熱浪對植物光合作用和水分傳輸與利用的影響研究進展
2022-05-17 23:00:51余海霞曲魯平劉南邵長亮董剛姜世成王拓范輝華胡亞林
安徽農業科學 2022年9期
余海霞 曲魯平 劉南 邵長亮 董剛 姜世成 王拓 范輝華 胡亞林
摘要 熱浪是指持續一段時間的極端高溫氣候事件,其發生的頻率和強度逐年增加。熱浪持續時間雖相對較短,但會顯著抑制植物的生長發育,甚至導致植物死亡,特別是熱浪末期的高溫與干旱的復合脅迫會嚴重抑制植物光合作用,造成水力障礙,加速植物死亡過程并增加其死亡率。其中,對植物的光合作用和水分傳輸與利用過程的研究是揭示熱浪導致植物死亡機制的關鍵,不同植物應對熱浪階段高溫干旱復合脅迫時會表現出不同的響應策略。目前,隨著全球氣候變化對陸地生態系統的影響加劇,有關熱浪這種極端高溫干旱氣候事件對植物光合作用和水分傳輸與利用的影響研究已成為生態學和植物學領域研究的熱點。對高溫和干旱脅迫對植物,特別是林木光合系統、水力傳輸系統、水分利用效率和生長發育等影響的研究結果進行總結歸納,探討了植物光合作用和蒸騰耗水對熱浪下的高溫與干旱脅迫的響應策略及其適應機制。同時,提出了已有研究存在的問題并對未來研究進行了展望,以期豐富植物光合作用和水分傳輸與利用等生理特征對極端氣候事件的響應理論。
關鍵詞 極端氣候;光合作用;水力結構;熱脅迫;水分脅迫;水分利用效率
中圖分類號 Q 948? 文獻標識碼 A? 文章編號 0517-6611(2022)09-0004-05
doi:10.3969/j.issn.0517-6611.2022.09.002
開放科學(資源服務)標識碼(OSID):
Research Progress of Heat Waves Influences on Plant Photosynthesis and Water Transport and Utilization
YU Hai-xia,QU Lu-ping,LIU Nan et al
(College of Forestry,Fujian Agricultural and Forestry University,Fuzhou,Fujian 350002)
Abstract Heat wave,defined as an extreme weather event with prolonged period of high temperature.And the frequency and intensity of heat waves have substantially increased and widely concerned continuously.Despite of a short duration,heat waves can significantly inhibit the growth and healthy of plant,even cause plant death.Especially the combination of high temperature and drought during the late stage of heat waves will severely inhibit? photosynthesis,cause hydraulic failure,finally accelerate plant death process and increase death rate.Understanding the plant photosynthesis and water transport and utilization process response to heat wave stress is the key to reveal the mechanism of how heat wave leading plant death,and different plant species exist divergence response strategies to the combined stress of high temperature and drought during heat waves.At present,as the impact of global climate change on terrestrial ecosystems intensified,studies on the effects of heat waves such as extreme high temperature and arid climate events on plant photosynthesis,water transport and utilization have become a hot topic in the field of ecology and botany.This study summarized the findings of coupled effects of high temperature and drought stress on plant photosynthetic and hydraulic transmission systems,the water use efficiency and plant growth etc.,and discussed the response strategies and adaptive mechanisms of plant photosynthesis and water transpiration to high temperature and drought stress under heat waves.Meanwhile,the problems of existing studies and prospects of future research are put forward. In order to enrich the mechanism and theory explanation of plant photosynthesis,water transport and utilization and other physiological characteristics to extreme climatic events.
Key words Extreme weather;Photosynthesis;Hydraulic structure;Thermal stress;Water stress;Water use efficiency
全球氣候變化不僅包括持續較長時間的氣候變動(如全球變暖、降水格局改變等),還包括氣候的短暫劇烈波動,即極端氣候的發生[1]。熱浪(heat wave)是指一段持續性(幾天)的高溫天氣過程,是一種典型的極端氣候事件,正受到科學界的廣泛關注。與全球變暖的緩慢增溫對比,熱浪發生時間雖短,但其伴隨的驟然高溫和干旱脅迫會對自然生態系統,特別是植物的生長發育造成顯著影響[2-3]。自20世紀中葉以來,全球陸地范圍內超過73%區域內熱浪發生的頻率和強度顯著上升,陸地生態系統受到極端氣候事件熱浪侵襲的可能性大大增加[4]。目前,氣候變化研究的焦點已從全球變暖和降水格局改變對植物生長發育的影響逐漸向極端氣候事件,特別是熱浪對植物生長發育的影響研究聚焦。
植物是自然和半自然生態系統的主體,熱浪對生態系統功能的影響的實質是對生產者(植物)的影響[5-6]。熱浪伴隨的高溫脅迫會直接灼傷植物,抑制植物光合作用,導致植物枯萎死亡[7]。 此外,高溫條件下植物的高耗水極易造成水分脅迫的發生,即熱浪末期植物面臨的是高溫與干旱的復合脅迫[8]。其中,干旱脅迫所導致的碳饑餓和水力障礙均會導致植物死亡[9],而與其復合的高溫脅迫則會增強熱浪脅迫強度和危害性,這種高溫與干旱的雙重脅迫的耦合作用會加速植物的死亡過程,并會顯著影響植物生長發育的過程和整個生態系統的物質生產與能量轉換[10-11]。綜上,更好的理解和闡釋熱浪過程中的多重脅迫對植物光合作用和水力傳輸的影響,明確植物應對熱浪脅迫的抗逆機理是當前全球氣候變化研究中的重大科學挑戰和熱點問題。該研究系統總結了當前國內外有關熱浪及其對植物光合作用與水分傳輸過程影響的相關研究,由此提出了熱浪相關研究中存在的問題,并對未來熱浪研究中的難點和重點問題進行了展望和研究方法上的建議,以期豐富全球氣候變化,特別是極端氣候事件下的植物抗逆生理理論。
1 熱浪是高溫與干旱主導的復雜脅迫
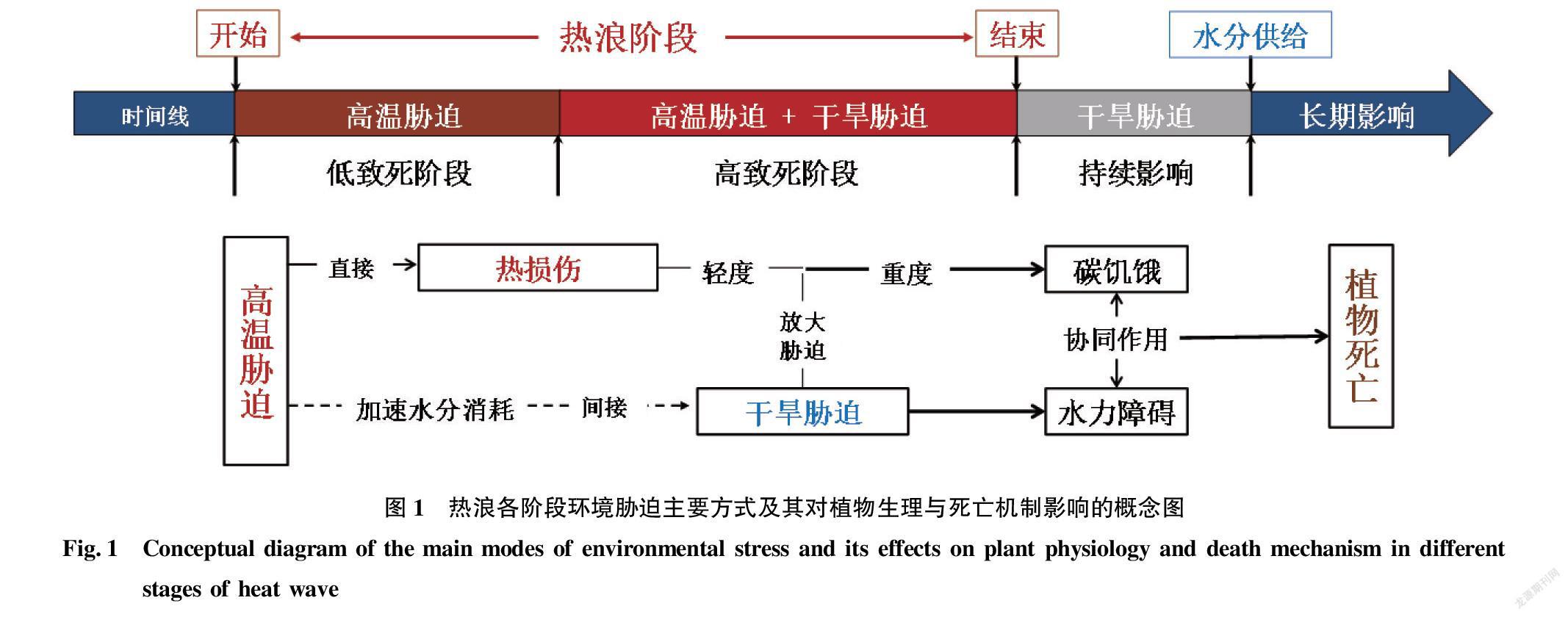
氣象學上將熱浪定義為持續長時間保持的過度炎熱,即高溫。植物抗逆生理學研究表明,高溫脅迫并不是熱浪影響植物生長發育的唯一因素,植物對于極端高溫的應激反應僅是該領域研究的一部分(圖1)。熱浪階段除溫度變化外,還伴隨著強光照(+69%)、低降水(-78%)和高水汽壓虧缺(VPD)(+111%),高溫下生態系統的高蒸發散和強耗水與低降水補給之間的矛盾會導致熱浪末期發生干旱脅迫[8]。光合作用是植物重要的生理過程,該過程對環境的變化十分敏感,熱浪階段光照、降水、風速及VPD的變化均會影響植物氣孔開閉、蒸騰、光電子傳遞等與光合作用的相關過程[12-14],降低光合作用,提高呼吸作用,從而加速非結構性碳水化合物的消耗,引起碳收支失衡,導致碳饑餓的發生。此外,干旱脅迫下的水分虧缺對植物水分傳輸功能的影響也是熱浪影響植物生長的關鍵因素[10],如干旱脅迫下木本植物發生的木質部栓塞現象會引發水力傳輸障礙[6],木質部栓塞不僅會進一步抑制植物光合作用,且隨著栓塞程度的增加,當超過一定閾值時嚴重阻礙土壤—植物連續體的水分傳輸,最終導致植物的死亡。因此,熱浪對植物生長發育影響的研究實質上需要關注的是高溫與干旱主導的變化中的復雜環境脅迫。
2 熱浪對植物光合作用的影響
光合作用是植物體內最重要的生命活動過程,對熱浪脅迫的響應十分敏感。首先,光合作用是植物對驟然增溫反應最為敏感的生理過程,極端高溫會顯著影響光合作用酶的活性,“光”(電子傳遞)和“暗”(卡爾文循環/CO2固定)反應都有熱敏感性,特別是光反應中的PSII和暗反應中的Rubisco活化酶(核酮糖二磷酸縮化酶)。極端高溫誘導的葉片葉綠體PSII損傷可預測不同物種如何應對極端天氣事件的機制特征。Guha等[14]對南方紅櫟(Quercus falcata Michx.)、舒瑪櫟(Q.shumardii Buckl.)、郁金香-白楊(Liriodendron tulipifera L.)和東方白松(Pinus strobus L.)模擬人工熱浪試驗發現非光化學猝滅在熱浪周期中顯著增加,而在最敏感的樹種東方白松和郁金香-白楊中PSII的瞬態和慢性損傷跡象明顯;與其他樹種相比,櫟類的耐高溫性更強,其PSII的光損傷閾值更高,光抑制的夜間恢復速度更快,熱引起的冠層壞死最小。Drake等[12]對桉樹(Eucalyptus parramattensis)進行原位模擬熱浪試驗,發現熱浪階段正午時分桉樹凈光合速率降低到接近于0。熱脅迫還可通過降低Rubisco酶活性和降低CO2的溶解度來影響光合CO2同化作用[15]。如棉花、小麥和玉米在熱浪脅迫下的Rubisco 酶活性受到抑制,隨著溫度的升高其光合速率因Rubisco酶活性的降低而下降[16-17]。
由于不同植物的光反應途徑和生理生態特性具有差異,故不同植物光合作用對高溫的耐受閾值不同。C3植物對高溫的適應性略遜于C4植物。研究表明,C3植物棉花和小麥的光合速率分別在葉片溫度高于35 ℃和30 ℃時受到抑制,而當葉溫增至42.5 ℃和40.0 ℃時,光合速率完全受到抑制[16]。C4植物玉米光合作用的熱敏感性研究表明,當葉溫超過37.5 ℃時光合作用才會受到抑制,且在45 ℃的高溫下,玉米仍能保持50%的最大光合速率[17]。植物對熱浪的耐受性的差異同樣表現在其他生態系統中,其中,草地生態系統熱浪的研究表明,C3植物加拿大一枝黃花(Solidago canadensis)較C4植物須芒草(Andropogon gerardii)存在更好的熱耐受性[18];而森林生態系統中,不同類型的苗木在熱浪影響下,其成活率也不同[19]。
相較于高溫脅迫對植物光合作用的直接影響,干旱脅迫不會直接損害植物進行光合作用的生物化學和光化學系統,而是通過增加進入葉綠體的CO2的擴散阻力影響光合作用[20]。在干旱脅迫下等水調節的植物通常表現為逐漸關閉氣孔以保持細胞內的水分,氣孔導度的下降直接導致葉綠體內CO2同化作用降低,光合速率降低,從而抑制葉片生長和植物生長[21-22]。氣孔關閉導致的低水勢還會導致植物細胞脫水、木質部氣蝕與栓塞、水力障礙發生,引發植物死亡[23]。Signarbieux等[24]在模擬干旱對雜類、豆科和禾本科植物的影響研究中發現禾本科植物梯牧草(Phleum pratense)和多花黑麥草(Lolium multiflorum)中氣孔導度和光合速率平行下降,而雜類植物鈍葉草(Rumex obtusifolius)和豆科植物白三葉(Trifolium repens)中氣孔導度和光合速率沒有表現出顯著差異,白草蒿(Artemisia sieberi alba)的光合速率在干旱脅迫10 d后顯著降低了,且氣孔導度、蒸騰速率、葉綠素熒光參數等光合屬性顯著降低[25]。
此外,熱浪階段的干旱脅迫會加劇熱脅迫對植物的影響,干旱脅迫和高溫迫并不是簡單的累加,而是交互影響,耦合作用加速植物的死亡[26]。若熱浪伴隨的高溫超過了植物的熱耐受閾值,會直接導致“熱損傷”或植株死亡。生理學上,植物為了對抗長期的高溫,最有效的生理適應手段是增加氣孔導度,加速葉片的蒸騰速率,通過增加潛熱的冷卻方式降低葉片溫度從而對抗熱脅迫[12]。但這種生理適應手段可行的前提是植物所處的生態系統水分儲備充足,而在熱浪后期的干旱脅迫下,較低的土壤體積含水量和高的VPD均表明生態系統無法提供足夠的水分以完成植物的這種熱適應機制。此時,高溫干旱對光合作用的負面影響疊加,植物氣孔關閉,光合作用降低,降溫調節失效,首先出現葉綠素降解、Rubisco酶活性降低和光系統(PSII)受損[19],進一步降低光合效率,植物異化作用高于同化作用,急速“碳饑餓”現象發生[27],最終引發氨中毒、生物膜損傷和蛋白質變性等一系列更為嚴重的影響和植物的死亡[28]。研究表明,楊樹凈光合速率(Pn)在高溫、干旱、高溫干旱復合脅迫下分別下降53%、50%及77% [29]。綜上,熱浪對植物光合作用的影響不是高溫和干旱脅迫的獨立存在或簡單疊加作用,而是兩者的交互耦合作用。因高溫與干旱的耦合脅迫,熱浪末期是植物死亡發生的高峰期,干旱脅迫是熱浪造成植物光合作用受抑制和植物死亡的主因,而同時伴隨的高溫脅迫又會加速植物的死亡進度,并放大熱浪對植物的影響程度[13]。
安徽農業科學2022年
3 熱浪對植物水分傳輸與利用的影響
水力失衡和水力傳輸障礙是熱浪脅迫導致植物死亡的機制之一,該機制與干旱脅迫的致死機制相似,表現為植物水分傳輸障礙的發生使葉片失水死亡[30]。植物對環境中的水分狀況的敏感性很高,但植物通常并不直接感知土壤含水量,而是通過土壤和根系之間的相互作用影響水分脅迫的水平和動態[31]。持續的水分虧缺會引發水通道蛋白活性的調節作用,植物根系水力傳導先升高后降低[32],根內皮層或者外皮層的栓化增加,根系水力傳導會進一步降低,由此減少了進入植物的水流量,也防止了水分從植物流失到干燥的土壤中[33]。而在土壤-植物-大氣連續體中,植物會通過氣孔調節來緩解蒸騰速率和水分散失,使葉片水勢保持在閾值以上。但隨著干旱加劇,植物水勢下降到閾值以下時,木質部就有發生空穴和栓塞的風險,從而阻礙水分傳輸。植物耐旱性研究表明,等水調節植物可通過調整水力結構來改變導水阻力和水分需求,進一步影響水分傳輸和水分平衡,從而適應干旱脅迫[6]。偏向非等水調節策略的植物在熱浪階段更易發生失水現象,因其氣孔在水分脅迫條件下依然保持較高的張開程度來維持最大的光合速率,而隨著生態系統水分耗盡,木質部導管水勢的降低,將面臨更高的導管栓塞化和水力障礙的風險[22]。另外,干旱脅迫時間越長,植物體內水力傳輸會通過降低木質部栓塞化來中斷植物莖和葉部的汁液流動,由此降低蒸騰速率和增強植物的水分利用效率[6]。
高溫脅迫通常會對植物的水分利用效率產生負面影響,主要表現為高溫降低了植物的光合速率[34]。Duan等[35]研究發現櫻桃番茄(Solanum lycopersicum)在干旱條件下的水分利用效率更高,是由于水分虧缺顯著降低了氣孔導度,其蒸騰強度和光合強度降低幅度存在不一致性。研究表明,同一植物的水分利用效率在熱浪的初期和末期具有差異,且不同植物的水分利用效率對熱浪的響應也存在特異性,如熱浪脅迫下刺槐 (Robinia pseudoacacia L.)和南方紅橡木(Quercus.falcata)的水分利用效率表現為先升高再下降,而杉木(Cunninghamia lanceolate)和鵝掌楸(Liriodendron tulipifera)的水分利用效率則是持續下降[11,36]。不同植物水分傳輸和利用效率對熱浪的不同響應反映了不同物種的耐熱性水平,其潛在機制仍需進一步研究和闡明。
4 熱浪對植物生長的影響
熱浪伴隨的高溫和干旱脅迫對植物光合系統的抑制與水力傳輸障礙的最終表現為對植物生長速率、葉片形態、營養物質分配和生物量的影響[37]。植物對熱浪脅迫的短期應激性反應并不能代表熱浪事件對植物影響的全過程,無論是熱脅迫造成的植物光合組織器官的熱損傷,還是干旱脅迫造成的木質部栓塞等水力結構障礙,植物的恢復均需要一定的時間,且并非能夠完全恢復至脅迫前的水平[7]。如亞熱帶地區的熱浪會導致杉木(Cunninghamia lanceolata)導水率顯著降低,并出現木質部栓塞現象且難以恢復;木荷(Schima superba)雖然木質部栓塞現象不明顯,但大量光合器官(葉片)受到熱損傷脫落,但熱浪會促進其地下生物量的增長,增強木荷的根冠比[6]。草本植物白草蒿(Artemisia sieberi alba)在熱浪影響下同樣會降低葉面積和植株高度等[25]。
此外,熱浪脅迫還改變了植物器官間的物質分配,如不同基因型玉米經過高溫處理后降低了穗的膨脹,穗部重量減少、果穗和果柄中纖維素組分的糖含量降低,而葉和莖的重量略有增加[38]。高溫脅迫下羊茅(Festuca ovina L.)的凈光合速率下降的同時,其總氮、硝酸鹽、可溶性蛋白和總游離氨基酸(20種氨基酸)的積累量均下降[39]。熱浪脅迫還將改變植物種子的質量,進一步影響植物的生殖過程,延長其影響時間。如熱浪脅迫會改變豆類籽粒中的氮含量、降低小麥和玉米籽粒中的淀粉顆粒、蛋白質和油含量,抑制小麥籽粒生長,限制作物產量,累積變化最終造成植物生長和生產力下降[40-41]。高溫干旱復合脅迫顯著抑制了不同基因型扁豆(Lens culinaris Medikus)植株的生長,縮短了植物的生殖期,降低了種子質量[42]。
5 展望
在全球氣候變化持續發生和極端氣候頻發的背景下,未來極端氣候事件——熱浪發生的頻率、持續的時間、覆蓋地域均會增加。因此,熱浪等極端氣候事件對植物的影響,特別是對植物光合作用和水力傳輸等重要生命過程的影響研究需要進一步深入和加強[43]。雖然,目前國內外關于熱浪對植物光合作用、水力傳輸與利用、植株生長和生產力影響的研究取得了一定進展,但仍存在一定的問題和一些新的挑戰,是熱浪研究中亟待規范和解決的。該研究基于對當前國內外有關對熱浪的研究成果的歸納總結,對未來有關極端氣候事件熱浪對植物影響的研究做出以下展望,建議明確和加強以下幾個方面的重點與難點問題的研究。
5.1 熱浪定義的統一與規范
目前國際上對于熱浪缺乏統一而明確的定義。日均溫、最高溫度、最低溫度、相對溫度或一些其他指數[(如EHF(excess heat factor)、SHI(standardized heat index)]均可用來定義高溫;此外,對于熱浪標準的持續時間同樣缺乏共識[44]。相較于全球變暖研究所需要關心的單一維度變量(即增溫幅度),熱浪的增溫幅度、持續長短、發生時間和頻率變化均直接關系熱浪對植物產生脅迫的強度和影響。熱浪自身的復雜屬性直接關系其效應強度。由于缺乏統一規范的定義,導致熱浪研究中所關注的熱浪強度差異性無法在同一尺度上進行比較。因此,在未來研究中首先要將熱浪事件的內涵統一與規范后,在相對標準的試驗條件下進行熱浪模擬和控制試驗。
5.2 熱浪是高溫與干旱主導的復合脅迫
早期研究中將高溫脅迫認為是熱浪影響植物生長的唯一因素,有關熱浪的研究被等同于植物高溫耐受性的研究。實際上,熱浪脅迫導致植物死亡的關鍵因素是后期的高溫與干旱主導的復合脅迫。因此,研究植物對熱浪脅迫的響應策略不能只考慮耐熱性,植物在干旱脅迫下的耐熱性才是研究的關鍵。但是,目前有關熱浪對植物影響的研究多注重高溫和干旱的獨立效應與簡單疊加效應,故在未來研究中應該注重高溫和干旱的耦合脅迫的復合作用對植物抗逆性機制的研究。
5.3 缺乏熱浪后植物恢復過程和能力的研究
相較于全球變暖,熱浪持續時間雖短,但其對植物生長的影響并沒有隨著脅迫的結束而終止,熱浪后植物的恢復過程和恢復能力是揭示熱浪對植物影響的關鍵。由于不同植物對熱浪耐受的特異性,熱浪對植物的影響會從種群水平延展至群落水平(如群落豐富度、多樣性及均勻度的改變)[45],最終表現為生態系統水分利用效率和碳循環的變化。因此,有關熱浪對植物生長和生理特性影響的研究,應該分為短時效應(rapid effect)、滯后效應(post effect)、遺留效應(legacy effect)3個階段或過程去深入研究,以此更加準確地量化熱浪對植物生長和生理特性的短、中、長期影響[46]。其中,短時效應表征植物對熱浪的耐受性,滯后效應表征熱浪后植物的恢復能力,遺留效應表征熱浪脅迫對植物的永久損傷,3個過程存在聯系也相互獨立。然而,目前國內外對熱浪的研究多集中在短期內的影響,故在未來研究中還應該注重熱浪滯后效應和遺留效應階段的研究。
5.4 熱浪研究方法的改進
熱浪研究方法目前主要分為自然熱浪研究和人工模擬熱浪研究。自然熱浪研究方法的優點是氣象條件和植被生長均處于自然狀態,獲得的數據更具真實性。但該方法缺乏明確的對照,作為對照的年或時間斷面的氣象水平、植物和生態系統狀態自身存在差異,會造成對熱浪影響的錯誤估計。人工模擬熱浪研究是指使用加熱裝置將一段時間的高溫施加到人工種植或者自然生長的個體、群落或者生態系統中的方法。該方法可彌補自然熱浪發生的時間和地域存在的隨機性和不確定性,并可設置響應的對照處理試驗。早期模擬熱浪的研究多是在人工氣候室內完成,以盆栽試驗為主。首先,人工氣候室模擬的熱浪一般為持續高溫,而非自然熱浪狀態下的不對稱增溫。其次,盆栽植物于自然群落中的生長狀態迥異。如林木幼苗在森林生態系統中通常處于下層,光照較弱,空氣濕度較高。此外,群落中生物間的相互作用有助于增強植物個體對熱浪的耐受性[3],而盆栽試驗忽略了這些問題,導致試驗結果與實際情況之間具有差異性。因而,野外原位模擬熱浪試驗可能是研究熱浪對植物影響較為理想的方法,但這類模擬試驗很難實現。目前用于野外原位熱浪模擬的方法耗資巨大,難以維護,便捷性差,且存在安全隱患[12,46]。因此,對于熱浪研究方法的改進也是未來熱浪對植物影響研究中亟待解決的問題。
5.5 關注熱浪發生時間和頻率的變化
有關熱浪變化趨勢的研究表明,未來熱浪發生的時間將明顯提前,且發生頻率增加,將會導致高頻連續多次熱浪事件的發生。不同時期的熱浪發生時伴隨的熱脅迫和干旱脅迫的強度必定不同,熱浪的提前發生勢必會打破植物的節律性適應,而高頻率的熱浪事件意味著植物獲得恢復時間更加不確定,且多次疊加的熱浪對植物的影響會存在累加效應。因此,植物對不同模式熱浪(不同頻率、不同持續時間等)的響應存在差異,在未來研究中應注重在不同模式的熱浪事件下研究植物對短期氣候事件的響應策略和適應機理。
綜上所述,熱浪作為極端氣候的代表性事件,對植物的影響將直接導致全球大多數生態系統的功能失衡,從而對地球生態系統產生災難性的后果。而對于這種災難后果的預測,其根本是理解熱浪對植物個體的影響的機理,才能延伸、擴展和探討更大尺度的問題。盡管熱浪對植物生長影響的研究是現階段植物學及生態學的熱點問題,但是目前熱浪研究的數量匱乏,研究方法存在爭議,對植物的影響機理的解釋仍不徹底和不全面。因此,在未來氣候變化的研究中,應當加強對熱浪等極端氣候的關注,以更好地理解全球氣候變化對植物生理、生態系統功能的影響并預測其變化走向,為管理和決策部門提供更加科學的參考依據。
參考文獻
[1]
IPCC.Climate change 2013:The Physical Science Basis.Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change[R].Cambridge:Cambridge University Press,2013.
[2] PIAO S L,ZHANG X P,CHEN A P,et al.The impacts of climate extremes on the terrestrial carbon cycle:A review[J].Sci China Earth Sci,2019,62(10):1551-1563.
[3] QU L P,DONG G,DE BOECK H J,et al.Joint forcing by heat waves and mowing poses a threat to grassland ecosystems:Evidence from a manipulative experiment[J].Land Degrad Dev,2020,31(7):785-800.
[4] YUAN W P,CAI W W,CHEN Y,et al.Severe summer heatwave and drought strongly reduced carbon uptake in Southern China[J].Sci Rep,2016,6(6):1-12.
[5] REICHSTEIN M,BAHN M,CIAIS P,et al.Climate extremes and the carbon cycle[J].Nature,2013,500(7462):287-295.
[6] QU L P,DE BOECK H J,FAN H H,et al.Diverging responses of two subtropical tree species (Schima superba and Cunninghamia lanceolata) to heat waves[J].Forests,2020,11(5):513-533.
[7] RUEHR N K,GROTE R,MAYR S,et al.Beyond the extreme:Recovery of carbon and water relations in woody plants following heat and drought stress[J].Tree Physiol,2019,39(8):1285-1299.
[8] DE BOECK H J,DREESEN F E,JANSSENS I A,et al.Climatic characteristics of heat waves and their simulation in plant experiments[J].Glob Change Biol,2010,16(7):1992-2000.
[9] MCDOWELL N G,GROSSIORD C,ADAMS H D,et al.Mechanisms of a coniferous woodland persistence under drought and heat[J].Environ Res Lett,2019,14(4):1-14.
[10] DE BOECK H J,BASSIN S,VERLINDEN M,et al.Simulated heat waves affected alpine grassland only in combination with drought[J].New Phytol,2016,209(2):531-541.
[11] RUEHR N K,GAST A,WEBER C,et al.Water availability as dominant control of heat stress responses in two contrasting tree species[J].Tree Physiol,2016,36(2):164-178.
[12] DRAKE J E,TJOELKER M G,VRHAMMAR A,et al.Trees tolerate an extreme heatwave via sustained transpirational cooling and increased leaf thermal tolerance[J].Glob Chang Biol,2018,24(6):2390-2402.
[13] DE BOECK H J,VAN DE VELDE H,DE GROOTE T,et al.Ideas and perspectives:Heat stress:More than hot air[J].Biogeosciences,2016,13(20):5821-5825.
[14] GUHA A,HAN J M,CUMMINGS C,et al.Differential ecophysiological responses and resilience to heat wave events in four co-occurring temperate tree species[J].Environ Res Lett,2018,13(6):1-13.
[15] CRAFTS-BRANDNER S J,SALVUCCI M E.Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2[J].PNAS,2000,97(24):13430-13435.
[16] LAW R D,CRAFTS-BRANDNER S J.Inhibition and acclimation of photosynthesis to heat stress is closely correlated with activation of ribulose-1,5-bisphosphate carboxylase/oxygenase[J].Plant Physiol,1999,120(1):173-182.
[17] CRAFTS-BRANDNER S J,SALVUCCI M E.Sensitivity of photosynthesis in a C4 plant,maize,to heat stress[J].Plant Physiol,2002,129(4):1773-1780.
[18] WANG D,HECKATHORN S A,MAINALI K,et al.Timing effects of heat-stress on plant ecophysiological characteristics and growth[J].Front in Plant Sci,2016,7:1-11.
[19] TESKEY R,WERTIN T,BAUWERAERTS I,et al.Responses of tree species to heat waves and extreme heat events[J].Plant Cell Environ,2015,38(9):1699-1712.
[20] ZHAO J B,HARTMANN H,TRUMBORE S,et al.High temperature causes negative whole-plant carbon balance under mild drought[J].New Phytol,2013,200(2):330-339.
[21] WOODRUFF D R,MEINZER F C,MARIAS D E,et al.Linking nonstructural carbohydrate dynamics to gas exchange and leaf hydraulic behavior in Pinus edulis and Juniperus monosperma[J].New Phytol,2015,206(1):411-421.
[22] HARTMANN H,LINK R M,SCHULDT B.A whole-plant perspective of isohydry:Stem-level support for leaf-level plant water regulation[J].Tree Physiol,2021,41(6):901-905.
[23] CHAVES M M,MAROCO J P,PEREIRA J S.Understanding plant responses to drought-from genes to the whole plant[J].Funct Plant Biol,2003,30(3):239-264.
[24] SIGNARBIEUX C,FELLER U.Effects of an extended drought period on physiological properties of grassland species in the field[J].J Plant Res,2012,125(2):251-261.
[25] ALHAITHLOUL H A S.Impact of combined heat and drought stress on the potential growth responses of the desert grass Artemisia sieberi alba:Relation to biochemical and molecular adaptation[J].Plants (Basel),2019,8(10):416-441.
[26] DE BOECK H J,DREESEN F E,JANSSENS I A,et al.Whole-system responses of experimental plant communities to climate extremes imposed in different seasons[J].New Phytol,2011,189(3):806-817.
[27] MCDOWELL N G,SEVANTO S.The mechanisms of carbon starvation:How,when,or does it even occur at all?[J].New Phytol,2010,186(2):264-266.
[28] KUCZYN′SKA-WIS′NIK D,MATUSZEWSKA E,LASKOWSKA E.Escherichia coli heat-shock proteins IbpA and IbpB affect biofilm formation by influencing the level of extracellular indole[J].Microbiology,2010,156(Pt1):148-157.
[29] 張燕紅,吳永波,劉璇,等.高溫和干旱脅迫對楊樹幼苗光合性能和抗氧化酶系統的影響[J].東北林業大學學報,2017,45(11):32-38.
[30] MCDOWELL N,POCKMAN W T,ALLEN C D,et al.Mechanisms of plant survival and mortality during drought:Why do some plants survive while others succumb to drought?[J].New Phytol,2008,178(4):719-739.
[31] LIPIEC J,DOUSSAN C,NOSALEWICZ A,et al.Effect of drought and heat stresses on plant growth and yield:A review[J].Int Agrophysics,2013,27(4):463-477.
[32] MAUREL C,SIMONNEAU T,SUTKA M.The significance of roots as hydraulic rheostats[J].J Exp Bot,2010,61(12):3191-3198.
[33] VANDELEUR R K,MAYO G,SHELDEN M C,et al.The role of plasma membrane intrinsic protein aquaporins in water transport through roots:Diurnal and drought stress responses reveal different strategies between isohydric and anisohydric cultivars of grapevine[J].Plant Physiol,2009,149(1):445-460.
[34] HAWORTH M,COSENTINO S L,MARINO G,et al.Physiological responses of Arundo donax ecotypes to drought:A common garden study[J].GCB Bioenergy,2017,9(1):132-143.
[35] DUAN H L,WU J P,HUANG G M,et al.Individual and interactive effects of drought and heat on leaf physiology of seedlings in an economically important crop[J].AoB PLANTS,2017,9(1):1-16.
[36] GUHA A,HAN J M,CUMMINGS C,et al.Differential ecophysiological responses and resilience to heat wave events in four co-occurring temperate tree species[J].Environ Res Lett,2018,13(6):1-14.
[37] BIRAMI B,GATTMANN M,HEYER A G,et al.Heat waves alter carbon allocation and increase mortality of Aleppo pine under dry conditions[J].Front in For Glob Change,2018,1:1-17.
[38] HOOVER D L,KNAPP A K,SMITH M D.Contrasting sensitivities of two dominant C4 grasses to heat waves and drought[J].Plant Ecol,2014,215(7):721-731.
[39] CUI L J,CAO R,LI J L,et al.High temperature effects on ammonium assimilation in leaves of two Festuca arundinacea cultivars with different heat susceptibility[J].Plant Growth Regul,2006,49(2/3):127-136.
[40] SEKHON H S,SINGH G,SHARMA P,et al.Water use efficiency under stress environments[M]//YADAV S S,REDDEN R.Climate change and management of cool season grain legume crops.Netherlands:Springer,2010:207-227.
[41] SEHGAL A,SITA K,KUMAR J,et al.Effects of drought,heat and their interaction on the growth,yield and photosynthetic function of lentil (Lens culinaris Medikus) genotypes varying in heat and drought sensitivity[J].Front Plant Sci,2017,8:1776-1998.
[42] AWASTHI R,KAUSHAL N,VADEZ V,et al.Individual and combined effects of transient drought and heat stress on carbon assimilation and seed filling in chickpea[J].Funct Plant Biol,2014,41(11):1148-1167.
[43] QU L P,CHEN J Q,DONG G,et al.Heat waves reduce ecosystem carbon sink strength in a Eurasian meadow steppe[J].Environ Res,2016,144:39-48.
[44] RAEI E,NIKOO M R,AGHAKOUCHAK A,et al.GHWR,a multi-method global heatwave and warm-spell record and toolbox[J].Sci Data,2018,5:1-15.
[45] KREYLING J,DENGLER J,WALTER J,et al.Species richness effects on grassland recovery from drought depend on community productivity in a multisite experiment[J].Ecol Lett,2017,20(11):1405-1413.
[46] QU L P,CHEN J Q,DONG G,et al.Heavy mowing enhances the effects of heat waves on grassland carbon and water fluxes[J].Sci Total Environ,2018,627:561-570.
