高寒半濕潤地區不同苜蓿品種越冬期根系生理及構型特征
2022-04-25 12:40:56孫萬斌馬暉玲周青平陳有軍張俊超劉文輝
草地學報 2022年4期
孫萬斌, 馬暉玲*, 周青平, 陳有軍, 張 偉, 張俊超, 劉文輝
(1.甘肅農業大學草業學院/草業生態系統教育部重點實驗室/甘肅省草業工程實驗室/中-美草地畜牧業可持續發展研究中心, 甘肅 蘭州 730070; 2.西南民族大學青藏高原研究院/四川若爾蓋高寒濕地生態系統國家野外科學觀測研究站/四川省發改委工程實驗室, 四川 成都 610041; 3.青海省青藏高原優良牧草種質資源利用重點實驗室青海大學畜牧獸醫科學院, 青海 西寧 810016)
青藏高原是我國重要的畜牧業生產基地,也是我國草地畜牧業的重要組成部分。近年來,牧區人口數量不斷增加,傳統放牧仍為當地主要的草地利用方式,加之全球氣候變化的影響,致使天然草地生產力降低和生態環境惡化,極大制約了該地區草地畜牧業的可持續發展[1]。建植優質高產的人工草地,利用現代化草牧業生產方式將是該地區畜牧業發展的必由之路[2]。苜蓿(MedicagosativaL.)是世界范圍內的一種品質高、適口性好的飼料作物[3],不僅具有優良的飼用功能,而且在現代草地農作制度中,起到促進農田生態系統物流及能流良性循環、保持耕地水土、提高后茬作物產量的重要生態作用。青藏高原地區冬、春季空氣干燥,風蝕作用強烈[4],嚴酷的自然環境威脅苜蓿越冬,制約著該地區優質苜蓿生產,因此,在該地區進行苜蓿越冬期根系生理及構型特征研究對高寒地區優質苜蓿品種的選擇和建植優質苜蓿人工飼草地具有重要意義。
青藏高原地區冬春季空氣干燥、氣溫低、紫外線強烈,導致植物體內活性氧積累,影響正常生理代謝[5]。低溫打破活性氧產生和清除之間的平衡,造成活性氧積累,損傷膜系統、蛋白質和核酸。活性氧積累是植物受到低溫損傷的重要原因之一[6],而酶類抗氧化系統是植物體內清除活性氧、維持植物細胞內活性氧平衡的重要調節機制[7];植物細胞內的丙二醛是脂質過氧化的產物,在一定程度上可以反映細胞膜脂過氧化的程度[8],累積的丙二醛也會加劇細胞膜脂的損傷;陶雅[9]、南麗麗[10]等人的研究認為可溶性糖含量與苜蓿的抗寒性有關,在抗寒性較強的苜蓿品種根頸中可溶性糖含量處于較高的水平。
苜蓿根系作為主要的越冬器官,可通過改變表型性狀和對養分的儲備影響紫花苜蓿的抗寒性[11]。劉志英等人[12]的研究認為根頸直徑和側根數是影響紫花苜蓿越冬率的正向貢獻因子,并且根系直徑等性狀并不是決定抗寒能力的主導因子,不同根系性狀的紫花苜蓿品種表現出對越冬率的差異化響應;劉香萍[13]認為抗寒和不抗寒品種根部組織結構中木質部分布上的差異與苜蓿的抗寒性存在一定的組織結構適應性關系;也有研究認為分枝型根系比單個直根更能忍受凍拔現象[14]。
在青藏高原海拔3 000 m以上地區,對第一年田間自然越冬苜蓿根部與根頸生理響應的研究鮮見報道。因此本研究在青藏高原海拔3 500 m半濕潤地區,在自然越冬的4個不同時間點,分別比較播種當年10個不同苜蓿品種根頸和根部抗氧化酶和滲透調節物質指標,并分析比較這些苜蓿品種的根系特征指標,研究這些指標與苜蓿越冬率的關系,以期為青藏高原高寒地區建植優質人工苜蓿草地和苜蓿新品種選育提供參考資料。
1 材料與方法
1.1 試驗地概況
于四川若爾蓋高寒濕地生態系統國家野外科學觀測研究站草種質資源圃內(32.83° N,102.58° E)開展試驗,試驗地海撥3 492 m,年均降水量為753 mm,80%降水集中于5-10月份;干雨季分明,雨熱同期;年平均氣溫約1.1℃,1月平均氣溫-10.3℃,7月平均氣溫10.9℃。該地氣候類型屬大陸性高原寒溫帶季風氣候;日照長,太陽輻射強烈。試驗地土壤為栗鈣土,pH值為5.85;土壤容重為1.02 g·cm-3;土壤有機質含量24.62 g·kg-1;全氮含量為 2.76 g·kg-1;全磷含量為0.94 g·kg-1。離地表10 cm,20 cm,30 cm處土壤溫度和土壤含水量利用預先置于試驗小區中的EM50土壤溫、濕度監測系統采集,每10 min采集1次溫濕度數據,土壤溫、濕度變化情況如圖1所示。
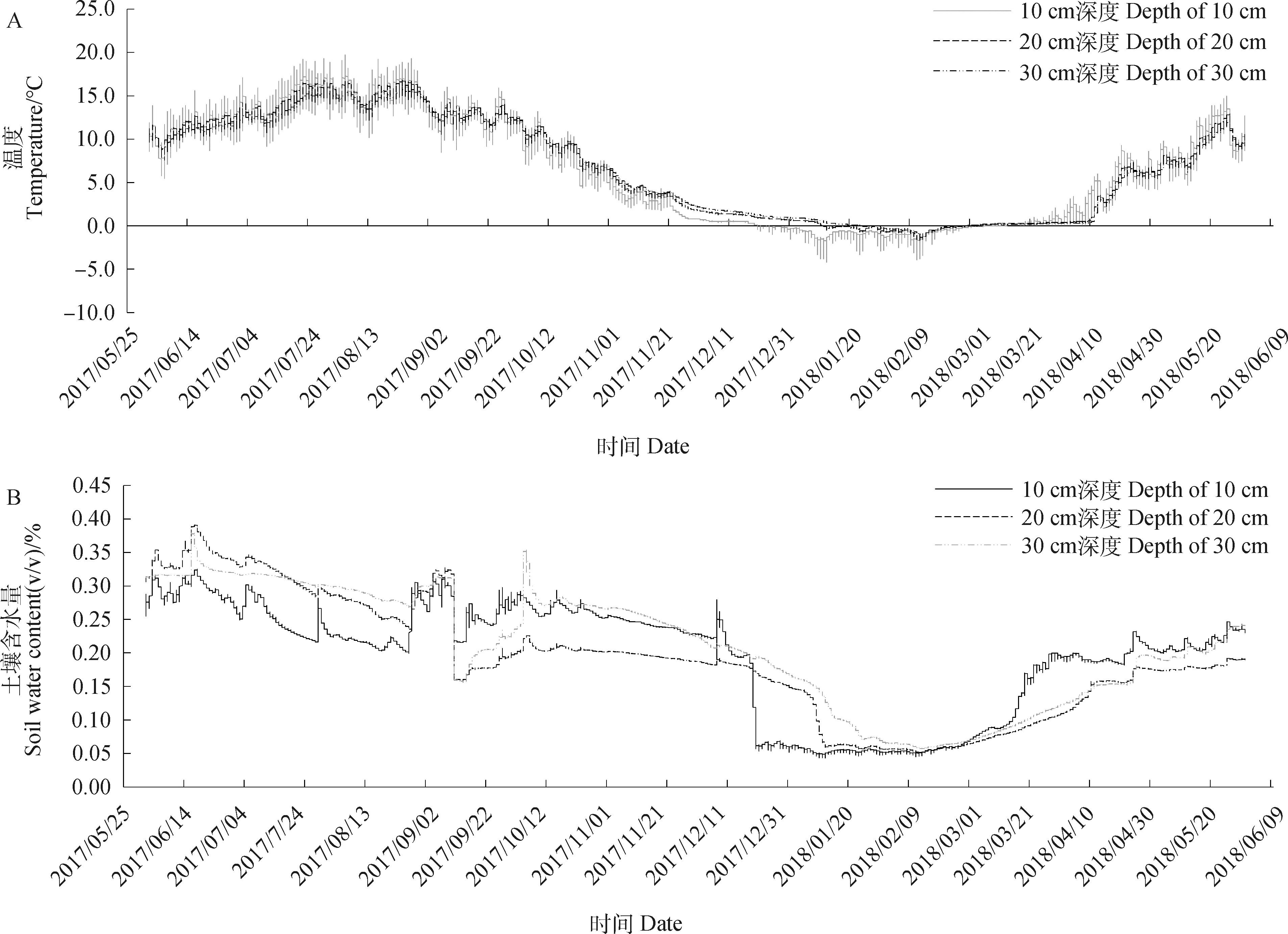
圖1 試驗地距離地表10 cm,20 cm,30 cm處土壤溫度和土壤含水量變化
1.2 試驗設計
將10個苜蓿品種(表1)于2017年6月2日種植于西南民族大學青藏高原基地牧草資源圃內,設3個重復,共30個小區,小區面積15 m2(3 m×5 m),采用隨機區組排列。播前翻耙平整后開溝,開溝深約2~3 cm;條播,行間距約30 cm,播種量為3 g·m-2。試驗期間各小區統一管理,人工除草3次。分別在2017年9月26日、11月16日和2018年1月15日、3月24日(分別記T1為抗寒鍛煉初期;T2抗寒鍛煉末期;T3土壤開始凍結;T4土壤解凍)取根及根頸組織鮮樣迅速投入液氮中并儲存至—80℃冰箱中保存,用于測定與抗寒性相關的生理生化指標。并于返青后2018年5月11日統計越冬率。且于越冬前,在上述每個小區內隨機選取30株苜蓿,挖取根系用于根系特征的測定。
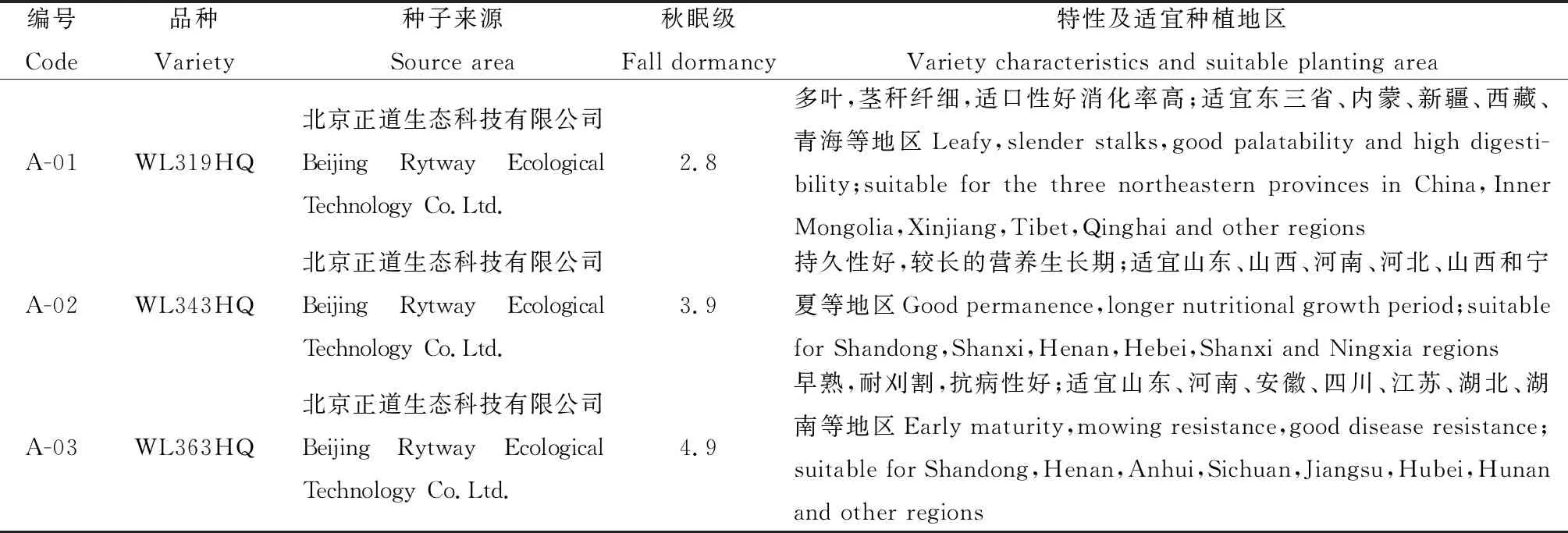
表1 供試品種

續表1
1.3 測定指標與方法
1.3.1越冬率(Overwintering rate)的測定 越冬率(%)=返青存活株數/越冬前存活株數×100[15]。
1.3.2生理生化指標測定 超氧化物歧化酶(Superoxide dismutase,SOD)、過氧化氫酶(Catalase,CAT)、過氧化物酶(Peroxidase,POD)活性和丙二醛(Malondialdehyde,MDA)、游離脯氨酸(Proline,Pro)、可溶性糖(Soluble sugar,SS)含量采用鄒琦[16]所述方法進行測定。
1.3.3根系形態指標的測定 用流水徹底清洗根系,然后將根系置于透明聚甲基丙烯酸根盤中,加蒸餾水使水位達1 cm高,用塑料鑷子撥動根系,使根系盡量避免交叉和重疊,然后利用Epson expression 12 000 xl掃描儀在600 DPI下獲取圖像,最后利用Win-RHIZO 2017a(Regent Instruments,Inc.,Quebec,Canada)軟件分析掃描的圖像,并得到所需根系數據。
1.3.4根系拓撲指數的測定 Fitter[17]和Bouma[18]等人提出了魚骨形分支(圖2B)和叉狀分支(圖2A)兩種極端根系拓撲類型,并使用根系拓撲指數(Topological index,TI)反映植物根系的不同分支模式。其中TI=lgA/lgM(M表示外部連接的數量,等于根中分生組織的數量;A表示根系從外部連接到最基礎連接的最長路徑上的連接數)。相關研究表明[19-20],大多數植物根系拓撲結構是在叉狀分支和魚骨形分支模式之間。當TI接近1時,根系呈現魚骨形分枝,當TI接近0.5時,根系呈現叉狀分枝。
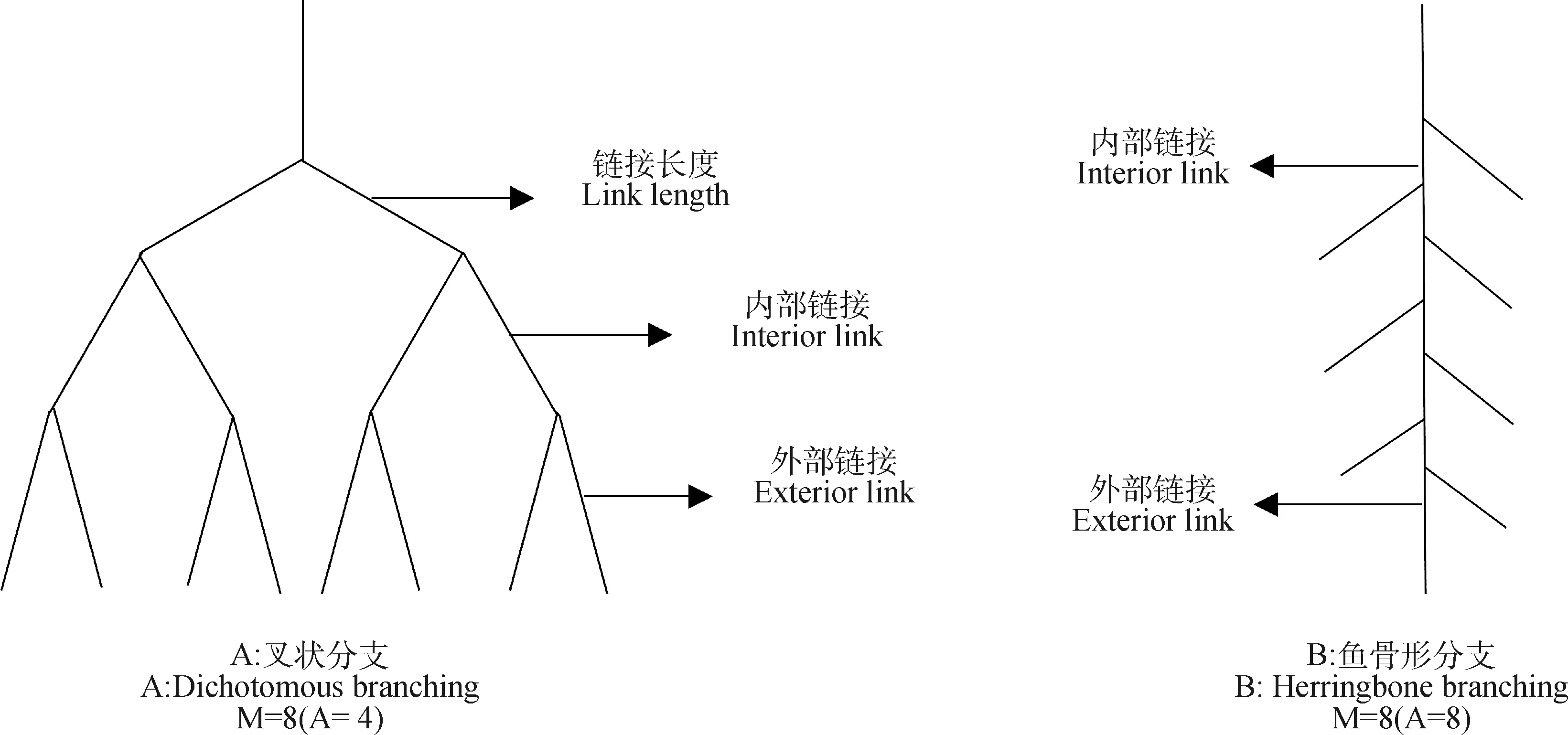
圖2 根系拓撲結構
1.4 數據統計分析方法
數據利用IBM SPSS Statistics 22.0軟件進行單因素方差分析(One-Way ANOVA)新復極差法(Duncan)多重比較和各指標的相關性分析(Pearson),用Microsoft Excel 2019進行數據初步整理和作圖。
2 結果與分析
2.1 不同品種苜蓿第一年越冬率的比較
如圖3所示,參試不同苜蓿品種越冬率差異顯著(P<0.05),其中A-07品種越冬率最高,越冬率為94.01%,A-04越冬率最低,為86.05%,該地區參試苜蓿當年越冬率均在85%以上。
2.2 越冬期不同品種苜蓿根部和根頸抗寒生理響應
2.2.1越冬期不同品種苜蓿根和根頸中3種抗氧化酶活性的比較 由表2可知,參試不同苜蓿品種根部和根頸的SOD活性在不同取樣時期差異顯著(P<0.05),T1時期,A-07根部SOD活性顯著高于其他品種(P<0.05),其活性為400.81 U·min-1·g-1FW;同樣在T1時期,根頸中A-09活性最高,SOD活性為484.68 U·min-1·g-1FW。SOD活性在根部呈先升高后降低再升高的趨勢,而在根頸中呈先降低后升高再降低的趨勢,該酶活性在根部與根頸中的變化特征剛好相反。

表2 參試苜蓿品種越冬不同時期根和根頸超氧化物歧化酶活性
由表3可知,參試不同苜蓿品種根部和根頸的POD活性在不同取樣時期差異顯著(P<0.05),T1時期,A-10根部POD活性顯著高于其他品種(P<0.05),其活性為53 176.59 U·min-1·g-1FW;同樣在T1時期,A-08根頸POD活性顯著高于其他品種(P<0.05),其活性為93 230.83 U·min-1·g-1FW。POD活性在根部和根頸中呈先略有升高然后持續降低的趨勢。
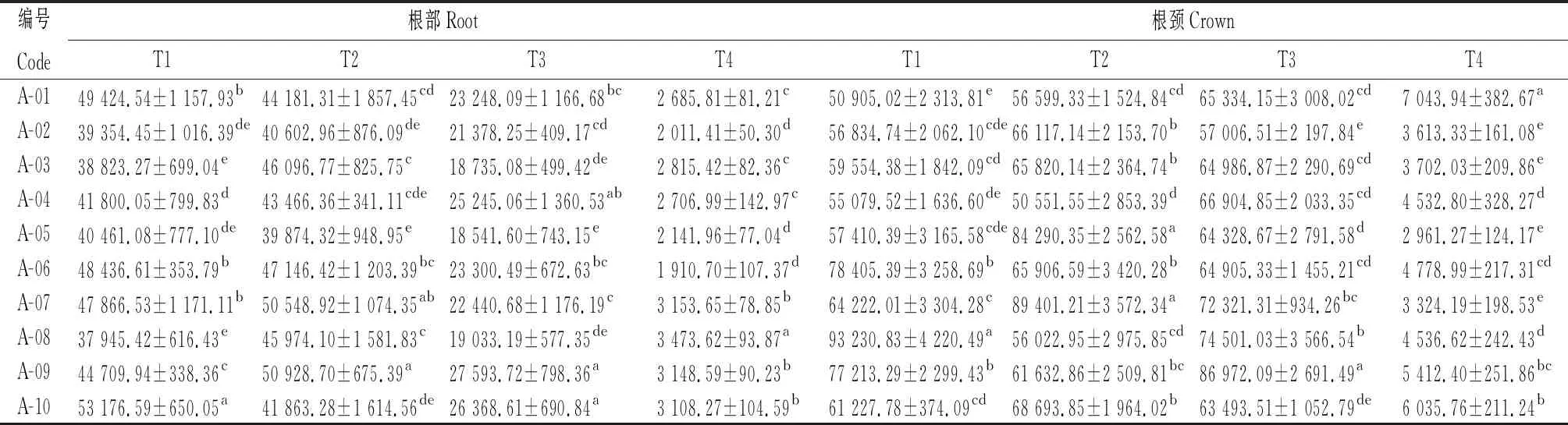
表3 參試苜蓿品種越冬不同時期根和根頸過氧化物酶活性
由表4可知,參試不同苜蓿品種根部和根頸的CAT活性在不同取樣時期差異顯著(P<0.05),T2時期,A-01根部CAT活性最高,其活性為531.43 U·min-1·g-1FW;同樣在T2時期,A-04根頸CAT活性顯著高于其他品種(P<0.05),其活性為1 125.74 U·min-1·g-1FW。根頸和根部的CAT活性變化特征相似,呈先升高后持續降低的趨勢。

表4 參試苜蓿品種越冬不同時期根和根頸過氧化氫酶活性
2.2.2越冬期不同品種苜蓿根和根頸中主要滲透調節物質含量的變化 由表5可知,參試不同苜蓿品種根部和根頸的SS含量在不同取樣時期差異顯著(P<0.05),在T3時期,A-09根部SS含量最高,其含量為131.19 mg·g-1FW;在T2時期,A-10根頸SS含量顯著高于其他品種(P<0.05),其含量為111.78 mg·g-1FW。所有參試品種根部SS含量先持續升高,T3時達到最高,然后下降;在根頸中A-02,A-03,A-04品種SS含量T3時達到最高,而其余參試品種根頸中SS含量在T2時已經達到最高。
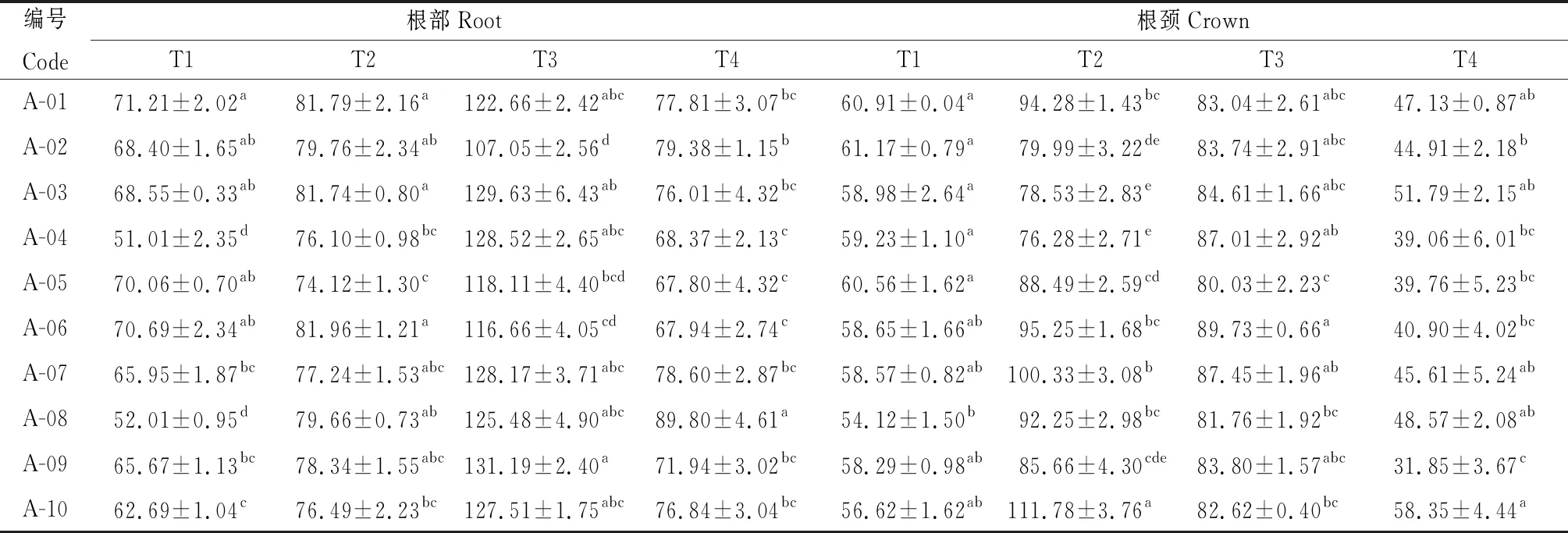
表5 參試苜蓿品種越冬不同時期根和根頸可溶性糖含量
由表6可知,參試不同苜蓿品種根部和根頸的Pro含量在不同取樣時期差異顯著(P<0.05),在T4時期,A-07根部的Pro含量顯著高于其他品種(P<0.05),其值為873.64 μg·g-1FW;在T3時期,A-01根頸中的Pro含量最高,其值為1 044.43 μg·g-1FW。根部和根頸中的Pro含量變化特征較為一致,從T1開始持續上升,到T3達到最高,然后開始下降,在整個越冬期4個取樣時間點,不同苜蓿品種根頸中Pro含量高于對應的根部。

表6 參試苜蓿品種越冬不同時期根和根頸脯氨酸含量
2.2.3越冬期不同品種苜蓿根和根頸丙二醛含量的變化 由表7可知,參試不同苜蓿品種根部和根頸的MDA含量在不同取樣時期差異顯著(P<0.05)。T2時期,A-08根部的MDA含量顯著低于其他品種(P<0.05),其值為10.49 μmol·g-1FW;同樣在T2時期,A-09根頸中的MDA含量最低,其值為17.01 μmol·g-1FW。越冬期間參試品種苜蓿的根部和根頸丙二醛含量在T1時最高,T1到T2間,根部和根頸丙二醛含量降低幅度較大。
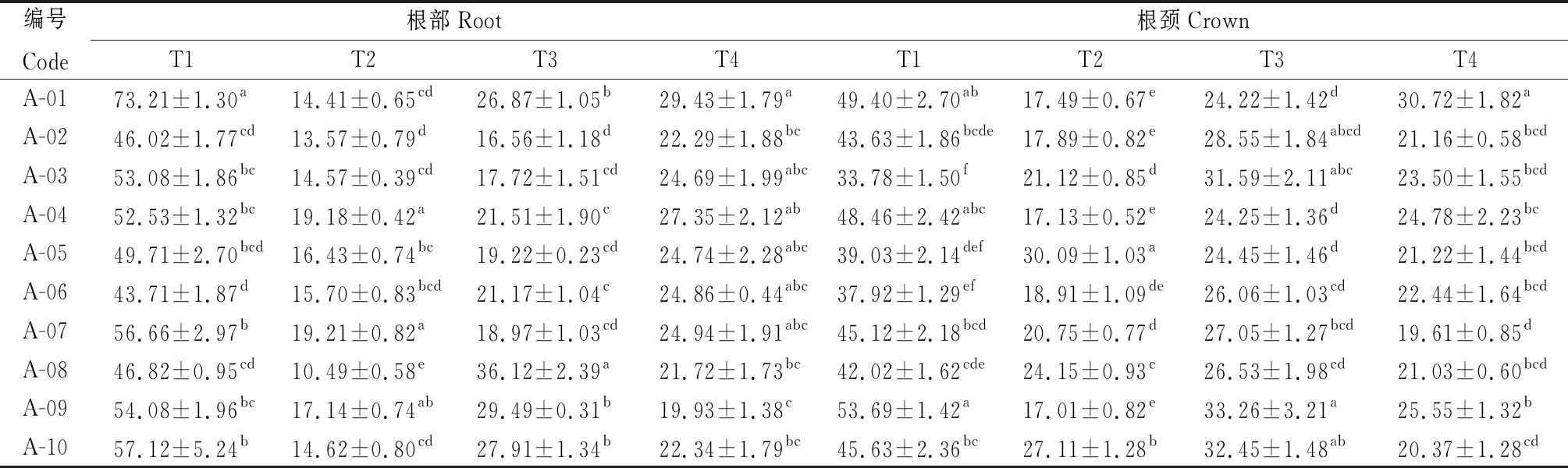
表7 參試苜蓿越冬不同時期根和根頸丙二醛含量
2.3 不同苜蓿品種根系特征分析
2.3.1不同苜蓿品種根系形態指標比較 由圖4可知,不同品種苜蓿根序、總根長、主根平均直徑、根體積4個指標差異顯著(P<0.05)。品種A-09根序值與總根長等2個指標最大,其根序值為6.36(圖4A),總根長為383.45 cm(圖4B);品種A-01主根平均直徑最大,其主根平均直徑為1.47 mm(圖4C);品種A-10根體積最大,其根體積為0.36 cm3(圖4D)。

圖4 不同品種苜蓿根序、總根長、主根平均直徑、根體積比較
由圖5可知,各參試不同苜蓿品種間總根表面積、平均根系直徑、根尖數、分叉數4個指標差異顯著(P<0.05)。品種A-09總根表面積、根尖數與分叉數等3個指標最大,其總根表面積為40.18 cm2(圖5A),根尖數為557.13個(圖5C),分叉數為734.14個(圖5D);品種A-08平均根系直徑最大,其平均根系直徑為0.41 mm(圖5B)。
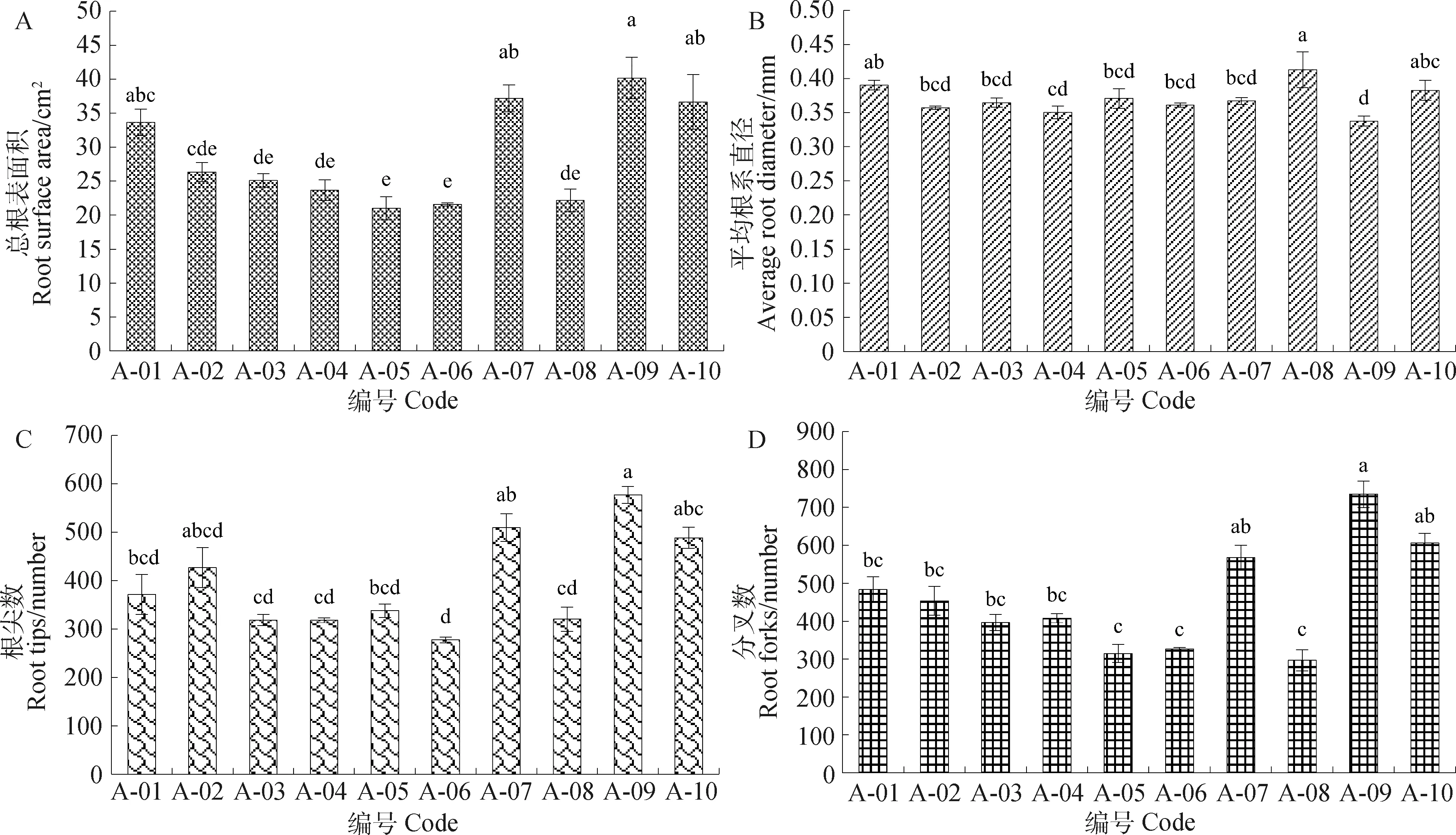
圖5 不同品種苜蓿總根表面積、平均根系直徑、根尖數、分叉數比較
2.3.2不同品種苜蓿根系拓撲指數比較 由圖6可知參試各苜蓿品種拓撲指數差異顯著(P<0.05),品種A-08拓撲指數最大,其拓撲指數為0.82;品種A-09根系拓撲指數最低,其拓撲指數為0.72。

圖6 不同品種苜蓿根系構型比較
2.4 苜蓿生理及根系特征指標與越冬率的相關性分析
如表8所示,參試苜蓿的總根長、總根表面積、主根平均直徑、T4時期根部脯氨酸含量及T1時期根頸可溶性糖含量與其越冬率呈極顯著正相關關系(P<0.01);總根體積、根尖數、分叉數及T4時期根頸脯氨酸含量與越冬率呈顯著正相關關系(P<0.05);而T2時期根頸過氧化氫酶活性與越冬率呈顯著負相關關系(P<0.05)。

表8 與越冬率有關指標的相關關系
選取10個與越冬率有顯著相關性的指標,利用主成分分析,如表9所示,前三個成分的特征值大于1,累計貢獻率為78.59%,提取的3個公因子可以代表絕大部分信息。
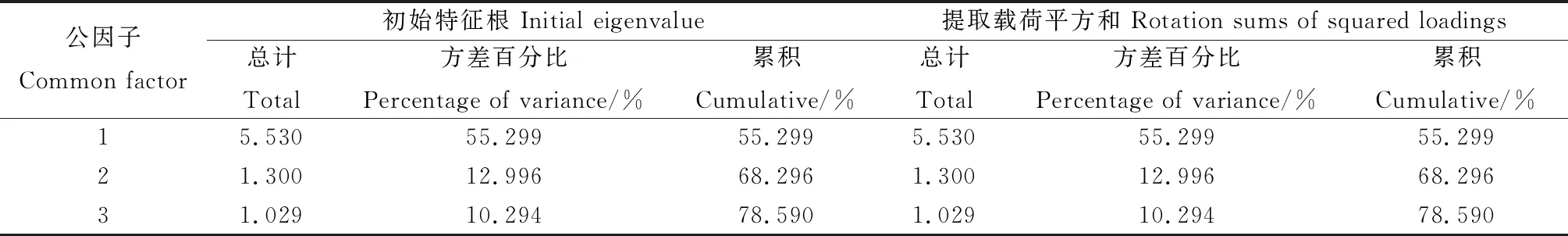
表9 與越冬率相關指標公因子和方差貢獻率
進一步的利用與越冬率有關的因子主成分載荷矩陣,如表10所示,可得到3個公因子F1,F2,F3(其中,X1表示總根長;X2表示總根表面積;X3表示主根平均直徑;X4表示總根體積;X5表示根尖數;X6表示分叉數;X7表示T4根部Pro;X8表示T2根頸CAT;X9表示T4根頸Pro;X10表示T1根頸SS,下同)。F1=0.964X1+0.978X2+0.752X3+0.941X4+0.921X5+0.953X6+0.502X7+0.003X8+0.379X9+0.204X10;F2=—0.106X1—0.141X2—0.089X3—0.175X4—0.074X5—0.110X6+0.531X7—0.096X8+0.588X9+0.759X10;F3=0.008X1—0.014X2—0.100X3—0.044X4+0.036X5+0.096X6+0.038X7+0.957X8—0.187X9+0.234X10。

表10 與越冬率有關的因子主成分載荷矩陣
利用公因子的方差百分比和上述分析中各參數所對應的系數,計算綜合因子,最終得到綜合模型:
F=0.520 13X1+0.521 059X2+0.393 988X3+0.493 091X4+0.503 393X5+0.522 586X6+0.350 521X7+0.087 696X8+0.266 75X9+0.235 538X10
X6,X2,X1的相關系數分別為0.522 6,0.521 1,0.520 1,對分叉數、總根表面積、總根長進行回歸后得到回歸方程:
Y=0.842—1.135X6+0.501X2+1.075X1(R2=0.325,P=0.015)
由上述分析可知,苜蓿根系的分叉數、總根表面積、總根長對苜蓿越冬率影響較大。
3 討論
3.1 越冬期苜蓿根和根頸中生理指標對低溫的響應
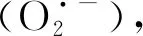

在植物抗寒過程中SS和Pro均具有滲透調節的功能,SS和Pro等滲透調節物質在低溫馴化階段的積累改變了細胞的滲透勢,從而減少了細胞間形成的冰與細胞內溶液之間的水勢差,使得水從細胞中滲出的速率降低。相關研究表明,低溫誘導的糖代謝調節與耐冷性之間存在因果關系[29]。本試驗中,苜蓿根部和根頸中可溶性糖含量隨著溫度的降低而升高,到了T4(返青前)可溶性糖含量明顯降低,與魏雙霞等人[30]的研究結果一致,在擬南芥中,糖積累的另一個作用是避免冰成核,抗寒鍛煉期間SS積累與較低的冰成核溫度(冰晶開始形成的溫度)相關[31],到苜蓿返青前,SS作為貯藏性物質被消耗利用。
Pro被認為是相容性溶質,據報道它在保護酶免受變性、穩定蛋白質合成機制、增加水結合能力以及充當碳氮源的儲藏中起著重要作用[32]。本試驗中,苜蓿根頸與根部Pro隨著溫度的降低持續升高,起到滲透調節等保護作用,到T4(返青前)根部和根頸中的Pro含量均維持在較高的水平,為返青提供碳和氮的來源來促進生長[33];根頸中的Pro含量均高于根部,Pro含量的變化與環境脅迫緊密相關;T4(返青前)根頸中的Pro含量與越冬率顯著正相關(P<0.05),但根頸中T1(抗寒鍛煉初期)的SS含量與越冬率呈極顯著正相關(P<0.01),關于這一結果,文獻中未見報道,需進一步深入研究。
MDA是脂質過氧化的最終產物,其含量反映了膜脂過氧化程度[34]。本試驗中根部與根頸的MDA含量在抗寒鍛煉初期就達到較高的水平,到抗寒鍛煉末期MDA含量達到最低水平,與此同時根部和根頸中的CAT活性達到最高,抗寒鍛煉階段MDA的累積量可能與CAT的活性有關。
3.2 苜蓿根系特征與抗寒性的關系
苜蓿為直根系多年生牧草,根系是苜蓿養分吸收、運輸、貯存的主要功能器官,其生長發育狀況直接影響其產量以及飼用品質,并且在自然越冬過程中根系與苜蓿的抗寒性能密切相關[35]。本試驗中,根序級與苜蓿越冬率相關性不顯著,根序級在一定程度上反映了不同苜蓿品種細根系統內部結構上的異質性;越冬率與總根長與總根表面積極顯著正相關,總根長和總根表面積反映了根系空間擴張、吸收養分和吸收土壤水分的能力,所以總根長和總根表面積較大的苜蓿品種獲得水分和營養元素的能力越強,適應環境的能力越強[36]。本試驗中主根平均直徑與越冬率呈極顯著相關關系,周洋洋[37]的研究也表明主根直徑與越冬率呈極顯著正相關關系,說明主根的營養物質積累和發育狀況與苜蓿的抗寒性密切相關;總根體積、根尖數和分叉數也與苜蓿越冬率顯著正相關,而有研究表明,總根體積越大、根尖數越多分叉數越多則說明根部在土壤中的分布越廣、根系代謝越旺盛、各級根向四周的擴展范圍能力越強[38]。根系的拓撲指數反映了不同植物根系的分支模式,其直接影響到植物根系吸收效率和根系構建過程中所需的碳量[39],本研究中越冬率最高的A-07,A-09兩個品種拓撲指數更接近于0.5,也有在藜科植物中的研究表明,叉狀分支模式的根系,在高海拔可以迅速獲得營養物質,這有助于它們在營養有限的生境中具有潛在的競爭力[19]。
4 結論
10個不同品種苜蓿在青藏高原東緣半濕潤地區的越冬過程中越冬率均在85%以上;返青前根頸和根部脯氨酸含量和越冬初根頸可溶性糖含量與越冬率有相關性;根頸生理活動較根部敏感;苜蓿根系特征指標如分叉數、總根表面積、總根長是影響苜蓿越冬率的主要指標;根系發達、擴展性較強的‘WL319HQ’、‘皇后’、‘太陽神’、‘馴鹿’等苜蓿品種在該地區具有較好的越冬能力。
