黃原膠寡糖對體外腸道菌群及其代謝產物的影響
2022-04-18 09:25:02徐靜靜孫武詹曉北張洪濤朱莉高敏杰
食品與發酵工業 2022年7期
徐靜靜,孫武,詹曉北*,張洪濤,朱莉,高敏杰
1(糖化學與生物技術教育部重點實驗室(江南大學),江蘇 無錫,214122)2(無錫格萊克斯生物科技有限公司,江蘇 無錫,214125)
人體腸道內有1014個共生微生物,它們形成人體最大的、最復雜的微生態系統,在調節宿主健康和疾病方面發揮著重要的作用[1-2]。膳食是影響腸道微生物群組成的關鍵可改變因素,膳食干預已成為調節微生物群組成和代謝活性的潛在有效策略[3]。
抗消化的寡糖(non-digestible oligosaccharides,NDOS)介于單糖和多糖之間,通常由≤10個單糖組成。近年來,越來越多的NDOS被用作益生元,調節腸道微生物結構及其代謝產物,進而改善人類健康[4-5],如低聚半乳糖、低聚果糖和低聚木糖等[6-7]。NDOS被腸道微生物群降解,可促進特定微生物的生長并產生短鏈脂肪酸;短鏈脂肪酸主要由乙酸鹽、丙酸鹽和丁酸鹽組成,它們在能量穩態[8]、腸道屏障功能[9]、脂質和碳水化合物代謝[10]中發揮重要的生理功能。
黃原膠常由油菜黃單胞菌以低成本大量生產,是美國食品和藥物管理局(Food and Drug Administration,FDA)批準的安全的食品添加劑。黃原膠寡糖是潛在的可商業化的寡糖,之前的研究通過黃原膠酸水解制備得到純度為94%的黃原膠寡糖,黃原膠寡糖由葡萄糖、甘露糖和葡萄糖醛酸組成,摩爾比為42.72∶36.73∶19.47,其聚合度范圍為2~8。該黃原膠寡糖具有抵抗模擬唾液和胃腸液消化的能力,可以穩定轉運到大腸[11]。在3個健康人的混合糞菌的體外發酵中黃原膠寡糖可增加產丁酸相關細菌的豐度和短鏈脂肪酸的水平[11-12],然而其對個體的腸道菌群及其代謝產物的動態調控過程仍不清楚。腸道反應器通過動態地模擬人體腸道的微生物發酵環境而逐漸引起關注。本實驗室自主研發了新型模擬胃腸道反應器[13],該胃腸道反應器的構造情況、控制系統以及其功能特性已驗證了進行動態發酵的可行性[14]。本研究通過3個健康人糞便微生物的體外靜態分批發酵和1個健康人糞便微生物的體外胃腸道反應器動態發酵共同探究黃原膠寡糖對糞便微生物結構及其代謝產物的影響。
1 材料與方法
1.1 實驗材料
黃原膠寡糖,本實驗室提供[11];果寡糖,Sigma公司;TIANamp糞便基因組DNA提取試劑盒,天根生化科技有限公司;Silica gel 60 F254薄層層析板,北京安捷飛科技有限公司;其他試劑,國藥集團化學試劑有限公司。
1.2 儀器與設備
HYQX-Ⅱ厭氧培養箱,上海躍進醫療器械有限公司;7890A氣相色譜儀,美國安捷倫科技有限公司;DHG-Ⅲ鼓風干燥箱,上海新苗醫療器械制造有限公司;BGR新型模擬胃腸道反應器,本實驗室提供[13]。
1.3 實驗方法
1.3.1 薄層色譜法(thin-layer chromatography,TLC)
TLC用于評價黃原膠寡糖的降解。將發酵上清液(2 μL)點在10 cm×10 cm的薄層層析板上,然后將薄層層析板轉移到鼓風干燥箱中,在60 ℃下干燥3 min。將薄層層析板放在V(正丙醇)∶V(水)=7∶3的展開劑中展開100 min。然后將薄層層析板轉移到鼓風干燥箱中,在60 ℃下干燥3 min。將干燥的薄層層析板浸入地黃酚試劑(900 mg地黃酚、25 mL水、375 mL乙醇、50 mL濃硫酸)中染色,并立即在105 ℃下加熱4 min顯色。
1.3.2 體外靜態糞便菌群發酵方法
新鮮的健康人糞便是從3名健康捐贈者(1名男性和2名女性,年齡在35~50歲,BMI在24.60~29.24 kg/m2)中用無菌的糞便收集管收集,然后放進厭氧產氣袋中,置于冰上。健康捐贈者在糞便樣本捐贈之前至少2個月沒有服用益生菌或益生元。將這3個新鮮的糞便分別用滅菌的0.1 mol/L PBS稀釋以獲得10%質量分數的糞便漿液,并在無菌條件下通過4層紗布海綿過濾以除去食物殘留物。基礎營養培養基(g/L)包含NaCl 0.1,K2HPO40.04,KH2PO40.04,NaHCO32.0,L-半胱氨酸鹽酸鹽 0.5,膽鹽 0.5,MgSO40.01,CaCl20.01,血紅素 0.025,維生素K 0.002,蛋白胨 2.0,酵母提取物 2.0,刃天青0.001,Tween 80 1 mL。基礎營養培養基用作空白對照;將果寡糖以0.5%的質量分數添加到基礎營養培養基中作為陽性對照組;將黃原膠寡糖以0.5%的質量分數添加到基礎營養培養基中。然后在8 h內將3名捐贈者糞便漿液分別以體積分數10%接種到以上培養基中。每組在3個平行容器中于37 ℃厭氧培養箱中孵育48 h。發酵液以12 000 r/min離心3 min,細菌沉淀用于檢測菌群變化,上清液用于檢測乙酸,丙酸和丁酸變化。
1.3.3 體外動態胃腸道反應器發酵方法
用無菌糞便收集管采集1名健康捐贈者(1名男性,年齡為45歲,BMI為27.86 kg/m2)的糞便,然后放進厭氧產氣袋中,置于冰上。健康捐贈者在糞便樣本捐贈之前至少2個月沒有服用益生菌或益生元。將新鮮糞便用滅菌的0.1 mol/L PBS稀釋以獲得10%質量分數的糞便漿液,并在無菌條件下通過4層紗布海綿過濾以除去食物殘留物。基礎發酵培養基(g/L)成分為:淀粉5,阿拉伯半乳聚糖 1.0,果膠 2.0,木聚糖 1.0,酵母提取物 3.0,胰蛋白胨 1.0,酪蛋白 2.0,L-半胱氨酸 0.5,KCl 1.0,NaCl 0.5,K2HPO40.5,KH2PO40.5,CaCl2·6H2O 0.15,MgSO4·7H2O 0.01,氯化血紅素 0.025,膽汁鹽 0.4,FeSO4·7H2O 0.005,吐溫80 1 mL,維生素混合液1 mL,pH值為5.8。其中維生素混合液(g/L)成分為:甲萘醌 1.0,D-生物素 2.0,維生素B12 0.5,泛酸 10.0,煙酰胺 5.0,對氨基苯甲酸 5.0,硫胺素 4.0。在實驗期將淀粉換成等量的5 g/L黃原膠寡糖,即為黃原膠寡糖培養基。
向腸道反應器(bionic gastrointestinal reactor,BGR)中裝入160 mL基礎發酵培養基(反應器的總體積為230 mL),用4.0和6.8的pH校準液對pH電極進行校準,然后在115 ℃條件下滅菌20 min。待反應器自然冷卻至室溫,于超凈工作臺內向基礎發酵培養基中以體積分數10%接入糞便漿液。接種后,將BGR轉移至固定位置,然后胃腸道反應器聯通循環水裝置,通過以規則的時間間隔將溫水泵入玻璃夾套和柔性軟管之間的空間,實現模擬腸道蠕動,溫水使模型內腔處于37 ℃。將pH值設置為5.8(模擬近端結腸的pH值),并通過pH傳感器結合泵入NaOH溶液(0.5 mol/L)使pH值自動調節在5.8左右。每隔8 h向BGR徐徐通入10 min N2以排出BGR中的氧氣使系統保持厭氧狀態。
BGR中糞便菌群經過16 h發酵后達到穩定階段,然后進行2 h饑餓處理使培養基中的碳源(淀粉)消耗殆盡,此時記錄為初始發酵時間(0 h)。然后以2 mL/min的速度用泵排出40 mL BGR中培養基,之后以2 mL/min的速度泵入40 mL黃原膠寡糖培養基,進入實驗期。每隔12 h補料40 mL黃原膠寡糖培養基,每隔12 h取樣,如此連續發酵48 h。發酵結束后,將得到的每個樣品12 000 r/min離心3 min,細菌沉淀用于檢測菌群變化,上清液用于檢測乙酸,丙酸和丁酸變化。使用TLC檢測發酵液上清液中黃原膠寡糖的降解。
1.3.4 短鏈脂肪酸濃度的檢測方法
通過使用氣相色譜法(gas chromatography,GC)分析短鏈脂肪酸的濃度,包括乙酸,丙酸和丁酸。將1 mL發酵液的上清液轉移至新的離心管中。將250 μL HCl溶液和1 mL乙醚添加至上清液,以終濃度為1 mmol/L 2-甲基丁酸作為內標,并將試管渦旋3 min。收集上層的有機組分并用無水硫酸鈉脫水,收集上清液并使其通過0.22 μm孔徑的過濾器。使用配備有HP-INNOWAX色譜柱的GC分析短鏈脂肪酸的濃度。烘箱溫度為60 ℃,并在4 min內升至190 ℃;進樣器溫度設置為220 ℃,檢測器溫度設置為250 ℃;將5 μL樣品以1∶20的分流比注入GC儀器,氮氣用作載氣,流速為1.5 mL/min。根據內標法計算短鏈脂肪酸的濃度。
1.3.5 糞便菌群組成的檢測方法
通過QIAamp DNA Stool Mini Kit提取細菌基因組DNA。使用通用引物338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)擴增16S rRNA基因的V3~V4區。使用TruSeq-DNA PCR-Free樣品制備試劑盒生成測序文庫,并通過MiSeq PE250平臺進行焦磷酸測序。分別使用FLASH(1.2.7版)和Qiime(1.9.1版)進行配對末端讀段的組裝和質量控制。Uparse(v7.0.1001)將具有>97%相似性的序列修剪分配給相同的操作分類單位(operational taxonomic units,OTU)。Silva數據庫(heep://www.arb-silva.de/)基于mothur算法用于注釋生物分類信息。使用R軟件進行冗余分析。
1.4 數據分析
所有實驗均重復3次,結果表示為平均值±標準偏差。使用Origin軟件作圖,采用SPSS軟件進行Tukey’s test,P<0.05表示統計學分析中差異顯著。
2 結果與分析
2.1 體外糞便菌群發酵中黃原膠寡糖的消耗
通過體外靜態分批發酵和動態的胃腸道反應器發酵,探究黃原膠寡糖對健康人腸道菌群及其代謝產物的影響。在健康人糞便菌群發酵過程中,黃原膠寡糖的降解通過TLC方法進行檢測。如圖1-a所示,在體外靜態糞便菌群發酵中,果寡糖和黃原膠寡糖經過3個健康人的糞便菌群發酵48 h后基本完全降解。在黃原膠寡糖的體外動態糞便菌群發酵中,本文呈現了BGR中的一組典型數據。如圖1-b所示,黃原膠寡糖在24~36 h的發酵過程中開始被部分降解,并且優先降解黃原膠寡糖中低分子質量的成分;黃原膠寡糖在36~48 h的發酵過程中被完全降解。以上結果表明黃原膠寡糖在糞便菌群發酵48 h后可被完全降解,并且主要在24~48 h的發酵過程中被降解。MOON等[15]指出阿拉伯低聚糖的緩慢發酵可能使得短鏈脂肪酸在整個結腸內發揮有益的健康影響。FU等[16]指出在結腸遠端發酵的碳水化合物可能是改善人類結腸遠端出現的大多數結腸癌病例的有效策略。因此,黃原膠寡糖的延遲降解可能有助于黃原膠寡糖在結腸遠端發揮作用。
2.2 黃原膠寡糖對糞便菌群主要代謝產物的影響
乙酸、丙酸和丁酸是腸道微生物發酵碳水化合物的主要終產物,在宿主健康中發揮重要的作用[17]。如圖2所示,在體外靜態糞便菌群發酵48 h后,與空白相比,黃原膠寡糖可顯著增加乙酸、丙酸、丁酸和總短鏈脂肪酸的濃度,濃度分別為31.30、11.57、5.19和48.07 mmol/L;與果寡糖相比,盡管黃原膠寡糖的丙酸和總短鏈脂肪酸產量低于果寡糖,但是黃原膠寡糖的丁酸產量顯著高于果寡糖的丁酸產量,是果寡糖丁酸產量的2.66倍。此外,與空白相比,黃原膠寡糖和果寡糖的發酵液pH值均明顯下降,其中果寡糖的pH值下降最多,這與果寡糖的總短鏈脂肪酸產量最高相對應。
如圖3所示,在0~24 h的發酵過程中,黃原膠寡糖對乙酸、丙酸、丁酸的產量影響不大;在36~48 h的發酵過程中,乙酸、丙酸、丁酸和總短鏈脂肪酸的產量和NaOH的消耗量均顯著增加,這與黃原膠寡糖在24~48 h的發酵過程中降解的結果一致。在發酵48 h時,乙酸、丙酸、丁酸和總短鏈脂肪酸的濃度分別達到49.44、28.64、1.65和79.72 mmol/L,與0 h相比,分別增加1.67、1.40、6.25和1.58倍。

a-乙酸;b-丙酸;c-丁酸;d-總短鏈脂肪酸;e-pH圖2 在體外靜態糞便菌群發酵中短鏈脂肪酸濃度和pH的變化Fig.2 Changes in short-chain fatty acid levels,and pH in vitro static batch fermentation注:值為平均值±標準偏差(n=3);*表示P<0.05(下同)
結果表明在體外靜態和動態糞便菌群發酵中黃原膠寡糖均可顯著增加乙酸、丙酸、丁酸和總短鏈脂肪酸的產量,尤其是丁酸的產量。已有研究指出具有β(1→4)糖苷鍵的二糖比具有α(1→2,1→3,1→4,1→5)和 β(1→1,1→2,1 →3,1→5)糖苷鍵的二糖產生更多的丁酸鹽[18],這暗示黃原膠寡糖發酵中丁酸產量的增加可能與黃原膠寡糖中的β(1→4)糖苷鍵相關。丁酸在糖異生和脂肪生成代謝途徑中發揮有益的作用[19]。例如丁酸可改善大鼠原代脂肪細胞中激素調節的脂質代謝和胰島素刺激的葡萄糖攝取[20]。以上結果說明黃原膠寡糖可能通過增加丁酸的產量,參與改善宿主健康。
2.3 黃原膠寡糖對糞便菌群結構的影響
通過16S rRNA測序檢測糞便菌群組成的變化。在體外靜態和動態糞便菌群發酵中,根據97%的序列相似性分別檢測到482和555個OTU。如圖4所示,在體外靜態和動態糞便菌群發酵中,擬菌門(Bacteroidota)、厚壁菌門(Firmicutes)和變形菌門(Proteobacteria)為優勢菌門。在體外靜態糞便菌群發酵中,與空白相比,黃原膠寡糖可降低糞便菌群中Firmicutes/Bacteroidota的比例,黃原膠寡糖的動態發酵呈現相似的降低的Firmicutes/Bacteroidota的比例(圖5)。多個研究表明[21],肥胖動物和人類的腸道微生物群表現出更高的厚壁菌門/擬桿菌門比率,建議將該比率作為潛在的生物標志物,不過這仍需更多的證據。以上結果暗示黃原膠寡糖可能通過降低腸道菌群中Firmicutes/Bacteroidota的比例改善宿主腸道菌群結構。
體外靜態糞便菌群發酵中,在門水平和屬水平上,果寡糖和黃原膠寡糖發酵后的3個不同健康人來源的糞便菌群呈現差異的變化(圖4和圖6)。在屬水平,與空白和果寡糖相比,黃原膠寡糖在3個不同健康人的糞便菌群發酵中可同時增加Parabacteroides屬和Hungatella屬的相對豐度。此外,通過lefse分析果寡糖和黃原膠寡糖中顯著變化的菌群,發現與果寡糖相比,黃原膠寡糖可顯著增加Parabacteroides屬、Hungatella屬和Lachnoclostridium屬的相對豐度。

a-靜態;b-動態圖6 在體外靜態和動態糞便菌群發酵中屬水平上主要的10個菌群的相對豐度變化Fig.6 Relative abundance of dominant genera(top 10) in in vitro static batch fermentation and dynamic BGR fermentation
此外,與果寡糖相比,黃原膠寡糖可顯著增加科水平上毛螺菌科(Lachnospiraceae)的相對豐度。毛螺菌科(Lachnospiraceae)和Lachnoclostridium屬被認為是腸道中產生丁酸鹽的細菌[16],在黃原膠寡糖的體外糞便菌群發酵中,毛螺菌科(Lachnospiraceae)和Lachnoclostridium屬的富集可能與丁酸產量的顯著增加相關。
體外動態糞便菌群發酵中,與0和24 h相比,黃原膠寡糖在48 h發酵時可極大的增加屬水平上Parabacteroides、Lachnospira(屬于Lachnospiraceae)和UBA1819的相對豐度。狄氏副擬桿菌(Parabacteroidesdistasonis)是Parabacteroides屬的主要成員之一,可通過在腸道中生產琥珀酸和次級膽汁酸減輕小鼠肥胖和代謝功能障礙[22]。毛螺菌科(Lachnospiraceae)被認為是包含產丁酸菌的重要的一個科,Lachnoclostridium屬和Lachnospira屬的產丁酸菌也屬于毛螺菌科(Lachnospiraceae)[16]。
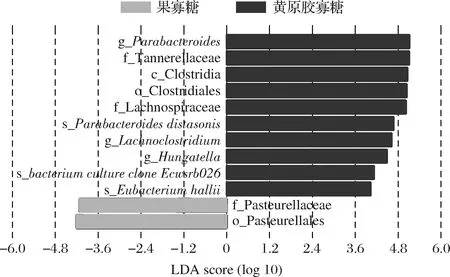
圖7 LEfSe分析在體外靜態糞便菌群發酵中顯著變化的菌群(LDA score>4.0)Fig.7 LEfSe was used to analyze significantly differential taxa(LDA score>4.0) in vitro static batch fermentation
產丁酸菌在人類健康中起著至關重要的作用,包括為腸道上皮細胞提供能量、維持腸道細菌平衡和調節宿主細胞反應[23]。以上結果表明,在體外靜態和動態糞便菌群發酵中,黃原膠寡糖可明顯富集毛螺菌科(Lachnospiraceae)和Parabacteroides屬,進而改善腸道菌群結構。
3 結論
本文通過體外靜態和動態糞便菌群發酵評價黃原膠寡糖的益生元潛能。在體外靜態糞便菌群發酵中,48 h時,黃原膠寡糖基本被完全降解,顯著增加乙酸、丙酸、丁酸和總短鏈脂肪酸的產量(P<0.05);與果寡糖相比,黃原膠寡糖表現出較強的產丁酸能力。在體外動態糞便菌群發酵中,黃原膠寡糖主要在24~48 h的發酵過程中被降解并顯著增加乙酸、丙酸、丁酸、總短鏈脂肪酸的產量和NaOH的消耗量(P<0.05)。在體外靜態和動態糞便菌群發酵中,黃原膠寡糖均可降低Firmicutes/Bacteroidota的比例,增加毛螺菌科(Lachnospiraceae)和Parabacteroides屬的相對豐度,進而參與改善宿主腸道菌群結構。研究結果表明黃原膠寡糖具有一定的益生活性。