雞糞模擬堆肥中多重耐藥菌、耐藥基因和整合酶基因的消減動力學解析
2022-04-25 05:26:12姜欣然孫興濱唐偉欣王旭明高浩澤仇天雷
環境科學研究 2022年4期
關鍵詞:耐藥
姜欣然,李 濤,孫興濱,唐偉欣,王旭明,高浩澤,仇天雷*
1. 東北林業大學林學院,黑龍江 哈爾濱 150040
2. 北京市農林科學院生物技術研究所,農業基因資源與生物技術北京市重點實驗室,北京 100097
由于抗生素在促生長、預防和治療動物疾病方面具有良好的效果,所以長期以來被廣泛用于集約化畜禽養殖業. 據統計,我國是主要的抗生素原料藥生產和抗生素產品消費國之一,年產量超過100萬噸,年消費量約為世界總量的12.5%. 獸用抗生素的長期使用直接導致畜禽糞便中抗生素耐藥菌和耐藥基因的大量殘留. 耐藥細菌隨畜禽糞便排出后,其攜帶的耐藥基因可以通過質粒、轉座子、整合子等可移動元件傳遞給其他細菌,從而導致耐藥基因的傳播和擴散. 整合子是耐藥基因傳播的重要元件,可以通過整合酶基因位點不斷從外部環境中整合一種或幾種耐藥基因盒,使細菌獲得耐藥性或多重耐藥性. 吳聰明等研究發現,長期使用氨基糖苷類和磺胺類藥物后,從豬糞中分離的菌株對多種抗生素表現出耐藥性,同時檢測到Ⅰ類整合酶基因中含有及基因盒. 魏秀麗等從山東地區肉雞體內分離的多重耐藥的大腸桿菌中也檢測到+5基因盒. Vo等研究發現,從牛體內分離的沙門氏菌攜帶有和等基因盒,并對磺胺類及氯霉素類等藥物表現出耐藥性,說明整合子結構對養殖糞便中條件致病菌的多重耐藥表型十分重要.
高溫堆肥能夠利用微生物的好氧發酵作用分解并穩定畜禽糞便中的有機物,形成腐植酸豐富的有機肥,是畜禽糞便無害化利用的常用技術. 目前研究表明,在畜禽糞便堆肥過程中,耐藥基因的相對豐度變化與整合酶基因呈顯著相關,堆肥后部分與Ⅰ類整合子-整合酶基因(,簡稱“Ⅰ類整合酶基因”)相關的耐藥基因〔如磺胺類耐藥基因()〕的豐度反而會出現增長. 然而目前多數研究關注的是堆肥過程中耐藥基因和可移動元件的豐度變化及其相關性分析,而對于耐藥基因消減動力學以及整合酶基因的影響研究較少. 因此,該研究通過在模擬堆肥中定量添加含有多重耐藥整合子的大腸桿菌,利用選擇平板計數、數字微滴PCR技術并結合非線性回歸分析,獲取多重耐藥菌數量、Ⅰ類整合酶基因()及相關耐藥基因(、)、非整合子耐藥基因(和)的絕對豐度和相對豐度,及其在堆肥過程中的變化規律,并進一步分析兩類耐藥基因的消減動力學模型和半消減周期,以期為有效去除養殖源耐藥菌以及可移動耐藥基因提供基礎數據.
1 材料與方法
1.1 樣品的來源及采集
所用雞糞取自北京市平谷區某蛋雞養殖場,輔料為玉米秸稈粉. 采集糞便樣品后,迅速帶回實驗室于4 ℃下保藏,在24 h內進行耐藥菌的計數. 菌株為分離自養殖場糞便中的多重耐藥大腸桿菌()菌株5A5.
1.2 堆肥試驗設計
堆肥原料及配比:取800 g干雞糞,按雞糞與玉米秸稈粉(粒徑1~3 cm)質量比(以干質量計)2.5∶1的比例進行混合,并攪拌均勻. 根據前期研究結果,雞糞中多重耐藥菌的濃度約為4.26×10CFU/g. 混勻后試驗組加入10%菌體濃度為4.77×10CFU/g的5A5多重耐藥菌懸液,空白組加入等體積蒸餾水,調整含水率至60%,初始C/N為15,攪拌混勻后分裝三角瓶后置于水浴中,堆肥原料成分見表1.
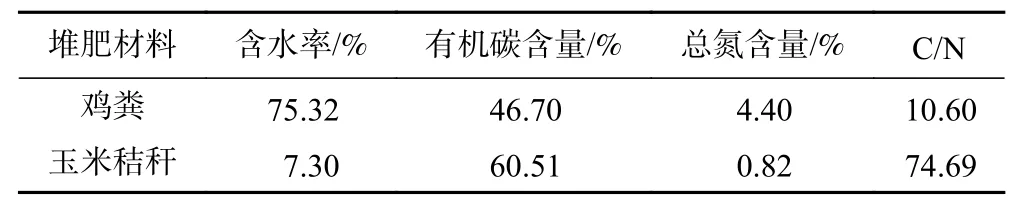
表 1 堆肥原料成分Table 1 Components of raw manure and corn stalks
試驗設計:堆肥試驗于恒溫水浴箱內進行,高溫堆肥模擬設置試驗組及空白組,每組3個重復. 采用水浴控制堆體溫度,試驗溫度設置見圖1. 每天攪拌混勻2次,曝氣泵充氧5 min.
1.3 總菌、大腸桿菌計數及多重耐藥菌計數
稱取10 g模擬堆肥樣品,倒入已滅菌的90 mL生理鹽水三角瓶中,搖床振蕩15 min使堆肥樣品均勻分散,吸取懸浮液進行梯度稀釋. 稀釋液涂布對應培養基:細菌總數采用普通LB營養瓊脂,大腸桿菌計數采用大腸桿菌顯色培養基. 滅菌后的培養基中添加抗生素混合液,涂布后平板倒置于28 ℃恒溫箱中,培養48 h,選取菌落數為30~300個的平板進行計數. 多重耐藥抗生素種類為家禽養殖中常用的抗生素,抗生素名稱及培養基終濃度分別為四環素(16 μg/mL)、恩諾沙星(1 μg/mL)、磺胺甲唑(76 μg/mL)、泰樂菌素(1 μg/mL),抑菌濃度參考2011年美國臨床實驗室標準化協會(CLSI)規定值.
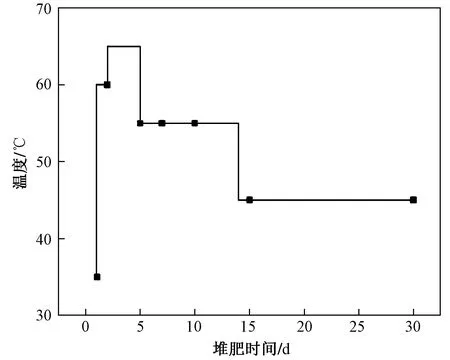
圖 1 堆肥試驗溫度控制示意Fig.1 Temperature diagram of composting experiment
1.4 多重耐藥菌耐藥質粒鑒定及耐藥基因定量檢測
細菌質粒DNA提取及測序
使用QIAGEN Plasmid Mini Kit試劑盒提取5A5菌株的質粒DNA,經檢測后送至諾禾致源公司進行基因測序.
在8 334張門急診處方中,不合理處方50張,不合格率為0.6%。不合理處方點評結果(見表2,表3,表4)。
耐藥基因引物設計與數字PCR定量檢測
根據多重耐藥菌5A5中質粒上攜帶的耐藥基因測序序列,設計氨基糖苷類耐藥基因()、磺胺類耐藥基因()、粘桿菌素耐藥基因()及喹諾酮耐藥基因()對應的Taqman探針引物;并根據多重耐藥菌株腸桿菌16S rRNA基因,設計特異性定量引物(見表2),Ⅰ類整合酶基因()和細菌16S rRNA基因引物參照文獻[11]. 數字微滴式PCR反 應在QX200 Droplet Digital PCR (ddPCR)Systems中完成,選用Trans PCR SuperMix. 反應條件:95 ℃預變性5 min;95 ℃變性30 s,58 ℃退火30 s,72 ℃延伸30 s,共40個循環;72 ℃延伸10 min.

表 2 菌株5A5引物名稱及序列Table 2 Primer names and sequences of strain 5A5
1.5 數據統計分析
采用Excel 2016軟件對試驗數據進行處理后,根據時間與耐藥基因豐度的變化,運用Origin 2019b軟件中的非線性回歸分析對試驗數據進行擬合,使用的一級動力學反應方程如下:

式中:為時耐藥基因的絕對豐度,copies/g;為堆肥開始時的耐藥基因的絕對豐度,copies/g;為耐藥基因消減速率常數,d.
2 結果與分析
2.1 耐藥菌株的質粒獲取及結構解析
將多重耐藥菌株5A5提取的質粒進行二代測序分析,結果表明,5A5菌株質粒上攜帶有多種類型耐藥基因及整合子(見表3),包括及等12種耐藥基因,以及Ⅰ類整合酶基因(). 這些耐藥基因主要對頭孢菌素、氨基糖苷類、磺胺類、二氨基嘧啶、多肽類、氟喹諾酮類、大環內酯類及氯霉素類8種常用抗生素具有耐藥性. 而根據5A5菌株的抗生素藥敏試驗結果,該菌在頭孢噻呋、頭孢哌酮、氟苯尼考、氯霉素、環丙沙星、恩諾沙星、青霉素、慶大霉素、諾氟沙星、氨芐西林、紅霉素、磺胺甲唑、左氧氟沙星等抗生素上均具有耐藥性,結合基因型和表型分析結果,說明該菌耐藥表型的獲得與耐藥質粒上攜帶的耐藥基因直接相關.

表 3 菌株5A5中耐藥基因及整合酶基因信息Table 3 ARGs and integrase gene on strain 5A5
氨基糖苷類、磺胺類、喹諾酮類抗生素是畜禽養殖過程中常規使用的抗生素,而大腸桿菌亦為家禽內優勢的腸道微生物,由于該菌適合腸道環境并且在有抗生素選擇壓力的情況下,能夠在家禽腸道中長期存在,其質粒上攜帶的多種抗生素耐藥性可在不同種類細菌中轉移和傳播,具有較大的環境風險;大腸桿菌在人、動物體內和自然環境中廣泛存在,是常見的條件性病原菌之一,具有接受、儲備和傳遞耐藥遺傳因子的能力,常作為耐藥水平的指示菌. 因此選擇該菌作為模式菌株添加進入堆肥系統中,研究其質粒上多重耐藥基因隨堆肥過程發生的變化,以期獲得高溫堆肥對多重耐藥菌及其耐藥基因的消減規律,降低環境中由于畜禽養殖帶來的多重耐藥污染的傳播風險.
2.2 高溫堆肥過程中可培養耐藥大腸桿菌及總菌數的變化
好氧堆肥的高溫條件能夠影響中溫菌的生長,對大腸桿菌等條件致病菌具有有效的滅殺作用. 將不同堆肥時段(1、2、5、7、10 d)采集的樣品分別進行大腸桿菌和總菌群計數,并同時在多重耐藥平板上進行多重耐藥大腸桿菌和總耐藥菌計數(見表4).
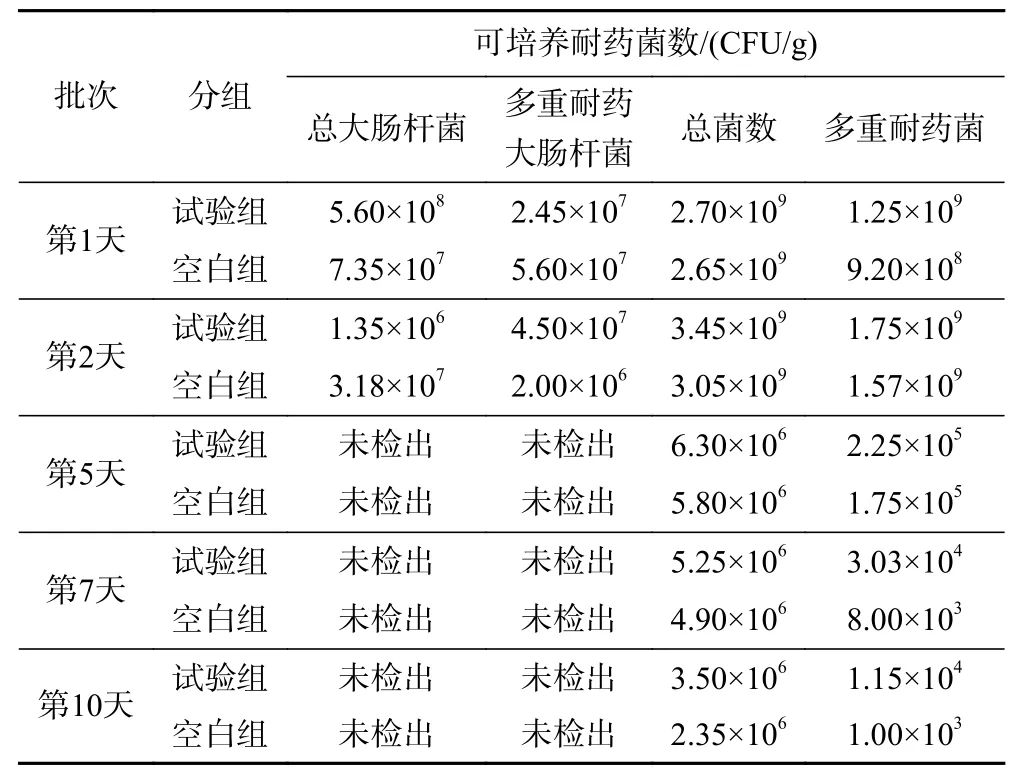
表 4 堆肥過程中可培養耐藥菌的計數結果Table 4 Counting results of cultivatable drug-resistant bacteria during composting
結果表明,無論常規還是多重耐藥的大腸桿菌,高溫堆肥對其都有良好的滅殺作用,堆肥5 d檢測時,已無可培養的大腸桿菌被檢出. 雖然接種了多重耐藥大腸桿菌的試驗組初期(第1、2天)大腸桿菌和多重耐藥大腸桿菌均高于對照組,但第5天的樣品中已檢測不到大腸桿菌,該結果與前期單一耐藥大腸桿菌結果相似,說明抗生素耐藥的獲得并不能改變大腸桿菌的耐熱性能. 高溫堆肥對中溫細菌的抑制作用明顯,與中溫細菌總菌數相比,可培養的多重耐藥菌對高溫堆肥更為敏感,從堆肥初始(第1、2天)到第10天,可培養的多重耐藥菌總數降低了4~6個數量級,而總菌數僅降低了3個數量級.
2.3 高溫堆肥過程中耐藥基因豐度的變化
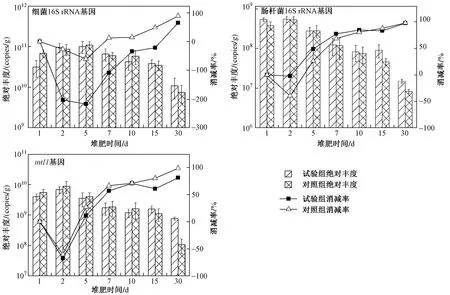
圖 2 高溫堆肥過程中16S rRNA和整合酶基因絕對豐度的變化Fig.2 Absolute abundance of 16S rRNA and integrase genes during composting
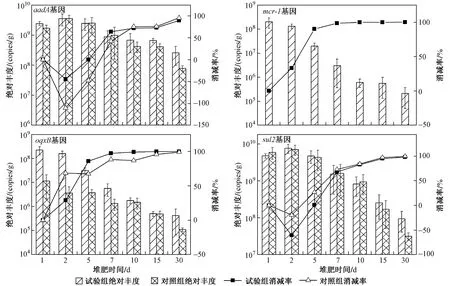
圖 3 高溫堆肥過程中耐藥基因絕對豐度的變化Fig.3 Absolute abundance of drug-resistant genes during composting
基因的絕對豐度表征每克堆肥樣品中該基因的絕對拷貝量,16S rRNA基因的絕對豐度代表環境中細菌總量,基因的絕對豐度代表環境中Ⅰ類整合酶基因的數量. 結果(見圖2)表明,在連續的30 d堆肥過程中,16S rRNA基因和Ⅰ類整合酶基因的絕對豐度逐漸降低,這與可培養的耐藥菌數量變化趨勢相一致. 其中,代表總菌的細菌16S rRNA基因絕對豐度最高,在10~10copies/g之間;代表大腸桿菌的腸桿菌16S rRNA基因絕對豐度最低,在10~10copies/g之間;而基因的絕對豐度在10~10copies/g之間. 到堆肥第30天,腸桿菌16S rRNA基因的絕對豐度從最初的5.37×10copies/g逐漸降至1.50×10copies/g,其消減率為97.15%;基因的絕對豐度從最初的3.95×10copies/g逐漸降至7.55×10copies/g,消減率為80.89%.
在、、及這4種耐藥基因中,和兩個耐藥基因和Ⅰ類整合酶基因的絕對豐度變化趨勢相同(見圖3). 由于耐藥基因在雞糞原料中并未被檢出,隨著高溫堆肥過程的進行,外源宿主菌5A5被高溫殺死,所以其上攜帶的耐藥基因呈明顯降低趨勢. 由于和基因在雞糞原料中有大量檢出,所以基因絕對豐度較高,堆肥過程其數值變化范圍在10~10copies/g之間. 到堆肥第30天,基因的絕對豐度從初始的2.42×10copies/g降 至2.57×10copies/g,消 減 率 為89.39%;基因的絕對豐度也從初始的4.71×10copies/g降至9.46×10copies/g,消減率為97.99%;基因的絕對豐度則從1.97×10copies/g降至2.09×10copies/g,消減率為99.89%;基因的絕對豐度從2.22×10copies/g降 至4.22×10copies/g,消 減 率 為99.81%. 由此可以看出,連續高溫的堆肥過程對Ⅰ類整合酶基因或是耐藥基因都有著較好的消減效果,而位于整合子基因盒上的耐藥基因(和)的消減效率低于不與整合子相連的耐藥基因(和).
相對豐度是將絕對豐度進行歸一化處理,即每16S rRNA拷貝數對應的特定基因拷貝數,表征該環境下微生物群落中該基因的相對豐度. 耐藥基因的相對豐度通常表征菌群中相應耐藥水平的高低. 堆肥過程中,耐藥基因及Ⅰ類整合酶基因的相對豐度和削減率的關系如圖4所示,可以看出,腸桿菌16S rRNA、、及基因的相對豐度在堆肥過程中均先降低,然后在15~30 d內有小幅上升,而、這兩個基因的相對豐度均隨堆肥時間的增加而持續降低. 經過高溫堆肥后,整合子中基因的相對豐度從0.26降至0.03;耐藥基因的相對豐度從0.17降至0.02;的相對豐度從0.30降至0.008;而耐藥基因、的相對豐度均下降了3個數量級. 腸桿菌16S rRNA、、、基因的削減率分別為95.61%、87.08%、86.37%、97.25%,而、基因的削減率則分別達到99.89 %和99.79 %. Liao等研究表明,雖然高溫堆肥能夠消除一部分初始微生物,但是很多ARGs保留在幸存的微生物群落中,推測堆肥過程中發生了水平基因轉移,因而傳統堆肥方式并不能有效去除ARGs和MGEs.Lin等研究表明,堆肥并不能有效控制磺胺類抗性基因向耐熱菌群(如)中轉移. 這些研究都說明和基因可能通過基因水平轉移到耐高溫的菌群中而得以存活.
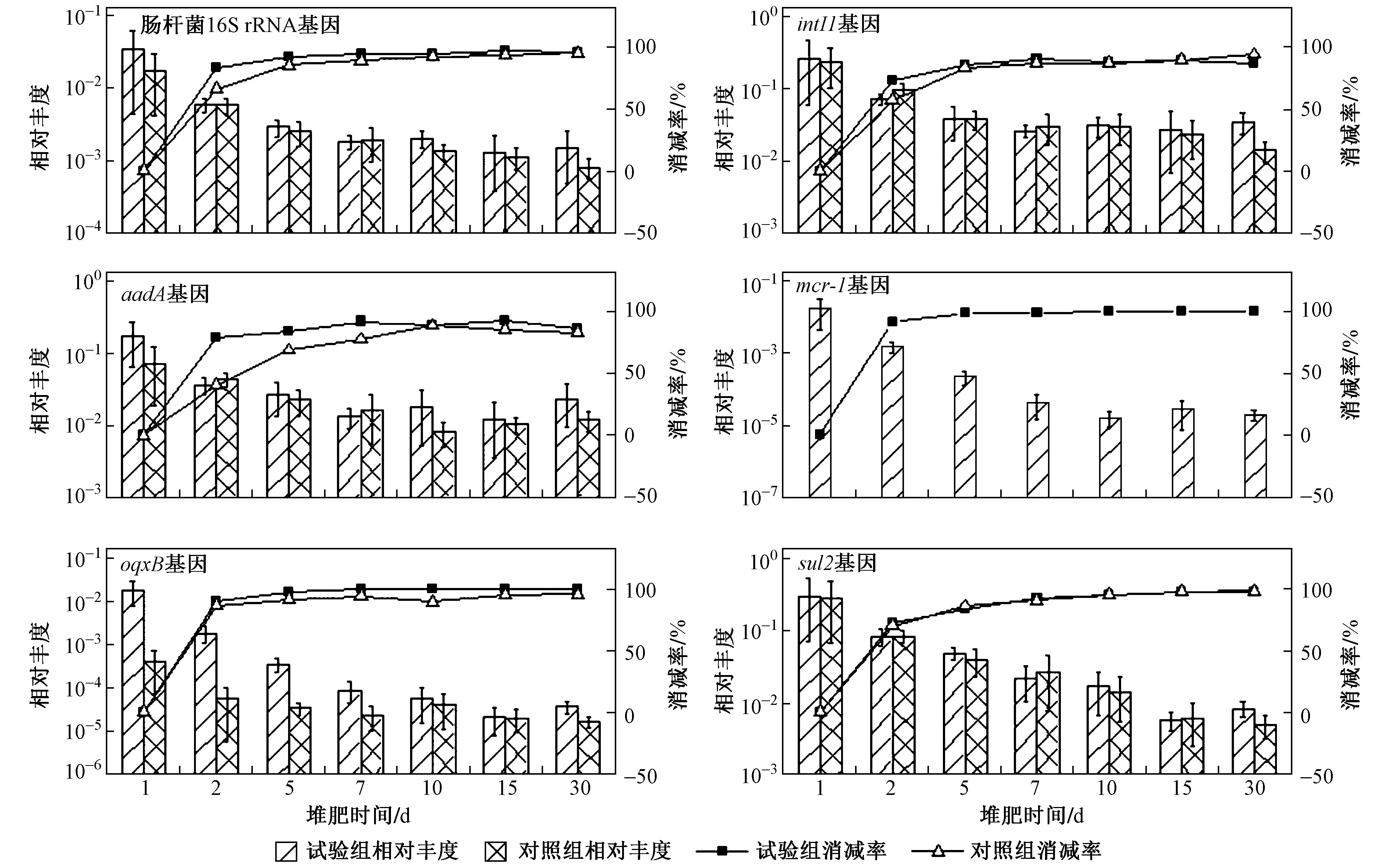
圖 4 高溫堆肥過程中16S rRNA基因、整合酶基因和耐藥基因相對豐度的變化Fig.4 Relative abundance of 16S rRNA, integrase genes and drug-resistant genes during composting
2.4 耐藥基因的消減動力學分析
以絕對豐度和堆肥時間數據為基礎,進行堆肥過程中耐藥基因絕對豐度消減曲線擬合,對比擬合曲線的決定系數()(見表5)發現:①添加多重耐藥菌可以有效模擬外源耐藥基因污染的消減情況,如耐藥基因的試驗組擬合曲線的為0.999,而對照組未檢出基因;耐藥基因的試驗組擬合曲線的為0.998,而對照組的偏低,為0.788. ②養殖環境中廣泛存在的耐藥基因、在試驗組和對照組中的消減規律類似,耐藥基因的試驗組擬合曲線的為0.961,對照組為0.972;耐藥基因的試驗組擬合曲線的為0.924,對照組為0.937;總細菌16S rRNA基因消減曲線的無論是試驗組還是對照組都偏低,分別為0.833和0.695,主要原因是,堆肥高溫過程除中溫菌受抑制外,高溫菌數量會有所提升,導致總細菌的消減規律并不符合一級動力學方程;而腸桿菌科特異16S rRNA基因,無論是試驗組和對照組均符合一級動力學方程,說明堆肥過程能有效處理養殖糞便中的大腸桿菌.
試驗組基因絕對豐度的消減速率從快到慢分別為喹諾酮類耐藥基因、多肽類耐藥基因、磺胺類耐藥基因、腸桿菌16S rRNA基因、氨基糖苷類耐藥基因、整合酶基因. 由于基因是Ⅰ類整合酶基因盒內耐藥基因,所以和基因的消減速率常數和半消減期較為相似,基因的消減速率常數為0.119 d,半消減期為5.81 d,而基因的消減速率常數為0.103 d,半消減期為6.71 d.基因的消減速率常數為0.243 d,半消減期為2.86 d;基因的消減速率常數為0.391 d,半消減期為1.77 d;基因的消減速率常數為0.411 d,半消減期為1.69 d. 腸桿菌16S rRNA基因的消減速率常數為0.128 d,半消減期為5.41 d,說明相對于可培養大腸桿菌在堆肥第5天即完全失活而言,腸桿菌的DNA存活時間要更長(見圖5).
對照組基因絕對豐度曲線擬合結果與試驗組基本一致,但由于糞便中耐藥基因、絕對豐度較低,二者在對照組中的消減規律并不符合一級動力學方程. 從堆肥第2天到第15天,對照組中基因的消減速率常數為0.155 d,半消減期為4.48 d;基因的消減速率常數為0.148 d,半消減期為4.68 d;基因的消減速率常數為0.265 d,半消減期為2.61 d;腸桿菌16S rRNA基因的消減速率常數為0.173 d,半消減期為4 d(見圖6). 總體來說,對照組耐藥基因的半消減期均略低于試驗組,說明耐藥基因的絕對豐度越高,其消減周期越長.
基于耐藥基因相對豐度和堆肥時間數據,進行堆肥過程中耐藥基因相對豐度消減曲線擬合,發現各耐藥基因相對豐度的消減速率普遍低于其絕對豐度的消減速率,如基因相對豐度的消減速率常數為0.192 d,半消減期為3.61 d,基因相對豐度的消減速率常數為0.284 d,半消減期為2.44 d,皆明顯低于相應的絕對豐度消減速率. 試驗組基因的相對豐度消減擬合曲線的僅為0.876,說明基因相對豐度的消減規律并不符合一級動力學模型.而、基因的相對豐度擬合曲線的分別達到0.984、0.999,說明“獨立”耐藥基因的消減規律符合一級動力學模型. 相對豐度數值實際是絕對豐度與16S rRNA基因絕對豐度的比值,如果符合一級動力學模型,即表明堆肥使得含有耐藥基因的比例能以穩定速率減少,即可以通過延長堆肥時間來有效減少該耐藥基因在堆肥中的相對豐度. 相反,相對豐度消減規律不符合一級動力學模型,說明該耐藥基因(如)相對豐度的消減速率并不穩定,需要進一步分析其影響因素.
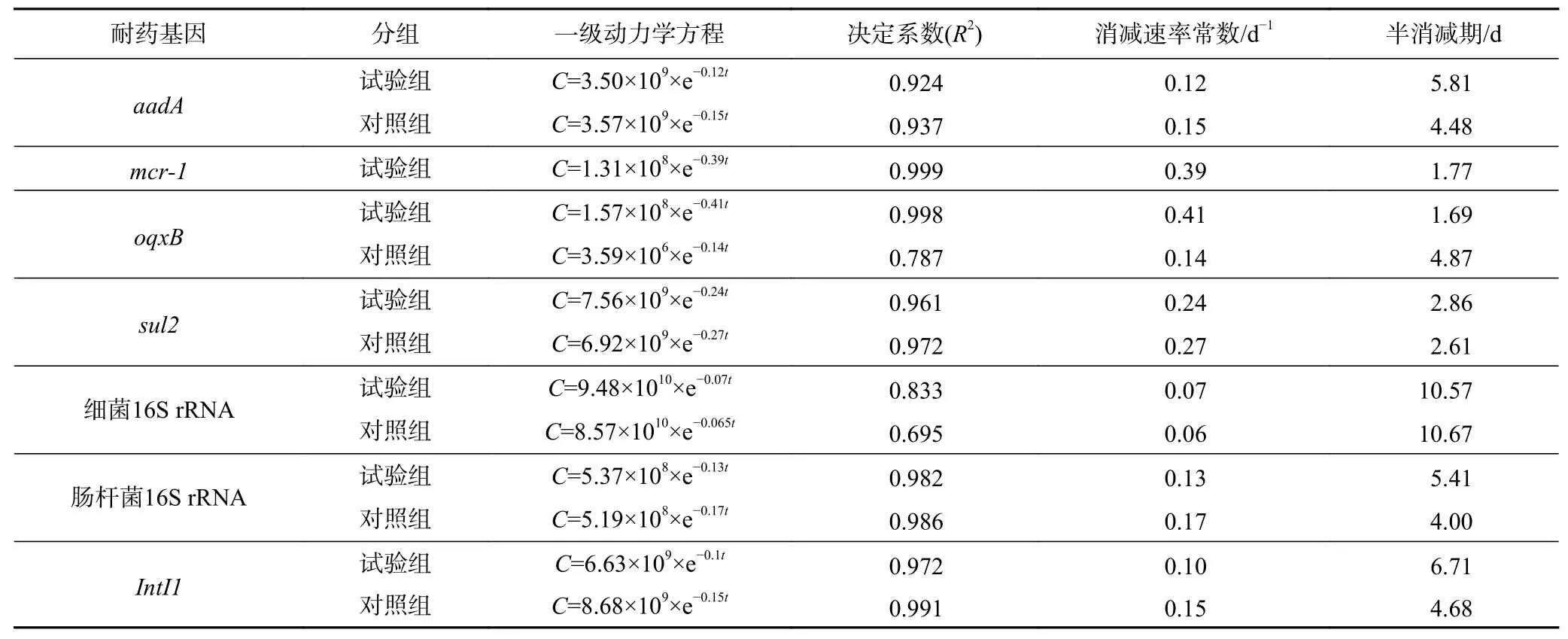
表 5 堆肥中耐藥基因絕對豐度(C)的消減速率和半消減期Table 5 Dissipation rate and elimination half-life of ARGs in compost based on absolute abuandance (C)
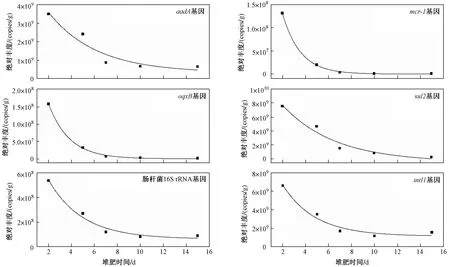
圖 5 試驗組高溫堆肥過程中耐藥基因絕對豐度的消減曲線Fig.5 Dissipation kinetics of ARGs during composting based on aboslute abundance
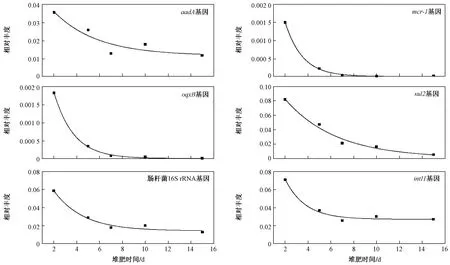
圖 6 試驗組高溫堆肥過程中耐藥基因相對豐度的消減曲線Fig.6 Dissipation kinetics of ARGs during composting based on relative abundance
3 討論
3.1 堆肥對耐藥大腸桿菌的去除效果對比
養殖業中大量使用抗生素使得大量未被利用的抗生素直接接觸環境,導致抗生素賦存濃度升高.農業生產中畜禽糞污還田還會將殘留的抗生素帶入農田土壤. 獸用抗生素不僅導致養殖環境的抗生素污染,還可引發環境內耐藥細菌的出現. 隨著廣譜抗菌藥物的廣泛應用,細菌的耐藥性正在不斷增加并呈現多重耐藥甚至泛耐藥性的趨勢. 堆肥是去除抗生素耐藥菌的有效手段,前期研究表明,經過二次腐熟后堆肥能夠有效殺滅致病菌(沙門氏菌和大腸菌群),并且能夠有效消減四環素耐藥菌的絕對數量.在此次模擬堆肥中,到第5天已經沒有可培養的大腸桿菌被檢出,腸桿菌16S rRNA基因的絕對豐度從最初的5.37×10copies/g逐漸降至1.50×10copies/g(第30天),其消減率為97.15%,同時可培養的多重耐藥菌總數也降低了5~6個數量級. 與這一結果類似的是,李含雄等研究發現,糞便中大腸桿菌數量大于10CFU/g,但通過控制堆肥溫度可以在12 h~5 d內將大腸桿菌全部殺滅.
3.2 堆肥對耐藥基因的去除效果對比
磺胺類藥物是第一批可以全身使用的選擇性作用于細菌的藥物,如今它們很少被使用,部分原因是磺胺類藥物極易產生耐藥性并會在環境中廣泛傳播.Le-Devendec等在對雞糞為期6周的堆肥試驗結果表明,磺胺類抗生素耐藥基因的相對豐度逐漸下降了3~4個數量級. Wu等在對豬糞的普通堆肥中發現,的絕對豐度有明顯下降,堆肥結束后其消減率為85.79%. 這與筆者所得結果一致,此次試驗中基因的絕對豐度從初始的4.71×10copies/g降至9.46×10copies/g(第30天),最終消減率為97.99 %.然而也有部分研究結果顯示,在堆肥過程中耐藥基因絕對豐度會有所升高或在堆肥結束后耐藥基因會有所富集. 這些結果說明,不同堆肥原料會影響磺胺類耐藥基因的去除,溫度可能通過影響磺胺抗性質粒在堆肥中的存活能力和接合轉移能力,進而使得堆肥進程中耐藥基因相對豐度發生改變.
多黏菌素是一種發現于多黏桿菌培養液中的具有抗菌活性的多肽.是在我國首次發現的第一個腸桿菌科中質粒介導的多粘菌素抗性機制的耐藥基因,并且含有基因的質粒具有較高的體外轉移率(10~10),會在人類群體中廣泛傳播,影響人類的正常生活和疾病治療. Xing等采用半透膜堆肥法對銅污染雞糞中和的去除效果進行了評價,結果表明,低銅處理組的相對豐度降低了80.1%. 還有研究顯示,在堆肥過程中,15 d的高溫(44~65 °C)堆肥后,糞便中超過90%的基因被消除,22 d后完全檢測不到基因. 這些研究都與筆者所得結果相近,在此次試驗中耐藥基因的絕對豐度從1.97×10copies/g降至2.09×10copies/g,消減率為99.89%.的相對豐度在堆肥結束后降低了3個數量級. 這些研究結果表明,堆肥能夠有效去除糞便中攜帶的高風險抗生素耐藥基因.
有研究表明,和基因有著緊密關系,Yang等在33個多重抗生素抗性細菌的分離菌株中檢測到了12種移動遺傳元件的存在,并且最終證實所有++均與Ⅰ類整合酶基因相關. 該試驗使用的5A5菌株攜帶質粒就是這種Ⅰ類整合酶基因的結構,因此基因的消減特征與基因類似,在模擬堆肥結束后,通過計算得出試驗組中和基因的消減速率常數分別為0.119和0.103 d,進一步說明連鎖基因結構使其具有相似的消減速率,另外,由于基因位于整合子結構上,其消減速率明顯低于、等獨立基因. 整合子能夠參與基因的水平轉移,使得耐藥基因且在堆肥過程中不容易被去除,這一理論在許多研究中得以證實. Tang等在堆肥過程中發現大多數耐藥基因的拷貝數減少,但基因是唯一絕對豐度增加的基因,同樣在4種可移動遺傳元件中,只有基因的絕對豐度增加,說明可移動遺傳元件可能顯著影響與其相連的耐藥基因的絕對豐度.
研究表明,經過好氧堆肥處理后,抗生素耐藥基因的絕對豐度均得到了有效降低. 孫偉等在污泥堆肥過程中發現,多種耐藥基因在堆肥過程中的消減率均超過85%. Li等研究了在雞糞堆肥中添加竹炭對耐藥基因的影響,發現26 d后,大多數耐藥基因和基因的相對豐度下降了21.6%~99.5%. 在此次模擬堆肥過程中耐藥基因的絕對豐度表現出隨堆肥時間的延長呈持續降低的特征,且在30 d的高溫堆肥過程中,筆者選取的耐藥基因(aadA、sul2、mcr-1、oqxB)和intI1基因的消減率為80.00%~97.99%.大多數耐藥基因的相對豐度表現出先降后略微升高的趨勢,其中,耐藥基因aadA、sul2、mcr-1、oqxB和intI1基因的相對豐度消減率為86.37%~99.89%. 綜上,堆肥是處理畜禽養殖源多重耐藥菌及其耐藥基因污染的有效手段.
4 結論
a) 在模擬堆肥過程中,經過3 d的高溫期后,未檢出可培養的多重耐藥大腸桿菌;經過10 d堆肥后,普通的多重耐藥菌數量降低了4~6個數量級.
b) 多重耐藥菌中耐藥基因的絕對豐度和相對豐度均隨著高溫堆肥過程出現顯著下降,其中4種耐藥基因(aadA、sul2、mcr-1、oqxB)的絕對豐度消減率為89.39%~99.89%,相對豐度消減率為86.37%~99.89%.值得注意的是,整合子上攜帶的aadA基因消減量低于mcr-1等非整合子基因盒內基因,相對豐度也有類似規律.
c) 通過擬合堆肥條件下intI1(整合酶)基因和耐藥基因的消減曲線,進一步獲得了高溫堆肥過程中不同耐藥基因的半消減期. 一方面,堆肥過程中耐藥基因的相對豐度比絕對豐度更難消減;另一方面,與整合子相連的耐藥基因有著更長的半消減期,即更難通過堆肥過程去除.
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
