Bacillus屬嗜鹽嗜堿細菌的研究進展
2022-05-17 23:46:07李冬瑩王爽肖洋
安徽農業科學 2022年9期
關鍵詞:分類
李冬瑩 王爽 肖洋
摘要 Bacillus屬由于其成員表現出廣泛的多系性以及彼此間幾乎沒有共同點,長期以來一直被認為是分類和系統發育異常的屬。隨著測序技術和純培養技術的進步,自Cohn建立Bacillus屬以來,不斷有新的菌株從不同生境中被分離,也有許多該屬的成員被重新分類為其他的屬。Bacillus屬由于其具有芽孢的特性而廣泛分布于自然界的不同環境,其嗜鹽嗜堿成員已經成為人們高度重視的雙重極端微生物類群,在鹽堿的雙重壓力下經過長期的進化選擇,形成了獨特的生物活性物質代謝途徑、新的調控機制及與這些生理生化特征有關的新基因類型。因此,針對Bacillus屬中嗜鹽嗜堿菌的發現歷史、分布與分類現狀、生理生化特征及應用前景進行綜述,以期為嗜鹽嗜堿芽孢桿菌新物種資源的挖掘及功能的開發利用提供參考。
關鍵詞 嗜鹽嗜堿細菌;芽孢桿菌;分類;嗜鹽機制;嗜堿機制
中圖分類號 Q 935? 文獻標識碼 A
文章編號 0517-6611(2022)09-0019-04
doi:10.3969/j.issn.0517-6611.2022.09.006
開放科學(資源服務)標識碼(OSID):
Research Progress on Halophilic and Alkaliphilic Bacteria of Genus Bacillus
LI Dong-ying1,WANG Shuang2,XIAO Yang1
(1.College of? Modern? Agriculture and Ecological Environment,Heilongjiang University,Harbin,Heilongjiang 150080;2.Institute of Soil Fertilizer and Environmental Resources,Heilongjiang Academy of Agricultural Sciences,Harbin,Heilongjiang 150086)
Abstract The genus Bacillus has long been recognized as a taxonomic and phylogenetic anomaly as its different members exhibit extensive polyphyly and share very little in common with one another.As the development of sequencing and pure culture technology,since Cohn established the genus Bacillus,many novel strains have been isolated from various habitats,and some members of this genus have been reclassified into other genera.Bacillus spp.was widely distributed in different natural environments because of its spore producing.Halophilic and alkaliphilic bacillus have become a type of double extremophiles functioning optimally in saline and alkaline habitats.Under the dual pressure of salinity and alkalinity,through long-term evolutionary selection,these microorganisms have formed unique metabolic pathways of bioactive substances,new regulatory mechanisms and new gene type related to these physiological and biochemical characteristics.Therefore,in this paper,the discovery history,distribution and classification status,physiological and biochemical characteristics and application prospects of halophilic and alkalophilic Bacillus spp.were reviewed,in order to provide reference for the exploration of new species resources and the development and utilization of functions of halophilic and alkalophilic Bacillus spp.
Key words Halophilic and alkaliphilic bacteria;Bacillus;Taxonomy;Halophilic mechanism;Alkaliphilic mechanism
由于鹽化或堿化導致的土壤退化問題是一個重大的環境障礙,給農業生產及可持續發展帶來了不利影響。據統計,我國鹽堿化土壤總面積超過3 000萬hm2,并占據總耕地面積的10%,甚至更多[1]。蘇打鹽堿土對作物的危害程度較其他鹽堿土亞類重,不僅形成缺苗燒苗現象,而且作物生長期間也常因堿害而出現黃萎、爛根等病癥,使植株生長不旺、產量低。我國的蘇打鹽堿土主要分布于黑龍江、吉林兩省的西部區域與內蒙古東部的草原與干草原的堿池子(即塘)周圍和低濕地區[2]。
蘇打鹽堿土的鹽分積累雖然與含有蘇打的地下水有關,但蘇打的形成則與土壤中微生物活動有密切的關系[3]。Na2CO3和NaHCO3是蘇打鹽堿土中鹽分的重要成分,而硫酸鹽和氯鹽含量只在蘇打鹽堿土中占有較小的比例,土壤呈強堿性反應(pH 9~10)[4],同時土壤膠體中有多量的代換性鈉,土壤遇水易分散,干后板結堅硬,耕性差。在這些條件下生存的生物無疑具有獨特的生理特征、表型特性和適應機制,這些機制能夠控制細胞膜的通透性、控制細胞內滲透平衡、細胞壁的穩定性、細胞內蛋白質和其他細胞成分[5]。由于這些生物具有在鹽和堿環境中生長的適應能力,所以有越來越多的嗜鹽和嗜堿性的微生物被發現。芽孢桿菌屬中包含極端微生物,并有非常大的核糖體16S多樣性,具有顯著的生物技術潛力。芽孢桿菌相關菌種可在各種環境中生長(pH 2~12,溫度5~78 ℃,鹽度0~30% NaCl)[6]。自1986年以來,我國已經發表了近2 000篇關于嗜鹽嗜堿芽孢桿菌的科學論文,該研究綜述嗜鹽嗜堿菌中芽孢桿菌的發展和其獨特的生理機制,特別是在現代各領域上的應用,以期為嗜鹽嗜堿芽孢桿菌新物種資源的挖掘及功能的開發利用提供參考。
1 嗜鹽嗜堿Bacillus細菌的分布與分類多樣性
早在2.5億年前,Bacillus spp.作為土壤中的腐生菌就大量存在,可見,對于芽孢桿菌屬的研究有著十分漫長的歷程,該屬在微生物學的領域中具有舉足輕重的地位[7]。它們在自然界中分布十分廣泛,從荒漠[8]、溫泉[9]到北極土壤[10],從淡水[11]到海洋沉積物[12],甚至存在于飲水處理廠[13]、古墓[14]和生物電能源[15]中,這類革蘭氏陽性的芽孢桿菌無處不在[16]。Kushner[17]定義嗜鹽菌為最適生長鹽濃度在0.2 mol/L(相當于1.17 %NaCl)以上。隨后,Kroll[18]定義嗜堿菌最適生長pH為9.0以上。Chester于1889首次描述了嗜堿細菌Sporosarcina pasteurii(舊分類系統里稱Bacillus pasteurii),1934年,Vedder[19] 報道Bacillus alcalophilus為Bacillus屬的第二個嗜堿成員,可在pH 8.6~11.0的條件下生長良好,但是在“普通”pH條件下不能生長。1982年,Gordon和Hyde將為數眾多的嗜堿Bacillus菌株進行分類。2020年,Patel等[20]將部分嗜鹽嗜堿Bacillus菌株歸類為Alkalihalobacillus屬。
2 嗜鹽嗜堿Bacillus細菌的特征
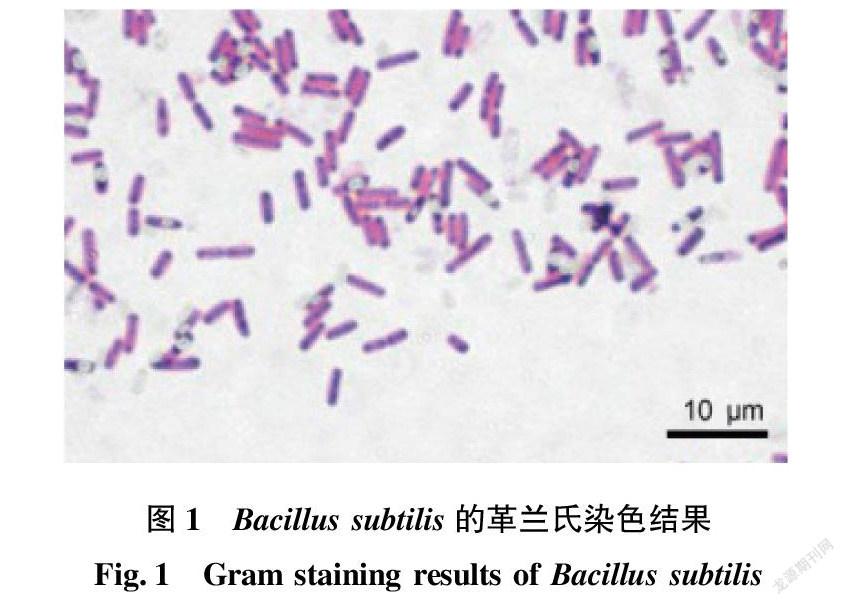
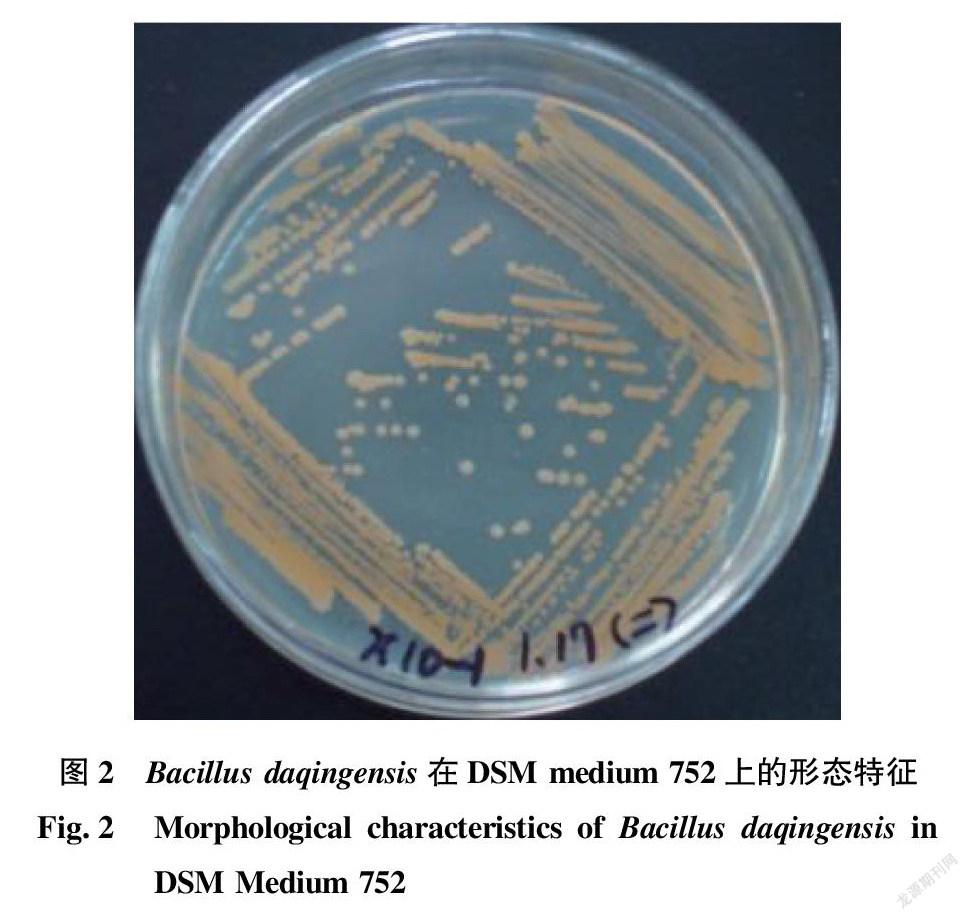
Bacillus屬的成員一般為嚴格好氧或兼性厭氧的化能有機營養類型,革蘭氏陽性、內生孢子、直或彎曲的桿狀細菌(圖1)。然而,非孢子構成的細菌在這個屬里被發現,這些細菌是否缺少孢子形成的基因,是否在進化過程中失去了這些基因(或部分基因),亦或在生理上無法成功地形成孢子,仍有待澄清[21]。B.saliphilus是芽孢桿菌屬中唯一的嗜鹽嗜堿的非孢子球菌[22]。Bacillus屬的菌株菌落不透明、乳白色或褐色,在某些培養基上可以產生色素,如Wang等[23]報道的嗜鹽堿芽孢桿菌屬的新種Bacillus daqingensis,其模式菌株X10-1可在DSM 752培養基(DSMZ)上顯橙色菌落(圖2)。跟大多數細菌的運動方式一樣,嗜堿芽孢桿菌利用絲狀或螺旋槳狀鞭毛來運動。對嗜堿芽孢桿菌的調查表明,嗜堿芽孢桿菌C-125的鞭毛蛋白與枯草芽孢桿菌有較高的同源性,該菌株的鞭毛形成與pH有關,在堿性pH條件下鞭毛呈扁平狀,具有很強的動力;在中性pH條件下只有少量的直鞭毛,且鞭毛不活動,嗜堿芽孢桿菌的運動也是具有Na+依賴性的。
3 嗜鹽嗜堿微生物的生理機制
嗜鹽(halophilic)和耐鹽(halotolerant)的微生物細胞具有吸K+排Na+的作用,在維持細胞內滲透壓的過程中起到至關重要的作用[24]。而且嗜鹽微生物會攝取、合成并積累具有親和性的溶質,如氨基酸、嘧啶、糖類等小分子物質,這些物質都是高度可溶的,在此過程中,細胞的代謝并不受影響,同時胞內的水活度也得到了提高。這些親和性溶質的聚集恰好填補了外界高滲透壓條件下導致的內外部滲透壓的不均衡,在緩解高滲透壓環境對于細胞的脅迫中具有一定的支撐作用,同時也有助于嗜鹽微生物順利應對高鹽環境下的滲透壓[25-26]。
嗜堿(alkaliphilic)和耐堿(alkalitolerant)的微生物可以耐受對于嗜中性微生物來說致命性的環境,然而相比較外界的堿性pH,這些微生物的細胞質pH是維持近中性的[27]。細胞外被(細胞壁和細胞膜)在堿的耐受過程中起了重要作用,細胞壁充當許多生物體的第一道屏障。相對于嗜中性菌而言,嗜堿菌的膜與壁有著更加特殊的成分,嗜堿菌的細胞壁具有很多酸性化合物,如半乳糖醛酸、葡糖酸、谷氨酸、磷酸,這些化合物均帶有負電荷,因而可降低細胞表面pH,這是嗜堿菌對堿性環境的適應。此外,嗜堿菌的質膜能夠通過Na+/H+、K+/H+反向轉運載體系統來維持細胞內pH的相對穩定[28-29]。
4 嗜鹽嗜堿Bacillus細菌基因組研究
嗜鹽嗜堿Bacillus細菌的G+C含量普遍不高,單純嗜鹽的Bacillus中G+C含量普遍在35.0%~48.6%,單純嗜堿的Bacillus中G+C含量普遍在33.5%~43.9%,既嗜鹽又嗜堿的Bacillus中G+C含量普遍在36.3%~48.9%。嗜鹽嗜堿Bacillus細菌的基因組大小差異顯著,普遍在3.28~5.87 Mb。嗜堿桿菌是工業微生物中最重要的主力,為了獲得高pH條件下生理適應性的分子信息和研究這些嗜堿桿菌的工業潛力,特別是嗜堿桿菌菌株產生的獨特堿性酶的應用,第一個嗜堿芽孢桿菌Bacillus halodurans C-125的全基因組序列于2000年被測定[30]。后來,學者又分別測定了B.halodurans C-125(2000)和B.clausii KSM-K16(2005)等嗜堿桿菌的全基因組序列。雖然目前已發表的嗜鹽嗜堿Bacillus細菌的基因組數量較少,但其功能基因具有潛在的開發價值,其基因產物可運用于工業、農業、醫學等領域。因此,對嗜鹽嗜堿Bacillus細菌基因組數據的補充和分析是非常有必要的,不僅有利于揭示嗜鹽嗜堿Bacillus細菌的相關生理特性,還能在基因組水平上對具有不同生理代謝能力的嗜鹽嗜堿Bacillus細菌進行分析和比較,為未來嗜鹽嗜堿Bacillus細菌工程菌的遺傳改造提供必要的遺傳基礎。
5 嗜鹽嗜堿Bacillus細菌的應用
Bacillus屬成員的數量如此之大,毋庸置疑地說明了這個屬在環境中的重要性[31]。嗜鹽嗜堿芽孢桿菌在工業、農業、醫學方面具有重要的作用。
5.1 工業
近年來,對嗜堿Bacillus菌株研究的加強除了在分類學和生理學方面外,對于其應用方面也更為關注,由于它們能夠生產嗜堿酶,所以在工業上具有廣泛的應用價值[32-33]。嗜堿Bacillus生產的木聚糖酶具有水解木聚糖產生木糖和寡聚糖的能力,可用于處理人造纖維廢物,而堿性β-甘露聚糖酶降解甘露聚糖產生的寡糖可作為保健品的添加劑[34]。此外,來自嗜鹽堿的芽孢桿菌菌株中的堿性纖維素酶和堿性蛋白酶經常用于生物洗滌劑的添加劑,例如Bacillus halodurans CAS6和Bacillus horikoshii可以產生堿性蛋白酶[35-36],而堿性纖維素酶具有去除衣物污漬的作用,同時也能使衣服變得更加軟化和鮮艷,是一種工業上十分重要的洗滌劑[37]。這種酶可以利用堿性環麥芽糖糊精葡聚糖來生產環糊精,大大降低生產成本,為環糊精在食品、化工和醫藥領域的大批量應用開辟了新的市場[38]。
5.2 農業
來自不同物種的鹽堿壓力耐受基因今后可能被用于提高轉基因植物品系的產量,對農業生產具有重要意義[39]。嗜鹽堿芽孢桿菌及其產生的脂肽抗菌物質對水稻紋枯病、三期根腐病、煙草黑脛病等植物病害都具有良好的防治效果,國內已開發成功并投入商業化生產[40]。來自土壤的蘇云金芽孢桿菌Bacillus thuringiensis的“晶體蛋白”基因在棉花這類作物遺傳改造中的應用,也顯著表明來自嗜鹽嗜堿生物的基因可以為農業生產帶來新的變革[41-42]。朱燕云等[43]研究發現,由嗜鹽性的Bacillus subtilis subspecies stercoris? strain XP菌株合成的球形生物納米硒針對草莓病原真菌具有潛在的生防功能,并表現出了劑量依賴的效應,即納米硒越高其抑制活性越好,此研究還發現這種高濃度生物納米硒對處理草莓枯萎病病原菌、紫斑病病原菌、紅葉病病原菌均具有顯著的抑制效果,為草莓種植過程中有害病原真菌的防控提供了寶貴的參考信息,同時菌株XP合成的生物納米硒也有望在提高草莓品質方面發揮積極作用。
5.3 醫學
隨著醫學的不斷進步,芽孢桿菌在醫藥方面也有很多應用。嗜鹽堿性芽孢桿菌菌株中的堿性蛋白酶在醫療上可以用于消腫、清炎等各種炎癥的治療[44]。嗜堿性的Bacillus sp.菌株分泌的CK11-4具有溶解纖維蛋白的能力,能夠在醫學上用于溶解血栓[45]。此外,在國際市場中,這種堿性蛋白酶在胰島素、分析和藥物用酶中具有廣泛的應用[46]。我國在研究蛋白酶上也有了較大的進步,其中嗜堿性短小芽孢桿菌B45等分泌的蛋白酶已經得到了廣泛地生產和應用[47]。zgren等[48]研究發現,從嗜堿性的Bacillus marmarensis DSM 21297分離出的PHB具有生產生物聚合物的能力,并且比市面上的PHB更耐高溫,是制備用于組織工程應用的納米纖維復合支架的潛在候選材料,由于其毒性風險更低,因此對污染的耐藥性可能在醫學應用上具有更大的優勢。
6 結論與展望
由于高鹽堿環境中的微生物具有豐富的物種多樣性、耐鹽堿基因多樣性和代謝機制多樣性,具有產生多樣的生理活性物質的巨大潛力,因而將高鹽堿環境微生物作為尋求創新藥物的來源已成為眾多研究者的目標之一。伴隨嗜鹽嗜堿芽孢桿菌新物種的不斷增加,將會有越來越多的嗜鹽嗜堿芽孢桿菌基因組被測序,從而有助于進一步研究嗜鹽嗜堿芽孢桿菌的耐鹽堿分子機制,發掘出更多新的和耐鹽堿相關的基因資源,并廣泛應用于改造農作物、增加糧食產量等領域。
為了澄清嗜鹽嗜堿芽孢桿菌領域的生態功能和環境變化之間的關系,今后的研究工作不應僅局限于分離和保藏新的菌種,還要結合宏基因組技術來研究不同環境的嗜鹽嗜堿芽孢桿菌的多樣性,以加強人們對嗜鹽嗜堿芽孢桿菌這一極端微生物類群的生長環境及其分布特征的了解。另外,在基于群體遺傳學基礎上對許多細菌菌株的多基因位點進行遺傳分析,將有助于進一步揭示嗜鹽嗜堿芽孢桿菌的進化過程與環境分布之間的關系。
參考文獻
[1]
范王濤.土壤鹽堿化危害及改良方法研究[J].農業與技術,2020,40(23):114-116.
[2] 楊真,王寶山.中國鹽漬土資源現狀及改良利用對策[J].山東農業科學,2015,47(4):125-130.
[3] 楊賽,俞冰倩,胡信玉,等.東北蘇打鹽堿土壤微生物群落對植被進展演替的響應[J].土壤通報,2019,50(3):632-640.
[4] 俞冰倩,楊賽,朱琳,等.響應土壤陰離子類型的鹽堿土古細菌群落多樣性研究[J].中國環境科學,2018,38(7):2731-2739.
[5] MESBAH N M,WIEGEL J.Life under multiple extreme conditions:Diversity and physiology of the halophilic alkalithermophiles[J].Applied and environmental microbiology,2012,78(12):4074-4082.
[6] TAKAMI H.Genomics and evolution of alkaliphilic Bacillus species[M]//HORIKOSHI K.Extremophiles handbook.Tokyo:Springer,2011:183-211.
[7] VREELAND R H,ROSENZWEIG W D,POWERS D W.Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal[J].Nature,2000,407(6806):897-900.
[8] ZHANG L,WU G L,WANG Y,et al.Bacillus deserti sp.nov.,a novel bacterium isolated from the desert of Xinjiang,China[J].Antonie van leeuwenhoek,2011,99(2):221-229.
[9] NAZINA T N,LEBEDEVA E V,POLTARAUS A B,et al.Geobacillus gargensis sp.nov.,a novel thermophile from a hot spring,and the reclassification of Bacillus vulcani as Geobacillus vulcani comb.nov.[J] International journal of systematic and evolutionary microbiology,2004,54(6):2019-2024.
[10] YOU Z Q,LI J,QIN S,et al.Bacillus abyssalis sp.nov.,isolated from a sediment of the South China Sea[J].Antonie van leeuwenhoek,2013,103(5):963-969.
[11] BAIK K S,LIM C H,PARK S C,et al.Bacillus rigui sp.nov.,isolated from wetland fresh water[J].Journal of? systematic and evolutionary microbiology,2009,60(Pt9):2204-2209.
[12] JUNG M Y,KIM J S,PAEK W K,et al.Bacillus manliponensis sp.nov.,a new member of the Bacillus cereus group isolated from foreshore tidal flat sediment[J].The journal of microbiology,2011,49(6):1027-1032.
[13] VAZ-MOREIRA I,FIGUEIRA V,LOPES A R,et al.Bacillus purgationiresistans sp.nov.,isolated from a drinking-water treatment plant[J].International journal of systematic and evolutionary microbiology,2012 62(Pt1):71-77.
[14] GATSON J W,BENZ B F,CHANDRASEKARAN C,et al.Bacillus tequilensis sp.nov.,isolated from a 2000-year-old Mexican shaft-tomb,is closely related to Bacillus subtilis[J].International journal of systematic and evolutionary microbiology,2006,56(Pt7):1475-1484.
[15] MA C,ZHUANG L,ZHOU S G,et al.Alkaline extracellular reduction:Isolation and characterization of an alkaliphilic and halotolerant bacterium,Bacillus pseudofirmus MC02[J].Journal of applied microbiology,2012,112(5):883-891.
[16] DASTAGER S G,MAWLANKAR R,TANG S K,et al.Bacillus enclensis sp.nov.,isolated from sediment sample[J].Antonie van leeuwenhoek,2014,105(1):199-206.
[17] KUSHNER D J.Life in high salt and solute concentrations:Halophilic bacteria[M]//KUSHNER D J.Microbial life in extreme environments.London:Acdemic Press,1978:317-368.
[18] KROLL R G.Alkaliphiles[M]//EDWARDS C.Microbiology of extreme environments.New York:Oppen University Press,1990:55-92.
[19] VEDDER A.Bacillus alcalophilus n.sp.; benevens enkele ervaringen met sterk alcalische voedingsbodems[J].Antonie van leeuwenhoek journal of microbiology and serology,1934,1(1):141-147.
[20] PATEL S,GUPTA R S.A phylogenomic and comparative genomic framework for resolving the polyphyly of the genus Bacillus:Proposal for six new genera of Bacillus species,Peribacillus gen.nov.,Cytobacillus gen.nov.,Mesobacillus gen.nov.,Neobacillus gen.nov.,Metabacillus gen.nov.and Alkalihalobacillus gen.nov.[J].International journal of systematic and evolutionary microbiology,2020,70(1):406-438.
[21] FRITZE D.Taxonomy of the genus Bacillus and related genera:The aerobic endospore-forming bacteria[J].Phytopathology,2004,94(11):1245-1248.
[22] ROMANO I,LAMA L,NICOLAUS B,et al.Bacillus saliphilus sp.nov.,isolated from a mineral pool in Campania,Italy[J].International journal of systematic and evolutionary microbiology,2005,55(Pt1):159-163.
[23] WANG S,SUN L,WEI D,et al.Bacillus daqingensis sp.nov.,a halophilic,alkaliphilic bacterium isolated from saline-sodic soil in Daqing,China[J].Journal of microbiology,2014,52(7):548-553.
[24] 許珂,陳紅,劉志千,等.新疆鹽堿地土壤耐鹽芽胞桿菌的分離和鑒定[J].生物資源,2019,41(2):174-178.
[25] 王偉偉,唐鴻志,許平.嗜鹽菌耐鹽機制相關基因的研究進展[J].微生物學通報,2015,42(3):550-558.
[26] VENTOSA A,NIETO J J,OREN A.Biology of moderately halophilic aerobic bacteria[J].Microbiology and molecular biology reviews,1998,62(2):504-544.
[27] HORIKOSHI K.General physiology of alkaliphiles[M]//HORIKOSHI K,ANTRANIKIAN G,BULL A T,et al.Extremophiles handbook.Tokyo:Springer,2011.
[28] 孫瑩,蘇進進,李潮流,等.可可西里堿性土壤樣品細菌的分離和生物學特性[J].微生物學通報,2011,38(10):1473-1481.
[29] 王建明,關統偉,賀江舟,等.嗜堿菌的研究進展及利用前景[J].陜西農業科學,2008,54(1):83-86.
[30] TAKAMI H,NAKASONE K,TAKAKI Y,et al.Complete genome sequence of the alkaliphilic bacterium Bacillus halodurans and genomic sequence comparison with Bacillus subtilis[J].Nucleic acids research,2000,28(21):4317-4331.
[31] YANG G Q,CHEN M,YU Z,et al.Bacillus composti sp.nov.and Bacillus thermophilus sp.nov.,two thermophilic,Fe(III)-reducing bacteria isolated from a compost[J].International journal of systematic and evolutionary microbiology,2013,63(8):3030-3036.
[32] HORIKOSHI K.Alkaliphiles:Some applications of their products for biotechnology[J].Microbiology and molecular biology reviews,1999,63(4):735-750.
[33] NOGI Y,TAKAMI H,HORIKOSHI K.Characterization of alkaliphilic Bacillus strains used in industry:Proposal of five novel species[J].International journal of systematic and evolutionary microbiology,2005,55(Pt6):2309-2315.
[34] TAMAKI H,SEKIGUCHI Y,HANADA S,et al.Comparative analysis of bacterial diversity in freshwater sediment of a shallow eutrophic lake by molecular and improved cultivation-based techniques[J].Applied and environmental microbiology,2005,71(4):2162-2169.
[35] ANNAMALAI N,RAJESWARI M V,THAVASI R,et al.Optimization,purification and characterization of novel thermostable,haloalkaline,solvent stable protease from Bacillus halodurans CAS6 using marine shellfish wastes:A potential additive for detergent and antioxidant synthesis[J].Bioprocess and biosystems engineering,2013,36(7):873-883.
[36] JOO H S,CHANG C S.Oxidant and SDS-stable alkaline protease from a halo-tolerant Bacillus clausii I-52:Enhanced production and simple purification[J].Journal of applied microbiology,2005,98(2):491-497.
[37] 郭成栓,崔堂兵,郭勇.嗜堿芽孢桿菌產堿性纖維素酶研究概況[J].氨基酸和生物資源,2007,29(1):35-38.
[38] HORIKOSHI K.Introduction and history of alkaliphiles[M]//HORIKOSHI K.Extremophiles handbook.Tokyo:Japan,2011:22-24.
[39] DAS P,BEHERA B K,MEENA D K,et al.Salt stress tolerant genes in halophilic and halotolerant bacteria:Salt stress adaptation and osmoprotection[J].International journal of current microbiology and applied sciences,2005,4(1):642-658.
[40] 林玲.內生芽孢桿菌及抗菌物質的鑒定和對作物黃萎病的防治效果[D].南京:南京農業大學,2009.
[41] 張文飛.蘇云金芽孢桿菌分離與新型cry基因資源的挖掘[D].南寧:廣西大學,2011.
[42] JENKINS J N,PARROTT W L,MCCARTY J C,et al.Field test of transgenic cottons containing a Bacillus thuringiensis gene[J].Technical bulletin-mississippi agricultural and forestry experiment station (USA),1991,174(9):174-176.
[43] 朱燕云,孔祥平,吳娥嬌,等.耐高鹽枯草芽孢桿菌XP合成球形納米硒及其抑制草莓病原真菌生物活性[J].生物工程學報,2021,37(8):2825-2835.
[44] 賈天梅.產耐熱型堿性蛋白酶菌株篩選、發酵條件優化及其酶學性質研究[D].杭州:浙江工商大學,2018.
[45] KIM W,CHOI K,KIM Y,et al.Purification and characterization of a fibrinolytic enzyme produced from Bacillus sp.Strain CK 11-4 screened from Chungkook-Jang[J].Applied and environmental microbiology,1996,62(7):2482-2488.
[46] 楊麗媛.堿性蛋白酶的分離純化及其酶學特性的研究[D].長春:東北師范大學,2015.
[47] 姚剛,程建軍,孫鵬,等.枯草芽孢桿菌發酵產堿性蛋白酶的研究[J].食品科學,2009,30(23):347-351.
[48] ZGREN T,PINAR O,BOZDAGˇ G,et al.Assessment of poly(3-hydroxybutyrate) synthesis from a novel obligate alkaliphilic Bacillus marmarensis and generation of its composite scaffold via electrospinning[J].International journal of biological macromolecules,2018,119:982-991.
猜你喜歡
西北民族大學學報(自然科學版)(2021年4期)2021-12-29 02:54:24
數學小靈通(1-2年級)(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
小聰仔(科普版)(2020年12期)2021-01-18 09:16:52
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
學生天地(2019年32期)2019-08-25 08:55:22
中學生數理化·七年級數學人教版(2019年4期)2019-05-20 10:06:32
中學生數理化·七年級數學人教版(2018年6期)2018-06-26 08:36:06
小天使·一年級語數英綜合(2017年11期)2017-12-05 18:49:56
初中生世界·七年級(2017年9期)2017-10-13 22:27:46
