Pcsk9 基因干擾對高脂誘導的大鼠非酒精性脂肪性肝病合并動脈粥樣硬化的影響
2022-04-22 06:03:36張曉文
上海交通大學學報(醫學版) 2022年2期
張曉文,王 祎,張 嬋,張 迪,贠 航,黃 笛
1.商洛學院健康管理學院護理系,商洛 726000;2.陜西“四主體一聯合”秦嶺健康食品配料及核桃產業技術校企聯合研究中心,商洛 726000
近年來,慢性疾病已成為人類健康的重大威脅。其中,非酒精性脂肪性肝病(non-alcoholic fatty liver disease,NAFLD)和動脈粥樣硬化(atherosclerosis,AS)引起的心血管疾病在慢性疾病中所占比例相當大[1]。NAFLD是最為常見的肝臟疾病,在一般人群中發病率為20%~30%,而在肥胖人群或糖尿病患者中發病率高達70%~90%[2]。某些NAFLD患者可能會進展為肝硬化和肝細胞癌[3]。而且,有研究[4]提出脂肪肝與AS及冠心病關系密切。NAFLD患者具有較高的心血管疾病的患病風險和病死率,且NAFLD患者發生心血管疾病而導致死亡的概率遠遠高于肝臟其他并發癥[5]。因此,對于NAFLD的預防與控制顯得尤為重要。
前蛋白轉化酶枯草溶菌素9 (proprotein convertase subtilisin kexin 9,Pcsk9)基因,編碼一種前蛋白轉化酶,名為神經細胞凋亡調節轉化酶1(neural apoptosis-regulated convertase 1,NARC-1)[6]。該蛋白可介導低密度脂蛋白受體(low-density lipoprotein receptor,LDLR)降解,升高血漿中低密度脂蛋白膽固醇(low density lipoprotein cholesterol,LDL-C)的水平,Pcsk9基因突變會導致異常膽固醇血癥[7]。先前報道[8]顯示,使用Pcsk9抑制劑除了對心血管疾病有良好作用外,還可改善NAFLD,并降低LDL-C 水平。然而,關于Pcsk9如何影響NAFLD 及其調控機制的報道較少。本研究采用基因干擾技術,在高脂誘導的NAFLD 合并AS 大鼠中,考察Pcsk9對大鼠肝臟和主動脈組織損傷,以及對胰島素、血脂及炎癥因子水平的影響。
1 材料和方法
1.1 實驗材料
1.1.1 實驗動物及細胞 40只SPF級,8周齡的雄性SD 大鼠,購自西安交通大學實驗動物中心,體質量為160~200 g,生產許可證號為SCXK(陜)2020-001,并飼養于該實驗動物中心。使用許可證號為SYXK(陜)2020-005。實驗動物倫理審批編號為No.20200521。
人腎上皮細胞系293T 細胞(武漢普諾賽生物科技有限公司),用于包裝重組慢病毒干擾質粒。
1.1.2 主要試劑和儀器Pcsk9-短發夾RNA(short hairpin RNA, shRNA) 和 shRNA- 陰 性 對 照(negative control,NC)重組慢病毒質粒,慢病毒包裝質粒pGag/Pol、pRev、pVSV-G(上海吉瑪基因公司);RNA purification system(德國Qiagen 公司);Superscript RNase H-逆反錄酶(美國Invitrogen 公司);隨機六聚體(美國Sigma-Aldrich 公司);ABI Prism 7000 系統(美國Applied Biosystems 公司);TUNEL細胞凋亡檢測試劑盒(顯色法)(上海碧云天生物技術有限公司);白細胞介素-1β(interleukin-1β,IL-1β)、IL-6 及誘導型一氧化氮合酶(inducible nitric oxide synthase,iNOS);ELISA 試劑盒(美國R&D Systems 公司);10% SDS 聚丙烯酰胺凝膠電泳(SDS polyacrylamide gel electrophoresis,SDS-PAGE)、聚偏二氟乙烯(polyvinylidenefluoride, PVDF)膜、羊抗兔IgG-辣根過氧化物酶(horseradish peroxidase,HRP)、ECL 化學發光檢測試劑盒(北京索萊寶科技有限公司);PCSK9 抗體(#13366)、Toll 樣受體4(Toll-like receptor 4,TLR4)抗體(#14358)、P65 抗體(#8242)、p-P65 抗體(#3033)、腫瘤壞死因子α(tumor necrosis factor α,TNF-α)抗體(#11948)及GAPDH 抗體(#97166)(稀釋度均為1∶1 000,美國CST 公司);p-TLR4 抗體(#PA5-105713,稀釋度1∶500~1∶2 000,美國Thermo Fisher公司)。
全自動生化儀(美國Beehman公司);γ放射免疫計數器(型號GC-2012,科大創新股份有限公司中佳分公司);ABI Prism 7000系統(美國Applied Biosystems公司);E100光學顯微鏡(日本Nikon公司);Multiskan Sky多功能酶標儀(美國Thermo Fisher公司)。
1.2 實驗方法
1.2.1 重組慢病毒干擾質粒的包裝和滴度測定 將構建好的重組慢病毒質粒與慢病毒包裝質粒pGag/Pol、pRev、pVSV-G 按照2∶1∶1∶1 的比例加入混勻,共同轉染293T 細胞。轉染24 h 后,通過熒光顯微鏡觀察熒光強度,與標準滴度的慢病毒載體對照組比較確定大鼠Pcsk9基因的shRNA慢病毒載體病毒滴度。1.2.2 造模與分組 40 只大鼠隨機分為對照組、模型(high fat)組、shRNA-NC 干擾模型(high fat+shRNA-NC)組及Pcsk9-shRNA 干擾模型(high fat+Pcsk9-shRNA)組,每組10只。對照組繼續給予普通飼料,其余組給予高脂飼料(0.3%膽固醇以及20%脂肪)構建NAFLD 大鼠模型;自由攝食,連續喂養14 周。實驗期間,shRNA-NC 干擾模型組和Pcsk9-shRNA 干擾模型組分別通過腹腔注射慢病毒載體shRNA-NC 和Pcsk9-shRNA,每10 d 注射劑量為每只大鼠1×108TU/mL。
1.2.3 實時熒光定量聚合酶鏈反應 采用RNA purification system 提取總RNA,隨后用Superscript RNase H-反轉錄酶和隨機六聚體將1 μg 總RNA 反轉錄成cDNA。使用SYBR Green Master Mix (美國Thermo Fisher公司)進行實時熒光定量聚合酶鏈反應(real-time quantitative polymerase chain reaction,RTqPCR)。檢測基因的引物如下:Pcsk9基因,正向引物5'-GCTGAGCTGCTCCAGTTTCT-3',反向引物5'-AATGGCGTAGACACCCTCAC-3';Gapdh,正 向 引物5'-TGCACCACCAACTGCTTAGC-3',反向引物5'-GGCATGGACTGTGGTCATGAG-3'。所有結果均以Gapdh為基準進行歸一化,用2-ΔΔCt計算。
1.2.4 實驗指標檢測 計算Lee's 肥胖指數:Lee's 指數=體質量(g)1/3×10/體長(cm)。采用放射免疫法測定空腹血清胰島素。全自動生化分析儀檢測大鼠血脂指標: 高密度脂蛋白膽固醇(high density lipoprotein cholesterol,HDL-C)、低密度脂蛋白膽固醇(low density lipoprotein cholesterol,LDL-C)、總膽 固 醇 (total cholesterol, TC) 和 三 酰 甘 油(triacylglycerol,TAG)。
1.2.5 蘇木精-伊紅染色 末次喂食后,處死大鼠,解剖,分別將大鼠肝臟和主動脈組織置于4%多聚甲醛緩沖液中固定后置4 ℃冰箱備用。經乙醇脫水、二甲苯透明后,石蠟包埋、切片,厚度5 μm。組織切片經二甲苯脫蠟,依次經過由高至低濃度梯度乙醇進行水化。接著,用蘇木精染色2 min,1%鹽酸乙醇分化10 s,稀氨水返藍20 s,伊紅染色3 min。再依次經過由低至高濃度梯度乙醇脫水,各5 min。二甲苯透明后用中性樹脂封片。光學顯微鏡觀察染色結果并拍照記錄。
1.2.6 TUNEL 細胞凋亡檢測 根據制造商的說明書進行操作。陽性著色細胞呈棕黃色,位于細胞核,為固縮狀,高倍鏡下呈現碎點或碎塊樣。每張切片隨機觀察10個高倍視野,每個視野計數200個細胞,統計陽性細胞數量,計算細胞凋亡率。細胞凋亡率=凋亡細胞個數/所有細胞個數×100%。
1.2.7 酶聯免疫吸附試驗檢測炎癥因子 經大鼠腹主動脈采血,于4 ℃自然凝固30 min,4 ℃、1 500×g離心10 min,分裝血清并保存于-80 ℃。使用酶聯免疫吸附試驗(enzyme linked immunosorbent assay,ELISA)試劑盒分別檢測IL-1β、IL-6 及iNOS 的表達水平。根據說明書進行操作,在Multiskan Sky 多功能酶標儀450 nm處檢測吸光度。
1.2.8 蛋白質印跡法 用10%SDS-PAGE分離肝臟和主動脈組織中的蛋白,并轉移到PVDF 膜上。5%脫脂牛奶封閉后,蛋白質與第一抗體(一抗)在4 ℃下孵育過夜。主要抗體如下:PCSK9、TLR4、p-TLR4、P65、p-P65、TNF-α 及GAPDH 的抗體。PBS洗滌,羊抗兔IgG-HRP 二抗孵育2 h,增強化學發光(enhanced chemiluminescence,ECL)顯色,分析蛋白質表達水平。
1.3 統計學分析
使用統計軟件SPSS18.0 進行分析。符合正態分布的定量資料采用±s 表示,多組間以單因素方差分析(One-Way ANOVA)進行比較,組間兩兩比較采用LSD法。P<0.05表示差異具有統計學意義。
2 結果
2.1 大鼠Pcsk9基因干擾效率、肥胖指數和胰島素水平的檢測
Pcsk9-shRNA干擾模型大鼠,使Pcsk9基因沉默。RT-qPCR 和蛋白質印跡法分別檢測大鼠肝臟組織中Pcsk9的mRNA 和蛋白水平以驗證干擾效率。如圖1所示,與對照組比較,模型組Pcsk9mRNA 和蛋白表達顯著上調(均P=0.000);與模型組比較,high fat+Pcsk9-shRNA 組Pcsk9的mRNA 和蛋白表達顯著下調(均P=0.000),干擾效率約76%。由此可見,成功敲減了大鼠的Pcsk9基因。表1 的檢測結果顯示,與對照組比較,模型組的肥胖指數和胰島素水平顯著升高(均P=0.000);與模型組比較,high fat+Pcsk9-shRNA 組肥胖指數和胰島素水平顯著下降(P=0.007,P=0.000)。
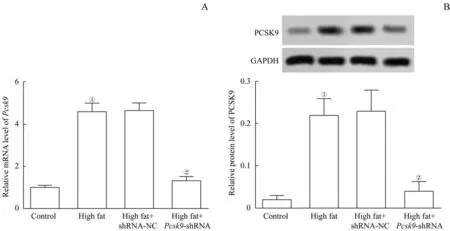
圖1 4組Pcsk9基因干擾的效率檢測Fig 1 Efficiency detection of Pcsk9 gene interference in 4 groups

表1 4組肥胖指數和胰島素水平比較Tab 1 Obesity index and insulin levels in 4 groups
2.2 大鼠肝臟損傷情況
蘇木精-伊紅染色(hematoxylin-eosin staining,HE染色)結果(圖2)所示,對照組大鼠肝組織細胞結構正常,肝細胞以中央靜脈為中心呈放射狀分布;模型組小鼠肝組織中央靜脈呈現不規則形狀,細胞排列不均勻,結構被破壞,出現明顯細胞壞死。與模型組比較,high fat+shRNA-NC 組無明顯改變;high fat+Pcsk9-shRNA組大鼠肝組織細胞的壞死情況得到減輕。由此可見,Pcsk9干擾可以改善NAFLD 大鼠的肝損傷情況。
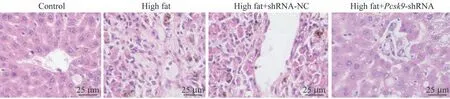
圖2 大鼠肝臟H-E染色(×200)Fig 2 H-E staining of rat livers(×200)
2.3 大鼠肝臟細胞的凋亡
如圖3所示,與對照組比較,模型組的細胞凋亡率顯著升高(P=0.000);與模型組比較,high fat+Pcsk9-shRNA組的細胞凋亡率顯著降低(P=0.000)。由此可見,Pcsk9敲減可抑制NAFLD大鼠肝臟細胞凋亡。
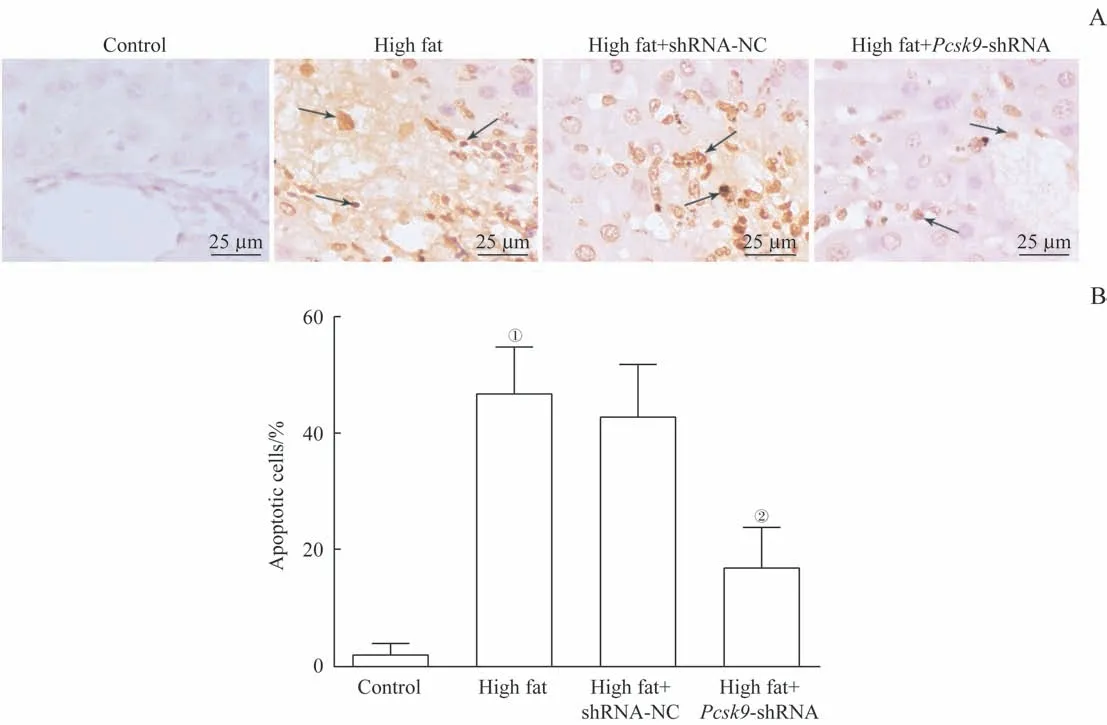
圖3 TUNEL檢測大鼠肝臟細胞凋亡Fig 3 Apoptosis of rat liver cells detected by TUNEL
2.4 大鼠血脂指標的檢測
如表2 所示,與對照組比較,模型組的HDL-C含量顯著降低(P=0.000),而LDL-C、TC 及TAG 含量顯著升高(均P=0.000);與模型組比較,high fat+Pcsk9-shRNA 組 的HDL-C 含 量 顯 著 升 高(P=0.000),而LDL-C、TC 及TAG 含量顯著降低(均P=0.000)。由此可見,Pcsk9敲減可減輕NAFLD 大鼠血脂指標的異常程度。
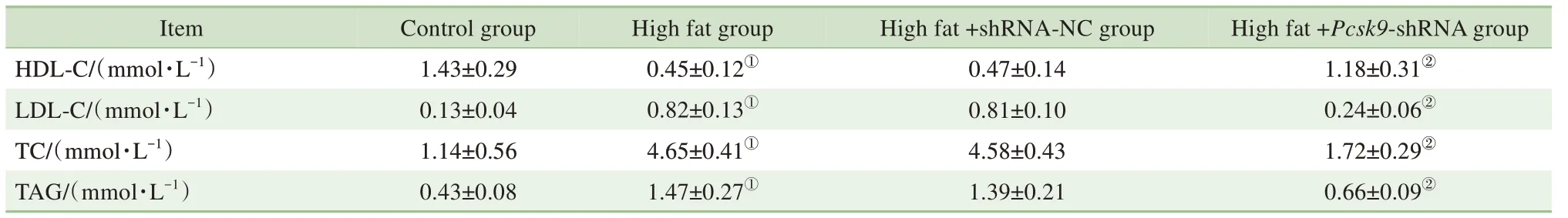
表2 4組大鼠血脂指標比較Tab 2 Blood lipid indicators of rats in 4 groups
2.5 主動脈血管壁形態學變化
顯微鏡下(圖4),對照組主動脈組織結構完整,層次分界清楚,未見斑塊。模型組的動脈內管壁內膜增厚,可見明顯的泡沫細胞和膽固醇結晶,各層細胞脂質浸潤,管腔內壁厚薄不均,有類似粥樣或纖維樣的斑塊。在Pcsk9敲減模型大鼠中,主動脈組織僅有較少斑塊形成,泡沫細胞較少,內膜增生程度較輕。由此表明,抑制Pcsk9表達可改善NAFLD大鼠AS的病理損傷。
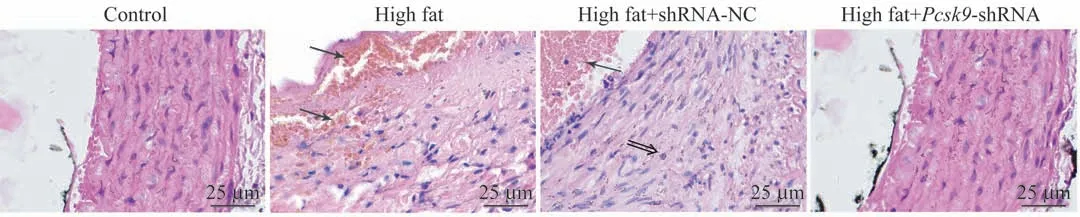
圖4 大鼠主動脈血管壁H-E染色(×200)Fig 4 H-E staining of the aortic wall in rats(×200)
2.6 大鼠外周血中炎癥因子表達水平
如表3 所示,與對照組比較,模型組的IL-1β、IL-6 及iNOS 表達水平顯著升高(均P=0.000);與模型組比較,high fat+Pcsk9-shRNA 組的IL-1β、IL-6及iNOS 表達水平顯著降低(均P=0.000)。由此可見,Pcsk9敲減可抑制NAFLD 大鼠外周血中炎癥因子的表達。
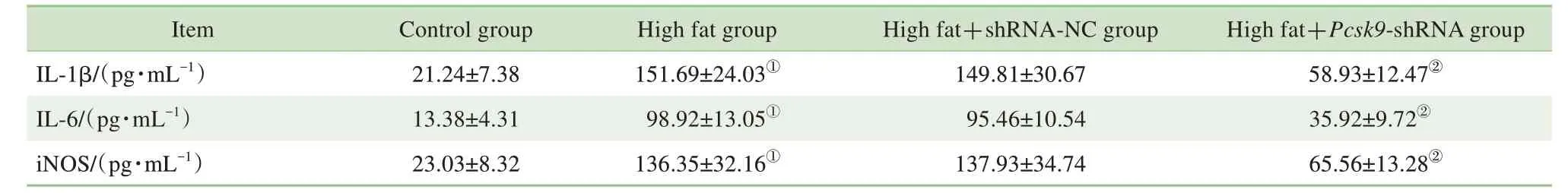
表3 4組大鼠外周血中炎性因子的表達水平比較Tab 3 Expression levels of inflammatory factors in peripheral blood of rats in 4 groups
2.7 大鼠主動脈TLR4、NF-κB P65、TNF-α 蛋白表達
蛋白印跡法檢測了大鼠主動脈組織參與免疫炎癥通路相關的蛋白TLR4 及NF-κB P65 的磷酸化和TNF-α 的表達水平。如圖5 所示,與對照組比較,模型組的TLR4 及NF-κB P65 的磷酸化水平,TNF-α 的表達均顯著上調(均P=0.000);與模型組比較,high fat+Pcsk9-shRNA 組的TLR4 及NF-κB P65 的磷酸化水平,TNF-α 的表達均顯著下調(均P=0.000)。由此可見,Pcsk9干擾可抑制TLR4 及NF-κB P65 的活化,TNF-α 的表達,進而減輕NAFLD 大鼠的炎癥反應。
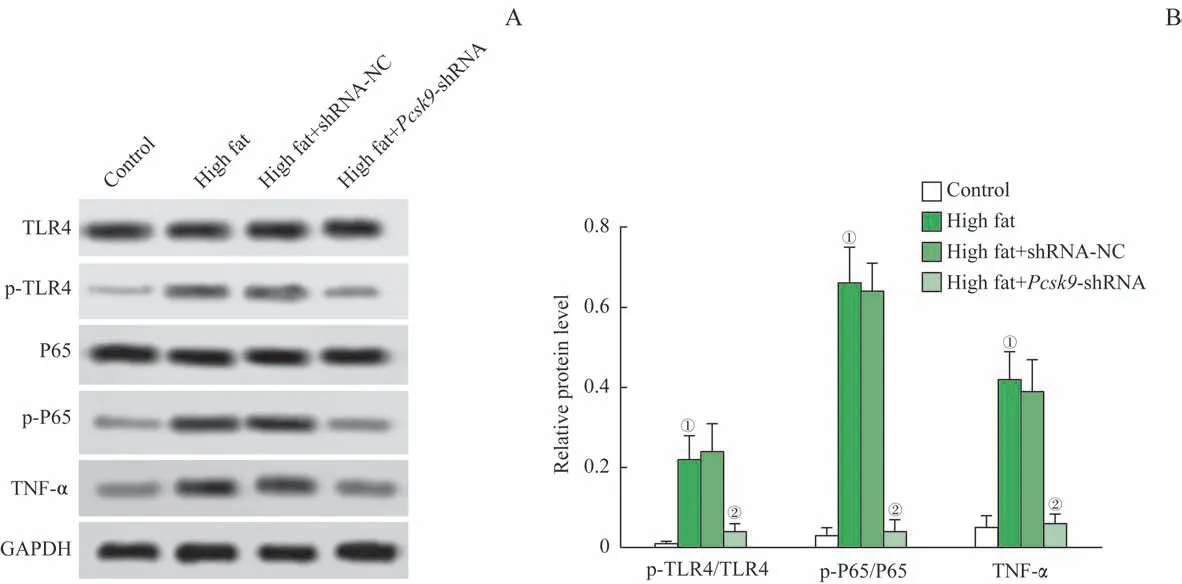
圖5 TLR4及NF-κB P65的磷酸化和TNF-α的表達水平Fig 5 Phosphorylation levels of TLR4 and NF-κB P65 and the expression of TNF-α
3 討論
NAFLD 與諸如胰島素抵抗、肥胖、血脂異常和高血壓等許多代謝性紊亂存在強相關性,因此,也被認為是代謝綜合征在肝臟的集中表現[9]。NAFLD 的發病機制尚不明確,可能與遺傳、代謝和環境等因素有關。目前對于治療NAFLD 尚無特效藥,主要以適量運動、合理飲食和降血糖為主,并適當給予調脂藥物治療[10]。因此,迫切需要探究治療NAFLD的相關分子靶點與機制。
目前研究Pcsk9在人類肝脂肪變性中的作用的研究仍然存在爭議。DEMERS等[11]研究表明在Pcsk9-/-小鼠中,Pcsk9通過下調CD36 的表達來阻止脂質進入肝臟。已知在肝臟中CD36通過促進TAG 的積累和隨后的脂質誘導的內質網應激,促進NAFLD 的進展和更嚴重的非酒精性脂肪性肝炎。在肥胖或者糖尿病的情況下,Pcsk9的缺失突變可以導致肝臟的脂肪積累。然而也有研究[12]發現在正常飲食條件下,Pcsk9-/-小鼠肝臟中的幾種代償機制可能協同作用抑制肝臟脂肪變性;但在高脂代謝挑戰期間,Pcsk9-/-小鼠內質網應激、纖維化、炎癥和凋亡標志物的表達以及血漿谷丙轉氨酶(glutamic-pyruvic transaminase,GPT)水平顯著升高。在本研究中,我們發現干擾Pcsk9表達后NAFLD 大鼠的肥胖指數和胰島素水平降低,HDL-C 水平升高,LDL-C、TC 和TAG 水平降低。已有研究[13]表明,NAFLD 患者的脂蛋白異常代謝是導致AS 進展的因素。高TAG 明顯促進游離脂肪酸(free fatty acid,FFA)水平增高,過多的FFA在肝臟沉積脂肪變性形成NAFLD[14]。長期的高胰島素血癥可加重NAFLD,促進血管內皮功能改變和平滑肌細胞功能增強,以及高凝血、低纖溶狀態的形成,加速AS的進展[15]。
在肝細胞脂肪變性進展為脂肪性肝炎(nonalcoholic steatohepatitis,NASH)、纖維化及肝硬化的過程中,炎癥反應起著重要的作用[16]。先前研究[17-18]表明多種化合物通過抑制NF-κB 途徑和促炎細胞因子的產生保護肝臟免受損傷。我們的研究顯示,Psck9敲減可以緩解NAFLD 大鼠肝臟和主動脈組織病理損傷,且抑制肝臟組織細胞凋亡。Psck9干擾還可抑制外周血中IL-1β、IL-6和iNOS的水平,進而緩解NAFLD 大鼠的炎癥反應。我們還進一步發現,Psck9敲減可阻礙免疫炎癥相關通路蛋白TLR4和NF-κB P65的活化及炎癥因子TNF-α的表達,表明Psck9可能部分通過TLR4/NF-κB P65 通路對NAFLD大鼠起到調節作用。
綜上所述,干擾Psck9基因可能通過下調TLR4/NF-κB P65磷酸化表達來緩解NAFLD 合并AS大鼠的脂質代謝異常及炎癥反應,改善肝臟及主動脈損傷。當然存在另一種可能性,即Pcsk9敲減降低了高脂飲食誘導的血脂異常,從而導致TLR4 和NF-κB P65 磷酸化表達并沒有顯著升高。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
人大建設(2019年12期)2019-05-21 02:55:32
光學精密工程(2016年6期)2016-11-07 09:07:19
海峽科技與產業(2016年3期)2016-05-17 04:32:12
