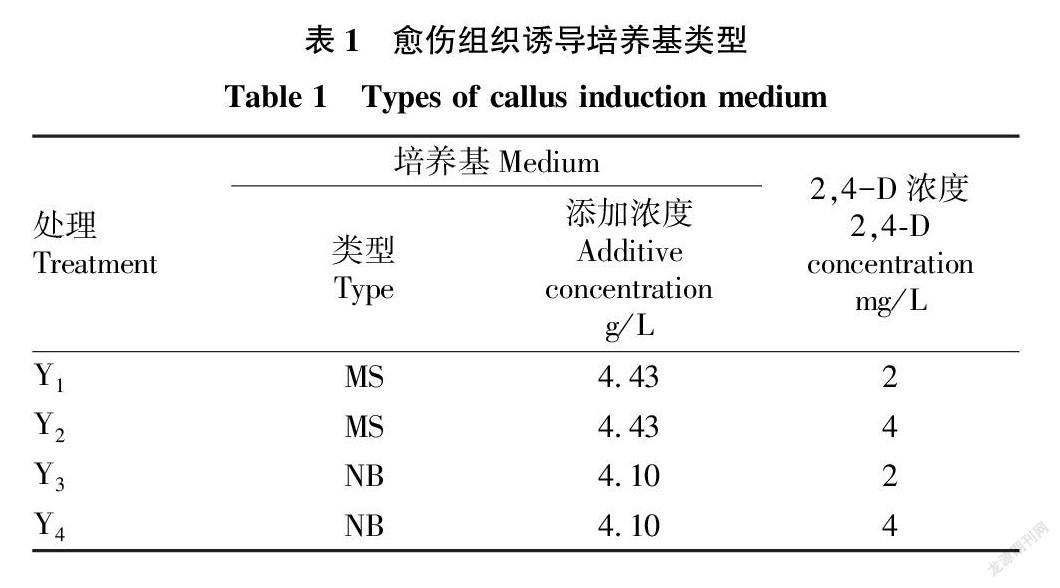
1.2.1 誘導培養基。基礎培養基+0.5 g/L L-脯氨酸+0.5 g/L谷氨酰胺+0.3 g/L水解酪蛋白+30 g/L蔗糖+3.5 g/L 植物凝膠+2,4-D(表1),調節pH為5.8。
1.2.2 篩選培養基。4.1 g/L NB+0.5 g/L L-脯氨酸+0.5 g/L谷氨酰胺+0.3 g/L 水解酪蛋白+30 g/L蔗糖+3.5 g/L? 植物凝膠+2 mg/L 2,4-D+40 μg/L潮霉素+400 μg/L青霉素,調節pH為5.8。
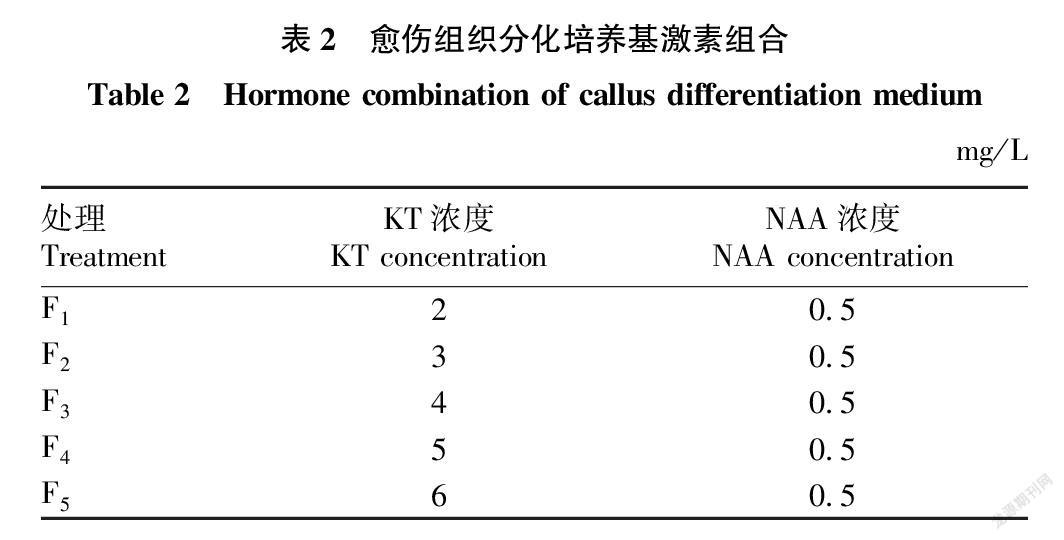
1.2.3 分化培養基。4.43 g/L MS+0.5 g/L L-脯氨酸+0.5 g/L谷氨酰胺+0.3 g/L 水解酪蛋白+0.1 g/L 肌醇+30 g/L 山梨醇+30 g/L蔗糖+3.5 g/L 植物凝膠+30 μg/L潮霉素+300 μg/L青霉素+不同激素組合(表2),調節pH為5.8。
1.2.4 生根培養基。2.215 g/L MS+30 g/L蔗糖+3.5 g/L植物凝膠+20 μg/L潮霉素+200 μg/L青霉素,調節pH為5.8。
1.2.5 AAM侵染液。4.1 g/L NB+68 g/L蔗糖+36 g/L 葡萄糖+0.5 g/L水解酪蛋白+0.5 g/L L-脯氨酸+0.5 g/L L-谷氨酰胺+177 mg/L L-精氨酸+7.5 mg/L甘氨酸+300 mg/L L-天冬氨酸+2.5 mg/L 2,4-D,調節pH為5.2,使用前加入0.5 mL濃度為20 mg/L乙酰丁香酮。
1.3 方法
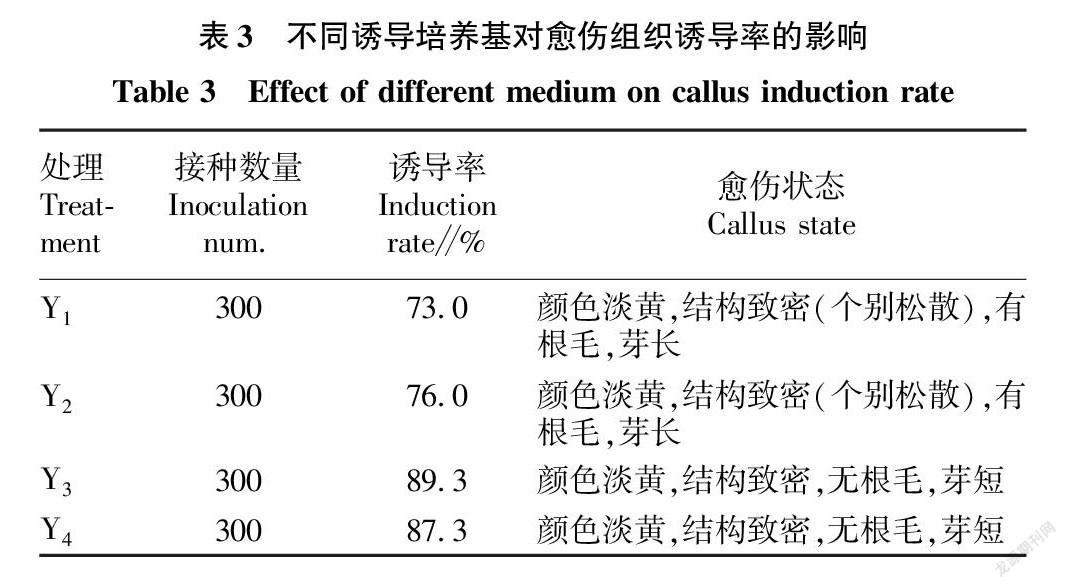
1.3.1 金粳818成熟胚愈傷組織誘導。收取當年新的金粳818成熟種子,剝離穎殼,取適量倒入50 mL離心管中,75%乙醇消毒1 min,30%次氯酸鈉(有效氯3%)消毒15 min,無菌水沖洗5~6次,去除多余水分,倒入鋪有3層滅菌濾紙的培養皿中,無菌風吹干15 min。最后接種到誘導培養基中,每皿25~35粒種子,28 ℃暗培養10 d,在水稻幼芽和成熟胚之間長出淡黃色、結構致密的胚性愈傷組織,掐芽剝離出愈傷后放入新的誘導培養基上3~5 d即可進行轉化,誘導10 d后統計誘導率。
誘導率=出愈傷的成熟胚數量/總成熟胚數量×100%
1.3.2
農桿菌培養及浸染。將含有pCAMBIA1301質粒的農桿菌EHA105在YEP(含25 mg/L卡那霉素)固體平板上劃線,28 ℃培養2 d。刮取平板上的農桿菌至AAM浸染液中,調整菌體濃度至OD600值為0.3~0.5,獲得農桿菌懸浮液。挑取足夠數量的愈傷組織放入無菌三角瓶中,加入農桿菌懸浮液,室溫放置浸染20 min,并間隔晃動幾次,倒掉菌液,將愈傷組織放置無菌濾紙上吸去多余菌液,轉移至鋪有一層無菌濾紙的固體培養基上,26 ℃黑暗培養3 d。
1.3.3 篩選培養。培養3 d后的愈傷組織,用無菌水清洗2次,用含有500 μg/mL羧芐青霉素的無菌水沖洗1次,吸去多余水分后將愈傷組織轉移到無菌濾紙上,晾干,轉移至篩選培養基上進行篩選培養,28 ℃,暗培養30 d。
1.3.4 分化再生。篩選30 d后,在原愈傷的底部可見顏色淡黃、結構致密、直徑1~2 mm的陽性愈傷長出,挑取陽性愈傷到分化培養基上進行分化再生,培養條件為28 ℃,16 h光照/8 h黑暗培養。15~20 d統計分化率,25~35 d統計成苗率。
分化率=分化的愈傷組織數量/總愈傷組織數量×100%
成苗率=分化的幼苗數量/分化的愈傷組織數量×100%
1.3.5 生根培養及馴化。分化出的幼苗長到2~3 cm有明顯根系時,轉移到生根培養基上,培養條件28 ℃,16 h光照/8 h黑暗培養。7~10 d后,幼苗長出大量側根,當小苗長至高10~15 cm時,打開蓋子在室外煉苗2~3 d,取出小苗后洗去根部培養基,移栽至土中。
1.3.6 轉基因植株鑒定。CTAB法提取幼苗DNA,并用潮霉素基因引物檢測植株。
hygF:ACGGTGTCGTCCATCACAGTTTGCC
hygR:TTCCGGAAGTGCTTGACATTGGGGA
2 結果與分析
2.1 培養基和2,4-D濃度對愈傷組織誘導的影響
共設計4組誘導培養基,包含2種類型基礎培養基和2種濃度2,4-D。試驗結果表明,同一激素濃度下,NB培養基的誘導率高于MS培養基的誘導率,即Y3>Y1,Y4>Y2,且愈傷組織狀態更好,無明顯根毛,芽較短。同一基礎培養基條件下,4和2 mg/L 2,4-D對愈傷誘導率差別不大。綜上,適宜的誘導培養基為Y3( 4.10 g/L NB+2 mg/L 2,4-D),誘導率為89.3%,且愈傷狀態良好(表3和圖1)。
2.2 不同濃度KT對愈傷組織分化再生的影響
設計了5種不同KT濃度分化培養基,金粳818愈傷組織在不同KT濃度分化培養基中分化成苗差異明顯。試驗數據表明,同一濃度NAA條件下,一定范圍內KT濃度越高,分化率和成苗率越高,表現為F4>F3>F2>F1,KT濃度升高至6 mg/L,其分化率和成苗率反而有所下降。分化21 d時,F4處理綠點數量最多,個別愈傷組織分化出小苗(圖2)。因此,適宜分化培養基激素配比為F4處理,即5 mg/L KT+0.5 mg/L NAA,可使分化率和成苗率分別達到81.1%和52.1%(表4)。
2.3 轉基因植株分子鑒定
提取24份再生幼苗基因組DNA作為模板,利用潮霉素基因特異引物進行PCR鑒定,結果表明,所有再生苗均為轉基因植株(圖3),農桿菌介導的遺傳轉化成功。
安徽農業科學2022年
2.4 金粳818成熟胚遺傳轉化完整過程
金粳818成熟胚誘導愈傷需10 d,愈傷繼代5 d,轉化共培養3 d,篩選30 d,分化30 d,生根約7 d,整體遺傳轉化過程共計約85 d完成(圖4)。
3 結論與討論
影響水稻愈傷分化和再生的主要因素有基因型、外植體來源、培養基、激素等[8]。水稻基因型是控制愈傷組織誘導率和再生率的主要因素,有研究表明,粳稻的愈傷組織誘導率及分化率明顯優于秈稻[9-11]。水稻常用的外植體有花藥、幼穗、幼胚及成熟胚,不同組織的愈傷誘導率和分化率不同,但由于花藥、幼穗、幼胚等外植體受到生長季節限制,取材不便,從而制約了其可用于遺傳轉化的數量,而成熟胚則不受限制[12-13]。劉書梅等[14]研究結果表明,MS和NB培養基分別更適合秈稻種胚愈傷和粳稻種胚愈傷的誘導培養,這與該研究結果一致,對金粳818而言,NB培養基誘導的愈傷狀態更好。激素種類、濃度和配比是影響愈傷組織誘導、分化再生的重要調控因子,其中2,4-D是誘導胚性愈傷組織必不可少的因子,該研究使用2 mg/L 2,4-D時,可達到較高誘導率;KT和6-BA是愈傷分化常用的細胞分裂素,沈秋平等[15]研究認為KT比6-BA分化效果要好。筆者研究了不同濃度KT對分化成苗率的影響,結果表明,一定范圍內KT濃度越高,分化成苗率越高。
綜上,該研究以金粳818成熟胚為外植體,設計4種激素配比不同的誘導培養基,將金粳818成熟胚外植體接種在誘導培養基中,10 d后觀察愈傷狀態并統計每組誘導率,得出適宜誘導培養基為4.10 g/L NB+2 mg/L 2,4-D,誘導率為89.3%;設計5種激素配比不同的分化培養基,將金粳818愈傷組織接種后進行分化培養,統計每組分化率和成苗率,得出適宜分化培養基為MS+5 mg/L KT+0.5 mg/L NAA,其分化率和成苗率分別是81.1%和52.1%;整個遺傳轉化體系從誘導到幼苗再生共需要85 d。金稻818成熟胚遺傳轉化體系的建立,為利用生物技術手段定向改良品種奠定了技術基礎。
參考文獻
[1] 佟清越.粳型水稻遺傳轉化體系的研究[D].哈爾濱:黑龍江大學,2010.
[2] 彭小群,王夢龍.水稻遺傳轉化研究進展[J].中國農學通報,2021,37(27):1-5.
[3] 毛建軍,楊秀芬,曾洪梅,等.水稻品種日本晴粳稻組培培養基的篩選及轉稻瘟菌蛋白激發子基因植株的獲得[J].農業生物技術學報,2008,16(5):824-830.
[4] 何少海,姚盛存,豐宇凱,等.農桿菌介導bar基因轉化水稻胚性愈傷組織的研究[J].浙江農林大學學報,2017,34(1):129-136.
[5] 曹含章,沈秋平,袁冰,等.3個粳稻品種成熟胚分化和再生適宜激素配比研究[J].雜交水稻,2020,35(4):86-91.
[6] 李素娟,樊秀霞,王華,等.水稻不同品種組培再生和轉基因頻率研究[J].核農學報,2013,27(12):1817-1827.
[7] 張秀香.水稻成熟種子組織培養體系的優化及建立[J].安徽農學通報,2012,18(12):39-40,42.
[8] 朱克明,陶慧敏,徐碩,等.水稻愈傷分化和再生研究進展[J].種子,2016,35(11):55-59.
[9] ABE T,FUTSUHARA Y.Genotypic variability for callus formation and plant regeneration in rice(Oryza sativa L.)[J].Theor Appl Genet,1986,72(1):3-10.
[10] 王萍,徐大勇,王罡,等.粳秈稻兩個亞種成熟胚組織培養與再生能力的比較研究[J].種子,2007,26(10):66-67.
[11] YAN J Q,XUE Q Z,ZHU J.Genetic studies of anther culture ability in rice(Oryza sativa)[J].Plant Cell Tissue Organ Cult,1996,45(3):253-258.
[12] 王秀紅,史向遠,吳先軍.水稻不同外植體培養效果及其相關性分析[J].中國水稻科學,2005,19(2):187-189.
[13] 鄭文靜,張燕之,王昌華,等.水稻不同外植體組織培養的差異性及其后代變異的研究[J].安徽農業科學,2008,36(4):1368-1370.
[14] 劉書梅,王姍姍,辛曉云,等.不同培養基對水稻胚性愈傷組織誘導及分化的影響[J].生物學雜志,2011,28(4):54-56.
[15] 沈秋平,嚴孫藝,薛宇彤,等.秈稻9311成熟胚高效再生體系的建立[J].雜交水稻,2019,34(2):49-52.
