鳥類物種組合模式的穩定性
——以珠海淇澳自然保護區紅樹林鳥類群落變化為例
2022-04-16 09:09:06葉錦玉虞皓琦廖寶文游奕來劉新科林壽明楊道德胡慧建
生態環境學報 2022年2期
葉錦玉 ,虞皓琦,廖寶文,游奕來,劉新科,林壽明,楊道德,胡慧建*
1. 中南林業科技大學野生動植物保護研究所,湖南 長沙 410004;
2. 廣東省科學院動物研究所/廣東省動物保護與資源利用重點實驗室/廣東省野生動物保護與利用公共實驗室,廣東 廣州 510260;3. 中國林業科學研究院熱帶林業研究所,廣東 廣州 510520;4. 廣東珠海淇澳-擔桿島省級自然保護區管理處,廣東 珠海 519000;5 廣東省林業調查規劃院,廣東 廣州 510520
群落構建規則(community assembly rules)是生態學研究的核心和基礎問題,也是恢復生態學的重要研究領域(Brown et al.,1991;Temperton et al.,2004;G?tzenberger et al.,2012)。群落的物種組合模式從不同側面反映了群落構建的特點,是其重要研究內容之一(Brown et al.,1991)。許多研究表明,動物群落的物種組合具有多種相對固定的模式,如種面積效應(Brown et al.,1991;Connor et al.,2001)、物種個體大小頻次偏正態分布(Brown et al.,1991)、物種多度頻次偏正態分布(Brown,1995),以及種-屬數量間強相關關系(Blackburn et al.,1999;Gaston,2000;胡慧建等,2003)和屬含物種的頻次分布可乘冪模型擬合(馮永軍等,2006;鄭孜文等,2008;連軍豪等,2011)等。這些模式的存在可能意味著群落的構建具有穩定性。
植被群落是動物棲息地的重要組成部分,本地植被群落的改變會導致動物棲息地和食物變化,從而導致物種發生重組(Davidsdottir et al.,2016;Lindbladh et al.,2019)。若物種重組后其物種組合模式仍與重組前一致,則說明群落的物種組合模式具有可重復性,這對解釋物種共存和物種多樣性的維持有重要作用。目前植被變化對動物群落產生影響的相關研究主要集中在物種數量(江海聲等,1998)、物種組成(Isacch et al.,2005;Fontaine et al.,2009;Nkwabi et al.,2010;張國鋼等,2008)、多樣性指數(Fontaine et al.,2009;Nkwabi et al.,2010;Curado et al.,2013)等,而對群落的種屬關系、個體大小等物種組合模式變化的影響研究仍存在空白。研究植被變化前后動物群落是否具有相同或類似的物種組合模式,或存在其他明顯的群落構建規則,是驗證動物群落構建可重復性的重要依據,也可對動物群落的保護與修復提供有力的理論支撐。
廣東珠海淇澳-擔桿島省級自然保護區的淇澳島片區曾是原生紅樹林重要的分布地,但由于大量圍海造田、圍墾養殖以及橋梁、碼頭建設等,逐漸演變為以互花米草(Spartina alterniflora)為優勢的植被群落。為解決紅樹林面積萎縮和互花米草入侵問題,自 2001開始在淇澳島試種以無瓣海桑(Sonneratia apetala)為主的紅樹林植被,如今,紅樹林已替代互花米草成為該區域的優勢植被(廖寶文等,2008)。本研究利用淇澳島紅樹林恢復前后的2次鳥類調查數據(恢復前于2001年3月—12月,恢復后于2015年10月—2016年6月),分析區域植被發生變化后鳥類物種及其組合模式的變化,探討鳥類群落構建中物種的組合模式,為動物群落的恢復提供新視角。
1 研究方法
1.1 研究地點
廣東珠海淇澳-擔桿島省級自然保護區成立于2004年,由珠海市的淇澳島和擔桿島兩個片區組成,總面積7373.77 hm2。該區域位于珠江口,屬海灣地貌,主要有丘陵、淺海沉積(灘涂)的地貌等類型;地處北回歸線附近,南亞熱帶海洋性季風氣候,年均氣溫22—23 ℃之間,年平均降雨量1975.1 mm。本研究于淇澳島內,范圍以植被覆蓋最邊緣處為界,主要位于海堤兩側中潮位以上,面積約 903 hm2, 四至邊界為 113°36′40″— 113°39′15″E ,22°23′40″—22°27′38″N。
上世紀 80年代初,該區域內有紅樹林面積112.2 hm2,樹種以秋茄(Kandelia obovata)和桐花樹(Aegiceras carniculatum)為主。1990年以后,由于圍海造地、人為砍伐和互花米草入侵等危害,紅樹林面積僅存32 hm2。保護區于1999年和2000年嘗試小規模種植以無瓣海桑為主的紅樹林,面積約 1 hm2;2002年后開始大量種植(2001年無種植),最大面積達500 hm2,占淇澳島片區總面積的9.80%,約占潮間帶面積的50.00%(圖1)。現紅樹林生態系統自 2008年開始趨于穩定,有紅樹植物32種,真紅樹20種,半紅樹12種,其中栽培種有21種(廖寶文等,2008)。

圖1 珠海淇澳島紅樹林及樣線樣點分布示意圖Figure 1 Location of mangrove and transect in Qi’ao Island, Zhuhai, Guangdong
2 調查方法
第1次調查在紅樹林大面積恢復前(下簡稱恢復前),時間為2001年3月31日—4月1日、7月8—9日、10月12—13日和12月26—27日,此時調查范圍內植被以互花米草為主。第2次調查在紅樹林恢復后(下簡稱恢復后),時間為2015年10月29—30日和12月24—25日、2016年3月29—30日和6月19—20日,此時的調查范圍內已形成以紅樹林為主的植被群落(廖寶文等,2008)。
2次鳥類調查的樣線和樣點相同,調查選擇天氣良好情況下進行,每日調查時間為07:00—11:00、15:00—17:00,中午鳥類活躍性較低,不進行調查。分別在該片區西邊海堤和東邊棧道各有1條樣線,長度分別為2.5 km和4.5 km,樣線單側寬度為100 m;東邊海堤高處有1個樣點,半徑寬200 m(圖1)。鳥類鑒定參照約翰·馬敬能等(2000)所著《中國鳥類野外手冊》,分類系統、區系和居留型等參照《中國鳥類分類與分布名錄 (第2版)》(鄭光美,2011)。
3 數據處理
3.1 物種累積曲線
采用物種累積曲線(species accumulative curve)判斷2次鳥類調查的充分性(李巧,2011),使用R語言vegan程序包計算物種累計曲線。如果累積曲線為直線上升,表明抽樣量不足,需要增加抽樣量;如果曲線在急劇上升后變為漸近線或增速趨勢放緩,則說明抽樣充分。恢復后調查記錄了物種個體數,因此直接計算物種累積曲線,并判斷其抽樣充分性。
由于恢復前調查記錄的為數量等級,即“+”<5只,“++”為5—99只,“+++”為100—500只,“++++”為>500只,因此用數量等級的最小值計算物種累積曲線,即物種數量等級為“+”則該物種數量換算為1只,“++”換算為5只,“+++”換算為100只,“++++”換算為500只。換算后計算得出的物種累積曲線在急速上升后變為漸近線(圖2),因此認為恢復后調查抽樣充分。

圖2 2次調查鳥類物種累積曲線Figure 2 Species accumulation curve of birds survey before and after mangrove restoration
3.2 物種組成
對2次調查的鳥類名錄進行分析,對比物種豐度、區系、居留型、季節差異及保護物種等。統計2次調查全部、共有物種以及單次調查記錄的區系(東洋界、古北界和廣布種)、居留型(留鳥、夏候鳥和冬候鳥)、季節、生態類型(涉禽、游禽、猛禽、陸禽、攀禽和鳴禽)的物種數,用卡方檢驗判斷其差異的顯著性。本節及下節均采用統計軟件IBM SPSS Statistics 25對數據進行分析。
3.3 種屬結構關系
動物的屬和種在數量上具有顯著的相關性(Connor et al.,2001),屬含物種數分布頻次隨物種數增加而減少,暨1種/屬最多,2種/屬次之,3種/屬更少……(Brown,1995),且顯示出明顯的規律性,可用乘冪模型擬合(馮永軍等,2006)。故本研究對比以上兩種關系在2次調查中的差異,以觀察其中分類結構的變化。
3.3.1 種-屬數量關系
分別統計2次調查按春至冬季節順序累計的物種數和屬數,對種-屬數量進行相關關系分析。采用K-S方法對數據進行分布型檢驗,凡符合正態分布的數據,采用Pearson線性相關分析,否則采用Spearman秩相關分析。采用協方差分析判斷恢復前后種-屬數量的回歸直線是否具有一致性。
3.3.2 屬含物種數的頻次分布
分別統計2次調查具有相同物種數的屬的頻次,暨統計1種/屬、2種/屬、3種/屬……出現的數量,并進行K-S方法對數據進行分布型檢驗,發現不符合正態分布。故根據其數據分布規律,采用乘冪模型進行擬合,再進行ln轉換為線性模型,采用協方差分析該模型的一致性,以判斷恢復前后屬含物種數的乘冪模型的一致性。
3.4 體長結構
采用馬敬能等著《中國鳥類野外手冊》(約翰·馬敬能等,2000)中鳥類物種體長數據對 2次調查名錄中的鳥類體長進行統計,并進一步以10 cm為間隔分別統計2次調查名錄中鳥類物種體長的頻次分布。2次調查的鳥類全部和單次調查記錄物種的體長頻次分布的差異均采用卡方檢驗進行分析。
4 結果
4.1 物種累積曲線
恢復前調查的鳥類物種數量等級數據通過數量等級的最小值換算后,物種累積曲線在快速上升后變為一漸近線;恢復后調查的物種累積曲線亦為先快速上升后增速放緩,說明2次調查取樣均較充分(圖2)。
4.2 物種組成
2次調查共記錄到鳥類99種,隸屬于13目35科70屬。其中,恢復前調查共記錄鳥類11目29科58屬78種;恢復后調查共記錄鳥類12目33科57屬80種,2次調查的鳥類物種豐度基本一致。2次調查共記錄鳥類有東洋界 44種(占總數的44.44%),古北界31種(占31.31%),廣布種24種(占24.24%);留鳥51種(占52.04%),冬候鳥37種(占37.76%),夏候鳥10種(占10.20%)(表1);共有鳥類物種有59種(表2),分別占2次調查的76.62%和73.75%,主要為涉禽類和鳴禽類(附表1)。

表1 紅樹林恢復前后鳥類區系和居留型Table 1 Fauna and residence types of birds survey before and after mangrove restoration

表2 紅樹林恢復前后各季節鳥類物種數Table 2 Number of bird species in different seasons before and after mangrove restoration
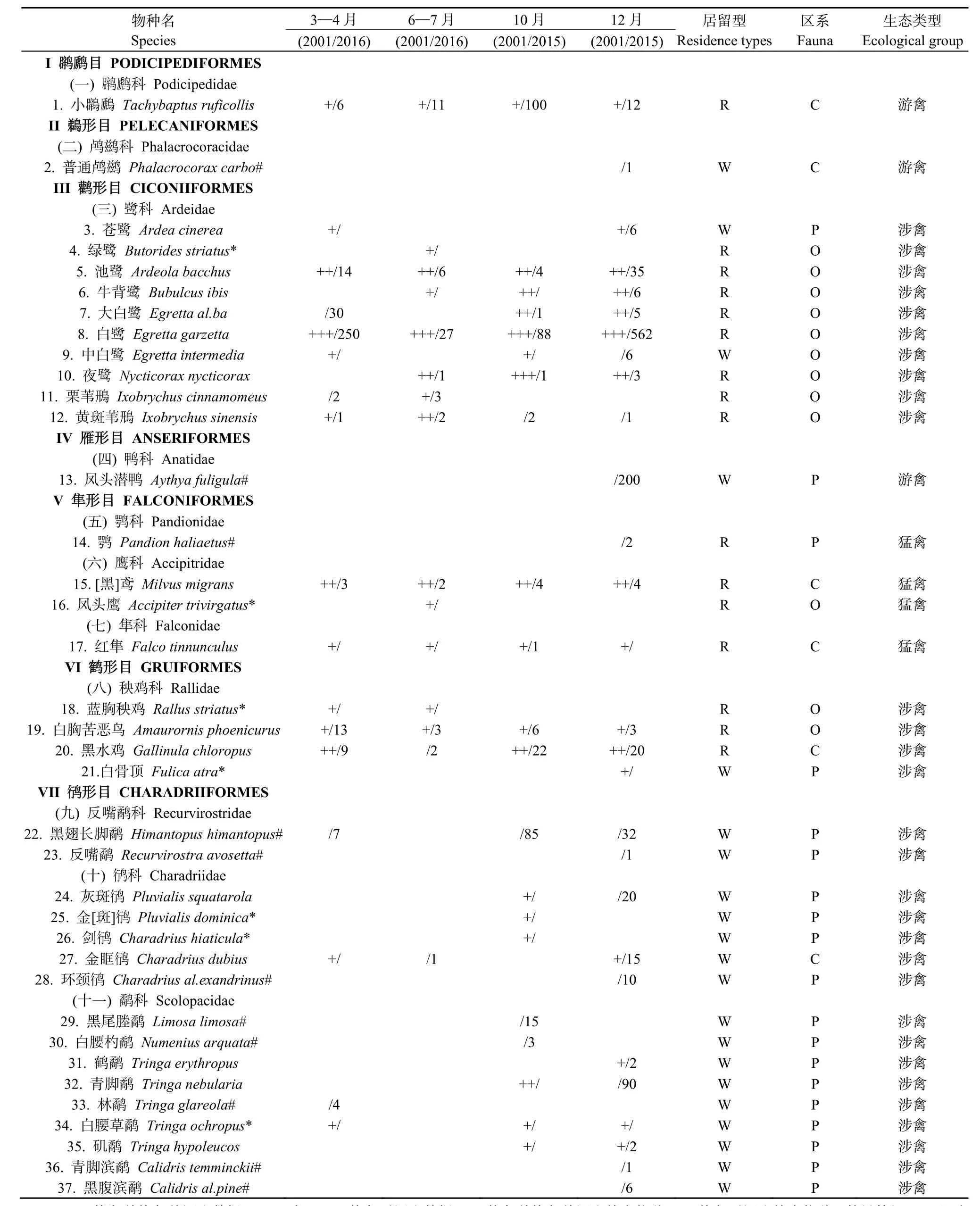
表1 紅樹林恢復前恢復前和恢復后不同季度鳥類種類與數量變化組成Appenddix 1 Variety and form of bird species and richness in each season before and after vegetation change

續附表1 紅樹林恢復前恢復前和恢復后不同季度鳥類種類與數量變化組成Continued appenddix 1 Variety and form of bird species and richness in each season before and after vegetation change

續附表1 紅樹林恢復前恢復前和恢復后不同季度鳥類種類與數量變化組成Continued appenddix 1 Variety and form of bird species and richness in each season before and after vegetation change
恢復前特有物種有5科17屬18種,占其總數的23.38 %,主要生態類型為鳴禽、涉禽和猛禽;恢復后特有物種有 6科 12屬 21種,占其總數的26.25 %,主要為涉禽和鳴禽(附表1)。恢復前與恢復后在區系(P=0.657)、居留型(P=0.638)和生態類型(P=0.151)上均無顯著性差異。
4.3 季節變化
在4個季節中,恢復前后的鳥類物種組成發生了部分替代,約占總數的24%,以候鳥為主。2次調查在4個季節的物種豐富度、區系及居留型上均無顯著差異(表3)。

表3 紅樹林恢復前后各季度鳥類的區系及居留型Table 3 Fauna and inhabitation pattern in different seasons of birds surveyed before and after mangrove restoration
4.4 分類階元結構
4.4.1 種-屬數量關系
2次調查的鳥類的物種-屬數量均呈正相關關系,且能用線性方程進行擬合:恢復前為y=0.5596x+15.093(r2=0.990,P=0.001);恢復后為y=0.5215x+15.568(r2=0.990,P=0.002)(圖3),協方差分析表明 2次調查的種-屬間數量的回歸直線平行,具有一致性(P=0.234)。

圖3 紅樹林恢復前后鳥類屬數量(y)與物種數(x)的線性模型Figure 3 Linear model of species-genera richness of birds surveyed before and after mangrove restoration
4.4.2 屬含物種數的頻次分布
2次調查屬含物種數的頻次分布均隨著每屬所含物種數增加呈遞減關系,能用乘冪模型進行擬合:恢復前,y=51.65x?2.7(r2=0.984);恢復后,y=48.47x?2.49(r2=0.937)(圖4a),協方差分析表明2次調查的屬含物種數的頻次分布的乘冪擬合曲線平行且具有一致性(P=0.688)。2次調查共有種的屬含物種數頻次分布亦可用乘冪模型進行擬合,單次調查記錄物種的均為單種屬占比較高(圖4b)。

圖4 紅樹林恢復前后鳥類屬含物種數的頻次分布Figure 4 Frequency distribution of number of species in genus before and after mangrove restoration
4.5 體長頻次分布
2次調查的體長頻次分布有相似,體長均以11—20 cm的最多,分別有27種和26種;體長為1—10 cm的鳥類物種數量均僅有4種;體長大于21 cm的體長組的物種數逐漸減少(圖5a)。2次調查的體長分布均呈正偏態分布,卡方檢驗結果表明2次調查鳥類各體長組的頻次分布無顯著性差異(P=0.992)。共有種和特有種的體長頻次分布皆的無顯著差異(P=0.957)(圖5b)。

圖5 紅樹林恢復前恢復前后鳥類體長頻次分布Figure 5 Frequency distribution of body size of species surveyed before and after mangrove restoration
5 討論
5.1 植被變化對鳥類群落的影響
2次調查共發現鳥類99種,紅樹林恢復前記錄到78種,恢復后調查到80種;恢復前特有的有19種,恢復后則有21種,約24%的物種發生變化。植物不僅是鳥類所有食物的最終來源,還為鳥類提供了巢址、巢材、棲木和隱蔽等各種必需的生存條件(鄭光美,2012),某植被類型或某幾種植被還可能與某些鳥類形成密切依存關系(Nkwabi et al.,2010;鄭光美,2012)。紅樹林恢復后,棕扇尾鶯(Cisticola juncidis)、紅尾水鴝(Rhyacornis fuliginosus)、劍鸻(Charadrius hiaticula) 和金鸻 (Pluvialis dominica)等在水邊灌叢或空地上覓食的物種消失,而藍磯鶇(Monticola solitaries)、灰背鶇(Turdus hortulorum)和叉尾太陽鳥(Aethopyga christinae)等在林間活動的鳥類,及青腳濱鷸(Calidris temminckii)、黑翅長腳鷸(Himantopus himantopus)和黑尾塍鷸(Limosa limosa)等在較深水域覓食的物種出現。這與淇澳島開始大面積人工種植無瓣海桑后,部分生境發生了變化,灘涂區面積減少,紅樹林區面積增加(廖寶文等,2008)有關。
5.2 鳥類群落物種組合模式的穩定性
動物群落的物種豐度具有種-面積效應,即物種豐度和區域面積呈正相關(Arrhenius,1921;唐志堯等,2009)。由此可推斷當區域面積一定時,群落中的物種豐度可能保持相對穩定。但目前少有研究對比同區域下環境變化前后的動物群落變化,并對此推斷進行驗證。江海聲等(1998)亦發現海南南灣半島在植被破壞前(1950年)和恢復后(1990年)記錄的鳥類物種豐度基本保持一致,分別為54種和53種,但未對此現象進行分析。本研究發現淇澳島大面積種植紅樹林前后鳥類物種豐度分別為78種和80種,與南灣半島的鳥類調查結果(江海聲等,1998)類似,均符合種-面積效益的推斷。但混合植被生境可能是此推斷成立的條件,因為專性鳥類對植被的依賴性較強,在單一植被生境下差異較大,但在混合植被生境中專性鳥類豐度差異不顯著(Ma et al.,2011)。本研究區域與海南南灣半島(江海聲等,1998)均為混合多種植被生境的區域,為鳥類提供了多樣化的生境,弱化了一些專性鳥類對群落的影響。因此,研究認為在區域面積不變,且為非單一植被生境時物種豐度可能保持相對穩定。
研究發現紅樹林恢復前恢復前后2次調查的種和屬在數量上呈正相關關系,屬含物種數頻次關系有1屬1種頻次最高,1屬2種顯著減少,以此類推的趨勢并可用乘冪曲線擬合,均與馮永軍等(2006)和鄭孜文等(2008)分別對中國鳥獸的研究結果相似。說明種和屬不僅在大尺度上有強相關性,在小區域尺度上也有此規律。胡軍華等(2008)發現長江中游不同湖泊的魚類在 1950s、1980s和2000s 3個時期的種-屬數量及屬含物種數頻次分布也有此規律,但未檢驗其擬合曲線的一致性。本研究通過協方差分析發現,2次調查的種-屬數量關系和屬含物種數分布頻次均無顯著差異(P>0.05),且南灣半島植被破壞前和恢復后鳥類物種(江海聲等,1998)的屬含物種數頻次分布也有一致性(圖6a),說明植被變化前后鳥類群落可在種-屬結構上保持一致。

圖6 南灣半島植被變化前后鳥類屬含物種數頻次和體長頻次分布Figure 6 Frequency distribution of number of species in genus and body size before and after vegetation change in Nanwan Peninsula
動物個體大小通過異速生長關系與物種的形態、生理和行為等密切相關(Blackburn et al.,1999)。有研究表明,個體大小相近的物種間的競爭也較強,物種個體大小分布頻次分布具有規律性(Stanley,1973;Koz?owski et al.,2002;Allen et al.,2006),在大尺度上多呈正偏態分布(Blackburn et al.,1999;Clauset et al.,2009)。
本研究結果表明,紅樹林恢復前后的鳥類體長均呈正偏態分布,與前人的研究結果一致;且協方差分析表明,物種體長頻次分布無顯著性差異(P=0.893)。同樣對南灣半島的鳥類體長頻次分布進行分析發現,植被破壞前和恢復后的鳥類的體長頻次分布相似(圖6b),且卡方檢驗結果表明其分布無顯著差異(P=0.923)。說明鳥類群落的體長頻次分布具有一定規律,并在植被變化前后可保持相對穩定的結構。2次調查記錄的鳥類物種在區系、居留型、季節、生態類型和體長組上的頻次分布,以及各季節的不同區系、居留型的物種數均無顯著差異(表3),也進一步說明紅樹林恢復前后鳥類群落有相似的物種組合模式。
有研究表明,植被變化后部分新進入的鳥類與消失的鳥類存在替代關系。Nkwabi et al.(2010)發現當塞倫蓋蒂平原的草原由長草變為短草時,2種云雀在功能上存在替代關系。從卡方檢驗結果來看,2次調查中單次調查記錄鳥類在體長頻次分布、生態類型、區系和居留型上均無顯著差異,但除斑魚狗(Ceryle rudis)與藍翡翠(Halcyon pileate)外,其余物種在體長組、區系、居留型和生態類型無對應關系。研究還發現2次調查共有種的屬含物種數與體長頻次分布均與總體的有較高相似性,說明紅樹林恢復前恢復前后鳥類物種的組合模式可能受其共有物種的影響較大。
目前,尚未有研究報道環境變化后物種組合模式的可重復性及其形成機制。本研究發現2次調查的鳥類群落的屬含物種數頻次分布、體長頻次分布均符合前人在大陸和局域尺度的研究,且在植被變化前后無顯著差異。從群落構建的生態位理論出發,同屬物種在形態和生態上均具有較高相似性(鄭光美,2012),在環境篩選下進入同一區域,又因競爭排除法則(Gause,1934),即生態上相似的兩個物種在同一條件下不能共存,使得過于相似的物種發生競爭排除,從而使各物種間的相似性受到限制,即群落共存物種間的極限相似性(limiting similarity)(Wilson et al.,1995)。這可能導致了群落中單種屬最多,也是其體長分化的原因(Blackburn et al.,1999)。但同時一些物種由于對環境和資源的適合度不同而導致生態位的分化(Chesson,2000),令某些屬或體長組內可共存多個物種。由此可推測物種經擴散到達某一區域后,在環境篩選和屬內物種的極限相似這兩個反作用力的共同推動下(Webb et al.,2002),形成了現有物種組合模式。調查區域植被變化未對這些模式造成顯著性影響,可能說明其影響因子在于當地較為穩定的環境因子,如光照、溫度和降水等。
5.3 物種組合模式的應用前景
目前生物群落構建的研究多集中群落構建的機制中的生態位理論(包含環境過濾和生物相互作用)(Clements,1916;Gleason,1926;Hubbell,2011)和中性理論(主要為擴散機制)(Hubbell,2011),且通常認為在不同尺度上起主要作用的構建機制不同(G?tzenberger et al.,2012)。相比群落構建機制,群落的物種組合模式在不同尺度上均具有類似的規律,且其研究對象為物種的分類階元、個體大小、生態類型和區系等變量,數據容易獲取,且均與物種形態和覓食、繁殖行為等生態特征密切相關,能更高效直觀地反映了動物群落的構建規則,因此在目前具有更高的應用前景。
本研究發現在植被發生變化后,鳥類群落仍依照變化前的屬含物種數頻次分布、體長頻次分布和生態類型頻次分布等物種組合模式進行構建。這些物種組合模式有望作為判斷鳥類群落完整性的重要依據,如通過鳥類物種數量(Fang et al.,2020;徐正春等,2019)、屬含物種數頻次分布及體長分布等數據的變化,判斷鳥類群落的穩定性和完整性。其中屬含物種數的頻次分布模型在動物群落恢復上有較高的應用價值:一方面可用于評價恢復效果,可模型的符合程度來評價群落是否已穩定;一方面在群落未達到穩定的情況下,可計算未穩定群落與穩定群落模型的差值來確定缺失的相關物種。
6 結論
本文研究結果驗證了:鳥類群落在當地植被變化后其物種組成發生變化,但在物種豐富度和群落物種分類結構、體長頻次分布、居留型、區系組成和生態類型上均保留了原有特征。故本研究認為區域內動物群落可依照固定的物種組合模式進行構建,具有可重復性。
致謝:感謝珠海市林業局、廣東珠海淇澳-擔桿島省級自然保護區巡護隊給予的支持和幫助;感謝廣東省科學院動物研究所的曹宏芬、劉金成等參與調查,以及胡一鳴博士對本文摘要的審閱修改。
猜你喜歡
歌海(2024年6期)2024-03-18 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
兒童故事畫報·自然探秘(2022年3期)2022-04-27 00:17:10
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
瘋狂英語·初中天地(2018年6期)2018-11-24 02:39:24
汽車觀察(2018年10期)2018-11-06 07:05:26
