沙棘根瘤內生菌株庫構建與微生物多樣性分析
2022-04-08 12:21:16魏繼華李佳益張建國羅紅梅何彩云
浙江農林大學學報 2022年2期
關鍵詞:物種
魏繼華,李佳益,劉 宏,張建國,2,羅紅梅,何彩云
(1.中國林業科學研究院 林業研究所 國家林業和草原局林木培育重點實驗室,北京 100091;2.南京林業大學南方現代林業協同創新中心,江蘇 南京 210037;3.中國林業科學研究院 沙漠林業實驗中心,內蒙古 磴口 015200)
沙棘Hippophae rhamnoides又名醋柳,是胡頹子科Elaeagnaceae沙棘屬Hippophae的落葉性灌木[1]。作為藥食同源植物的沙棘不僅在食療、醫藥、農林牧漁等領域具有較大的經濟價值,在水土保持、恢復生物鏈及防風固沙中也具有極大的生態價值[2-5]。生長過程中沙棘根部會遭受土壤中放線菌、細菌的侵染形成根瘤。部分菌種會在根瘤中高度富集發揮固氮、促生長、抵御逆境脅迫、防止有害病菌侵染等功能[6-8]。傳統的微生物研究方法主要以培養基進行分離純培養,再進而探究其培養特征、顯微結構、生理特性等[9]。而自然界中90%以上的微生物為不可培養微生物,且現有培養基與培養技術不適應未知菌群的生長,或部分菌群生長緩慢、豐度較小等情況都會對菌群的多樣性評估產生影響[10]。以二代高通量測序為基礎的16S rDNA技術通過對編碼原核核糖體小亞基rRNA的DNA序列進行測序,不僅克服了傳統方法難以獲得不可培養菌株的弊端,還能對樣品中的物種相對豐度進行排序,并分析各群組樣品中發揮重要作用的優勢物種,解析樣品中微生物之間的相互作用。該技術對研究沙棘根瘤內生菌微生物多樣性與環境關系以及微生物資源的開發利用有重要的理論和現實意義[11-16]。
本研究通過16S rRNA測序技術對沙棘根瘤內生菌進行物種注釋、分類學分析、α多樣性分析、β多樣性分析、組間差異顯著性分析,比較高通量測序和純培養方法的差異與優劣,為發掘具有應用價值的根瘤內生菌資源提供科學依據。
1 材料與方法
1.1 材料采集
采樣地為內蒙古自治區巴彥淖爾市磴口縣中國林業科學研究院沙漠林業實驗中心試驗林場(40°29′34″N,106°74′06″E)。該研究區海拔為 1 054 m,年平均氣溫為 7.4 ℃。2020 年 7 月,選取人為干擾因素較少的沙漠邊緣地帶采集沙棘根瘤樣品。在每個樣地10 m×10 m的區域內用網格法定9個點,運用梅花形采樣法在邊角及中心共5個點分別采集根瘤樣品并進行混合,共設計6組重復樣,分別命名為M1、M2、M3、M4、M5、M6。
1.2 沙棘根瘤內生菌的分離培養
1.2.1 沙棘根瘤內生菌的分離 依據文獻[17-18]的方法進行修改,使其更加適宜沙棘根瘤內生菌的分離。詳細步驟如下:選取新鮮飽滿的根瘤,沖洗掉土粒泥沙,將根瘤團用解剖刀分割成帶有單柄的瘤瓣,用紗布包裹,先用體積分數為95%的酒精溶液浸泡30 s,再用體積分數為10%的次氯酸鈉溶液表面滅菌5 min,取出后用無菌水沖洗數次。在滅菌濾紙上,用無菌解剖刀先切取根瘤頭部,再將其均分成2~3份薄片,置于固體培養基中28 ℃恒溫暗處靜置培養。根據相關研究,本研究選取BAP[19]、S[20]、JA[19]、高氏一號培養基[19]進行分離培養。
1.2.2 沙棘根瘤內生菌的鑒定 提取純培養的沙棘根瘤內生菌DNA后,對16S rDNA全長進行PCR擴增。序列引物采用YU等[21]設計的細菌通用引物(引物序列27F:5′-AGAGTTTGATCMTGGCTCAG-3′,1492R:5′-GGYTACCTTGTTACGACTT-3′),PCR 總反應體系為 50 μL,包括 10×緩沖液 (KOD buffer)5 μL、2 mmol·L-1三磷酸脫氧核糖核苷酸混合液 (dNTPs) 5 μL、基因組 DNA (genomic DNA) 1 μL、上游引物 (forward primer) (10 μm) 1 μL、下游引物 (reverse primer) (10 μm) 1 μL、DNA 聚合酶 (KOD DNA polymerase) 1 μL、超純水 (ddH2O) 36 μL。PCR 反應程序:94 ℃ 預變性 3 min,94 ℃ 變性 30 s,58 ℃ 退火 30 s,72 ℃延伸1 min,35個循環,最后72 ℃延伸10 min。用質量分數為1%的瓊脂糖凝膠電泳,確定有特異擴增后,進行PCR產物回收和測序注釋,并參考文獻[22-25]進行比對校驗。
1.3 沙棘根瘤內生菌的高通量測序分析
1.3.1 建庫測序 提取沙棘根瘤總 DNA 后,根據 16S rDNA 保守區設計引物 (引物序列 335F:5′-CADACTCCTACGGGAGGC-3′,769R:5′-ATCCTGTTTGMTMCCCVCRC-3′),在引物末端加上測序接頭,便于建庫時添加能區分樣本的堿基序列的條碼/索引(barcode/index)。再進行PCR擴增并對其產物進行紫外分光光度計定量及混樣、過柱純化和均一化形成測序文庫,建好的文庫先進行文庫質檢,質檢合格的文庫用Illumina HiSeq 2500進行測序[26]。高通量測序得到的原始圖像數據文件,經堿基識別分析轉化為原始測序序列,結果以FASTQ (簡稱為fq)文件格式存儲[27]。
1.3.2 測序數據處理 首先使用 Trimmomatic v.0.33 軟件[28],對測序得到的原始測序序列進行過濾;其次使用cutadapt 1.9.1軟件進行引物序列的識別與去除,得到不包含引物序列的高質量測序序列;然后使用FLASH v1.2.7軟件[29],按照最小重疊(overlap)長度為10 bp、重疊區允許的最大錯配比率為0.2的要求,對每個樣品高質量的一小段短的基因測序片段(reads)進行拼接,得到的拼接序列即原始序列質控后的高質量測序序列(clean reads);最后使用UCHIME v4.2軟件[30],鑒定并去除嵌合體序列,得到最終有效數據。使用Usearch軟件對reads在97.0%的相似度水平下進行聚類,獲得分類操作單元(OTU)[31],以測序所有序列數的0.005%作為閾值過濾OTU[32]。以SILVA (http://www.arb-silva.de/)為參考數據庫使用樸素貝葉斯分類器對特征序列進行分類學注釋,可得到每個特征對應的物種分類信息,進而在各水平(門、綱、目、科、屬、種)統計樣品群落組成,利用QIIME軟件生成不同分類水平上的物種豐度表,再利用R語言工具繪制樣品分類學水平下的群落結構圖[33]。使用QIIME軟件對樣品α多樣性進行評估和t檢驗(顯著性水平為0.01)。利用Mothur v1.30軟件和R語言工具包繪制稀釋曲線。基于獨立OTU,采用加權分析方法和Bray-Curtis算法,使用QIIME軟件進行非加權組平均法(UPGMA)分析,比較各組樣品間的物種差異。
2 結果與分析
2.1 分類操作單元 (OTU)聚類分析
使用Usearch軟件對clean reads在97.0%的相似度水平下進行聚類,共計獲得651個OTU。各樣品OTU個數分布較為均勻,樣品M1~M6分別為551、583、579、518、593、589個。如圖1所示:6組樣品中共有的OTU數為417個。M3、M5、M6中分別有4、2、9個特有的OTU,為樣品特有OTU,非單個樣品特有或所有樣品間共有的OTU在圖1未做展示。從整體來看,不同地點的各樣品間的OTU差異性遠小于共性,說明采樣方法設計合理。

圖1 沙棘(M1~M6)根瘤樣品分類操作單元(OTU)花瓣圖Figure 1 Petal image of operational taxonomic unit (OTU) of H.rhamnoides root nodule sample (M1-M6)
2.2 測序深度分析與 α 多樣性指數分析
對6組樣品測序共獲得 810 039對reads,雙端reads質控、拼接后共產生 617 188 條 clean reads。其中質量≥20的堿基占總堿基數的比例(Q20)為98.7%,質量≥30的堿基占總堿基數的比例(Q30)為95.4%,表明測序質量較好。從圖2可見:各樣品稀釋性曲線趨向平緩,表明在持續抽樣下新物種出現的速率逐漸趨于平緩,此環境中物種數量不會隨測序數量的增加而顯著增多[34],說明取樣合理,能較好體現6組樣品中根瘤內生菌的多樣性,可以進行數據分析。M5的Shannon和Simpson指數最大(表1),說明物種多樣性最高。同理,M4的物種多樣性最低。物種豐度方面M5與M6差別不大,均有較高水平。M4根瘤樣品的物種豐度最低。樣點的Shannon指數平均為4.24,Simpson指數平均為0.70,Ace指數平均為585.79,Chao1指數平均為595.47,樣本文庫平均覆蓋率為99.95%。說明采樣地的沙棘根瘤內生菌的物種豐富且多樣性較大,各物種分配相對均勻,其微生物物種信息得到了充分體現。
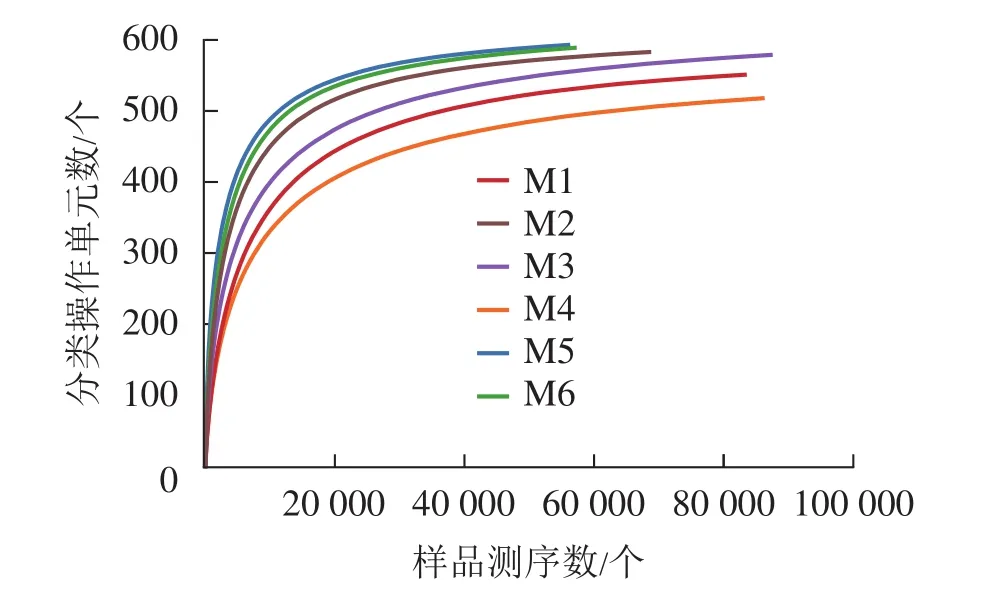
圖2 各樣品稀釋性曲線Figure 2 Dilution curve of each sample

表1 各組樣品的 α 多樣性指數Table 1 Alpha diversity index for each group of samples
2.3 傳統純培養與高通量測序分析沙棘根瘤內生菌的群落結構及其差異
通過傳統分離方法從BAP、JA、S、高氏一號培養基中得到純培養菌株96株。所有菌株均可傳代培養,但菌株之間培養周期差異較大,培養周期在1~30 d呈離散型分布。對各菌株進行分子鑒定,共有4門8綱8目13科19屬。在門的分類水平分別為變形菌門Proteobacteria、放線菌門Actinobacteria、厚壁菌門Firmicutes和柔膜菌門Tenericutes。在屬的分類水平上,96株菌分屬于支原體屬Mycoptasma1株、慢生根瘤菌屬Bradyrhizobim6株、土壤桿菌屬Agrobacterium7株、腸桿菌屬Enterobacter6株、小坂菌屬Kosakonia8株、檸檬酸桿菌屬Citrobacter1株、約克氏菌屬Yokenella1株、歐文氏菌屬Erwinia1株、克羅諾桿菌屬Cronobacter2株、泛菌屬Pantoea1株、莫拉菌屬Moraxella1株、貪噬菌屬Variovorax1株、草螺菌屬Herbaspirillum1株、假單胞菌屬Pseudomonas5株、鏈霉菌屬Streptomyces14株、小單孢菌屬Micromonospora1株、短桿菌屬Brevibacterium6株、葡萄球菌屬Straphylococcus1株和芽孢桿菌屬Bacillus32株。其中,優勢門為變形菌門和厚壁菌門,優勢屬為芽孢桿菌屬和鏈霉菌屬。
高通量測序分析發現:6組樣品共有14門34綱89目148科314屬。將相對豐度大于0.1%的門與相對豐度前10的屬進行匯總(圖3、表2、表3)發現:在門的分類水平上,6組樣品中相對豐度較高的主要為放線菌門和變形菌門,兩者相對豐度之和為87.5%~97.1%。其次為擬桿菌門Bacteroidetes、桿菌門Patescibacteria、厚壁菌門、酸桿菌門Acidobacteria。在屬的分類水平上,弗蘭克氏菌屬Frankia占絕對優勢,相對豐度為20.12%~74.81%,平均相對豐度為51.49%。其次為根瘤菌屬Rhizobium、類固醇桿菌屬Steroidobacter、糖單孢菌屬Saccharimonadales、腸桿菌屬、泛菌屬、歐文氏菌屬、假黃色單胞菌屬Pseudoxanthomonas、鞘脂單胞菌屬Sphingomonas、假單胞菌屬、固氮弓菌屬Azoarcus、伯克氏菌屬Burkholderia、芽單胞菌屬Blastomonas、聚集桿菌屬Congregibacter、拉恩氏菌屬Rahnella、鞘氨醇菌屬Chitinophaga、獨島桿菌屬Dokdonella、普雷沃氏菌屬Prevotella、鏈霉菌屬、Microtrichales屬。
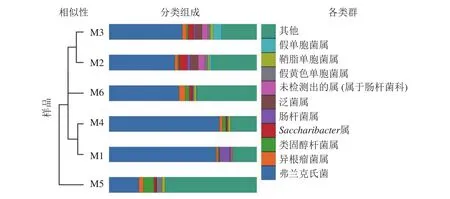
圖3 6組根瘤樣品的非加權組平均法(UPGMA)聚類樹與物種分布柱狀圖Figure 3 UPGMA clustering tree and the species distribution histogram of the six groups of nodule samples are combined drawing

表2 沙棘微生物區系門水平的相對分布Table 2 Relative abundance of microbiota taxa at the level of phylum

表3 沙棘微生物區系屬水平的相對分布Table 3 Relative abundance of microbiota taxa at the level of genus
在門、綱、目、科、屬的各分類單元中,高通量測序的檢測靈敏度(高通量測序/純培養)依次是純培養方法的3.50、4.25、11.20、11.38和16.53倍。在門水平上,純培養菌株中占比較高的厚壁菌門在高通量測序中占比并不高。在屬水平上,純培養菌株中占比較高的芽孢桿菌屬和鏈霉菌屬皆在高通量測序中占比很低。該對比結果差異性較大,說明高通量測序在微生物多樣性分析中占據優勢地位,要優于純培養方法。同時也說明,沙棘根瘤內共生細菌群落結構更為復雜,群落更為穩定。
3 結論與討論
3.1 結論
在運用傳統方法分離純培養微生物時,共分離純培養菌株96株,分屬于4門8綱8目13科19屬,未獲得弗蘭克氏菌屬的菌株,可能是培養基中弗蘭克氏菌屬的菌株生長緩慢,易被其他菌群取代,因此仍需探索新的培養基與培養方法以遏制根瘤中其他菌株的繁殖。在微生物多樣性分析中,由于環境中的微生物復雜多樣,各環境之間組成差異較大,通常采用非加權方法進行分析。該方法簡單易操作,主要考慮物種的有無,但未考慮物種的豐度,所以采用非加權的方法難以區別各樣品間的差異。
高通量測序分析共檢測到14門34綱89目148科314屬。在門、綱、目、科、屬的各分類單元中,高通量測序的檢測靈敏度(高通量測序/純培養)依次是純培養方法的3.50、4.25、11.20、11.38和16.53倍。與純培養獲得的菌株相比,高通量測序分析結果更加完整地揭示了沙棘根瘤內生菌的微生物多樣性。高通量測序表明:在門的分類水平上,樣品中相對豐度較高的主要為放線菌門和變形菌門,兩者相對豐度之和為87.5%~97.1%。在屬的分類水平上,弗蘭克氏菌屬占絕對優勢,相對豐度為20.12%~74.81%,平均相對豐度為51.49%。
3.2 討論
張愛梅等[35]和劉志強等[36]分別對甘肅榆中、遼寧通遼、內蒙古赤峰等地沙棘根瘤內生菌微生物多樣性做過類似研究,其高通量測序所得的微生物多樣性高于本研究結果,說明沙棘根瘤內生菌微生物多樣性受地理位置、土壤成分、氣候條件、宿主種類及生長環境等多種因素的影響。本研究的沙棘取樣于內蒙古烏蘭察布沙漠邊緣地帶,采樣地荒漠化土壤與干旱少雨氣候對內生菌多樣性有特別影響。
屬于非豆科Leguminosae植物的沙棘根瘤共生固氮體系是以弗蘭克氏菌屬為主導的[37]微生物—微生物—植物互作體系。高通量測序分析顯示:弗蘭克氏菌屬所占比例較高,然而本次傳統方法分離卻未得到純培養菌株,這可能是由于培養基中缺乏某種信號物質或與其他菌屬競爭存在劣勢導致的,建議添加制霉菌素、萘啶酮酸和放線菌酮抑制其他菌群的繁殖[18]。非豆科植物結瘤固氮過程,單一屬的菌株難以完成此任務。有研究[38]表明:純培養分離的貪噬菌屬是復雜微生物組中維持根生長的核心菌屬,并且具有產生和降解生長素的能力,是細菌—細菌—植物通訊網絡的關鍵角色。小單孢菌是植物益生菌,在促進植物生長的同時還可以分泌細胞壁降解酶促進細胞壁的降解,進而便于弗蘭克氏菌的侵染[39-40],但是小單孢菌的快速繁殖也對弗蘭克氏菌的生長起到抑制作用。沙棘作為胡頹子科植物,根部結瘤侵染方式為細胞間侵入。研究[41]表明:草螺旋菌屬Spirillum、慢生根瘤菌屬、腸桿菌屬的相關細菌與弗蘭克氏菌存在負相關性(即抑制關系),以上3個菌屬均在豆科、禾本科Poaceae植物中發揮固氮相關的重要作用,但在胡頹子科中此類細菌與弗蘭克氏菌屬相互作用的機制尚未明確。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
