林分密度和種植點配置對楊樹各器官非結構性碳水化合物的影響
2022-04-08 12:21:08曹鵬鶴孫杰杰王玉超韓夢華郭佳歡馮會麗王維楓
浙江農林大學學報 2022年2期
關鍵詞:質量
曹鵬鶴,徐 璇,孫杰杰,王玉超,韓夢華,郭佳歡,馮會麗,王維楓
(南京林業大學 生物與環境學院,江蘇 南京 210037)
植物通過光合作用固定大氣中的碳并產生碳水化合物,不同的碳水化合物在植物生長發育過程中的作用不同[1]。結構性碳水化合物(SC)主要用于植物的結構組織構建,非結構性碳水化合物(NSC)參與植物的生理代謝和滲透調節等過程,是植物進行生理調節的物質基礎[2]。葡萄糖、果糖和蔗糖等可溶性糖及淀粉,是NSC的主要組成部分[3]。植物利用可溶性糖調節自身生理狀態,同時將部分碳水化合物合成淀粉作為儲能物質。當植物面臨碳資源不足時,可將淀粉再活化形成可溶性糖以及時補充碳資源[4]。植物體內的NSC水平受生物和環境因素影響較大,土壤水分、氣溫、光照、損傷、常綠樹種的葉齡以及植物自身休眠與否等[5-6]都在不同程度上調控著植物體內的NSC水平[7-8]。除此之外,所有影響植物生長的因素也都會改變NSC在不同器官間的分配格局。NSC及其組分的含量水平是植物在不同生長階段中碳收支平衡的體現,反映了植物的生長狀態和碳代謝差異[9],因此對樹木中NSC進行探究是研究樹木適應性和固碳潛力的關鍵。
林分密度和種植點配置是人工林造林的重要技術措施,合理的林分密度和空間配置有助于植株的良好生長,可以優化樹木生長的地上空間結構。林分密度與種植點配置共同調控了地上部分和地下根系的協調發展和碳分配格局。不同的造林方式通過改變林分采光時間、光照角度和林分內微環境的氣象條件等影響地上冠層的光合作用,同時也作用于林分地下根系生長及其對水肥的吸收效率。研究表明:林分密度影響造林樹種的生長狀況(樹高、胸徑等)[10]和林下土壤理化性質[11]等。不同林分密度和栽種配置可構建不同的生長空間,林木在不同的生長空間中調節碳資源在各組織間的分配,加強自身的競爭能力和對環境的適應性。有研究指出:紅樺Betula albosinensis幼苗的光合適應性受林分密度的調控,隨林分密度的增加,苗木的二氧化碳同化率升高[12]。林分密度對不同樹種碳分配的影響各異,高密度林分中紫果云杉Picea pururea針葉中淀粉和可溶性糖含量均表現為較高水平[13],并且林分密度增大也能夠促進雜交楊Populus canadensis×Populus maximowiczii向地下部分投資碳資源[14]。然而也有研究表明:蘋果Malus×domestica中低密度配置時,具有更高的可溶性糖和NSC含量[15]。當林分密度為中低水平時,林分環境中土壤微生物多樣性較高,碳代謝能力更強[16]。林分密度對不同樹種NSC含量的影響存在差異,密度影響樹木各器官NSC分布仍需進一步探討。種植點配置對樹木的生物特性也有一定的影響,相比于長方形種植點配置,正方形配置中的林分板材質量更優[17],并且林分根系生長空間更加均一,根系對養分的吸收能力更高[18]。然而關于種植點配置對樹木內部各器官NSC分布影響的相關研究較少。
NSC代謝是生態系統中植物碳循環過程重要的組成部分[19],對NSC的合理分配和調節是植物生長發育的基礎。楊樹Populus作為一種快速生長型的落葉喬木,具有明顯的生長季和休眠季之分,樹木在生長季對養分的快速消耗使NSC在各器官之間的分配特征更加明顯。因此,本研究以蘇北地區楊樹人工林為研究對象,對不同林分密度和種植點配置下樹木中NSC的分配和利用進行了研究,以期為營造高效固碳人工林提供科學參考。
1 研究地區與研究方法
1.1 研究區概況
本研究地位于江蘇省宿遷市泗洪林場馬浪湖分場(33°33′N,118°32′E),地處江蘇中北部洪澤湖西岸,屬北亞熱帶和暖溫帶季風氣候交界區,年平均氣溫為14.4 ℃,無霜期為197 d,年平均降水量為973 mm,降水主要集中在6—8月,土壤母質為洪澤湖淤積土,土壤質地多為中壤至輕黏。
研究地中栽植的楊樹品種為‘南林95’楊Populus×euramericana‘Nanlin 95’,楊屬Populus黑楊派,具有品質優良、環境適應性廣、速生等特點,是長江中下游區域人工造林的主栽品種之一。于2007年以1年生帶根幼苗進行試驗林造林[20]。該試驗林以高(400 株·hm-2)、低(278 株·hm-2) 2個種植密度造林,在高、低林分密度下均設置有正方形和長方形2種種植點配置(表1)。不同林分密度和種植點配置下各設置3個30 m × 60 m的試驗小區(3個重復),并使用胸徑尺和激光測距測高儀(Nikon F550,日本)調查了整個林分中共計632棵楊樹的胸徑(cm)和樹高(m)用于估算生物量。

表1 試驗林分及其土壤 0~20 cm 土層概況Table 1 Basic status of experimental stands and basic characters of 0-20 cm soil layer
1.2 樣品采集
于 2018 年 7 月中旬 (楊樹生長季),分別在高密度正方形配置 (400 株·hm-2,5.0 m × 5.0 m)、高密度長方形配置 (400 株·hm-2,3.0 m × 8.0 m)、低密度正方形配置 (278 株·hm-2、6.0 m × 6.0 m)和低密度長方形配置(278株·hm-2,4.5 m × 8.0 m)的樣地中選取2株與標準木相似的、健康、直立、生長狀態良好和無病蟲害的樣株進行取樣。每樣株取健康葉片(大于30片)和1年生嫩枝(3~5枝,干質量大于10 g),葉和枝樣品均采集于冠層中上部向陽位置[21],采樣時間為8:00—11:00。使用生長錐在胸徑處于南北向、東西向各鉆取樹干木芯樣品2支,在距離根莖50 cm位置處人工挖掘2~10 mm的粗根樣品(由于細根采集樣本過少,不作為研究對象),將采集好的樣品立刻裝入配有冰板的保溫箱中低溫保存并帶回實驗室備用(<24 h)。在實驗室將樣品清洗干凈后,放入烘箱中65 ℃烘干至恒量,然后用組織研磨儀磨粉,室溫避光處保存待測。
1.3 測定指標
采用改進的苯酚硫酸法[22]提取樣品中的淀粉和可溶性糖,根據可溶性糖和淀粉的不同紫外吸光值,計算單位組織樣品中的可溶性糖和淀粉的質量分數(可溶性糖和淀粉占NSC的絕大部分,本研究以可溶性糖和淀粉質量分數之和為NSC總量[23])。
1.4 NSC 庫估算
根據生物量方程,來估算楊樹各器官的生物量[24]。各器官中NSC儲量由NSC含量和生物量計算得出,單株NSC儲量通過各個器官中NSC儲量加權得出,林分NSC儲量由林分密度和單株NSC儲量計算得到。

式(1) ~ (4)中:W葉為葉生物量;W枝為枝生物量;W干為樹干生物量;W根為根生物量;D為胸徑(cm);H為樹高(m)。
1.5 數據處理
采用嵌套方差分析法,分析不同林分密度和種植點配置間楊樹人工林總NSC儲量以及個體單株葉、枝、干和根中NSC及組分含量的差異性水平,顯著水平為0.05。使用單因素方差分析(ANOVA)比較楊樹不同器官間NSC及其組分含量的差異性,并使用最小顯著差異法(LSD)進行多重對比(P<0.05)。利用Pearson相關分析法,估算楊樹各器官NSC及組分含量間的相關關系。以上分析均使用SPSS 23.0 完成,用 Origin 2017 作圖。
2 結果與分析
2.1 楊樹各器官的 NSC 質量分數
從圖1可見:楊樹各器官的NSC及其組分質量分數存在顯著差異(P<0.05),從葉、枝、到干的可溶性糖質量分數依次遞減,到根部有所升高。楊樹各器官中的淀粉質量分數從樹干位置(最高)開始,向地上部分(枝、葉)和地下部分遞減,而NSC總量在各個器官中從大到小依次為根、枝、葉、干。楊樹葉、枝、干、根的可溶性糖質量分數占NSC總量的比例分別為62.3%、33.2%、15.9%和36.8%,淀粉質量分數占NSC總量的比例分別為37.7%、66.8%、84.1%和63.2%。

圖1 楊樹不同器官的非結構性碳水化合物(NSC)質量分數Figure 1 Non-structural carbohydrates (NSC) concentrations in the different organs of poplar
從圖2可見:養分利用的就近原則使NSC及組分質量分數的顯著相關關系主要發生在相鄰器官之間。枝中可溶性糖分別與葉的NSC和淀粉呈顯著負相關關系(P<0.05);樹干可溶性糖與根的NSC及各組分均為顯著正相關(P<0.05)。器官內部(葉、根)對碳資源的利用與儲存則主要表現為顯著正相關(P<0.05),體現了NSC及組分在不同器官間以及器官內調節與分配關系的多樣性。

圖2 楊樹各器官非結構性碳水化合物及各組分的相關關系Figure 2 Correlation analysis of non-structural carbohydrate content and its components among poplar organs
2.2 不同林分密度和種植點配置下楊樹各器官可溶性糖質量分數
從圖3可見:林分密度對不同器官中可溶性糖質量分數的影響存在差異。不同林分密度下楊樹葉中可溶性糖質量分數差異極顯著(P<0.01),枝、干和根中可溶性糖質量分數則不受林分密度影響。種植點配置對可溶性糖質量分數的影響因器官而異,不同配置下楊樹葉、根中的可溶性糖質量分數差異極顯著(P<0.01),樹干中可溶性糖質量分數差異顯著(P<0.05),枝中可溶性糖質量分數在不同配置間差異不顯著。
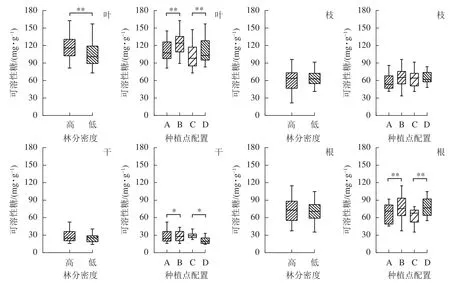
圖3 不同林分密度和種植點配置對楊樹各器官中可溶性糖的影響Figure 3 Effect of stand densities and spacing configurations on soluble sugar in different organs of poplar
根與葉中可溶性糖分布相一致,均為同密度下正方形配置高于長方形配置。不同配置林分楊樹樹干中可溶性糖表現趨勢不同,高密度林分表現為正方形配置大于長方形配置,而低密度林分則表現為正方形配置小于同密度下的長方形配置。
2.3 不同林分密度和種植點配置下楊樹各器官淀粉質量分數
從圖4可見:楊樹地上部分各器官中淀粉質量分數均不受林分密度和種植點配置的影響。林分密度對地下部分(根)的淀粉影響也不顯著,然而不同種植點配置對楊樹根中淀粉質量分數的影響差異極顯著(P<0.01)。高密度下,正方形配置的楊樹根中淀粉質量分數極顯著高于長方形配置(P<0.01);與之相似,低密度林分條件下正方形配置的楊樹根中淀粉同樣極顯著高于長方形配置(P<0.01)。

圖4 不同林分密度和種植點配置對楊樹各器官中淀粉的影響Figure 4 Effect of stand densities and spacing configurations on starch in different organs of poplar
2.4 不同林分密度和種植點配置下楊樹各器官NSC質量分數
從圖5可見:楊樹葉中NSC質量分數在不同林分密度間差異顯著(P<0.05),其他器官中NSC質量分數受林分密度影響差異不顯著,高密度林分的楊樹葉中NSC質量分數顯著高于低密度林分。葉中NSC質量分數受種植點配置的影響顯著(P<0.05),根中NSC質量分數受種植點配置的影響極顯著(P<0.01),其他器官中NSC質量分數在不同林分密度和種植點配置下差異不顯著。整體上,相同林分密度的楊樹葉和根中NSC質量分數均表現為對應的正方形配置大于長方形配置。

圖5 不同林分密度和種植點配置對楊樹各器官非結構性碳水化合物 (NSC)的影響Figure 5 Effect of stand densities and spacing configurations on non-structural carbohydrate (NSC) in different organs of poplar
2.5 不同林分密度和種植點配置下林分總NSC儲量特征
根據生物量估測公式(1) ~ (4),得到單株生物量(表2)。由表2可知:楊樹單株生物量在不同林分密度間差異極顯著(P<0.01),低密度楊樹單株生物量顯著大于高密度單株生物量;楊樹單株生物量在不同種植點配置間差異極顯著(P<0.01)。在林分密度相同的情況下,楊樹單株生物量均表現為正方形配置大于長方形配置。

表2 不同林分密度及種植點配置中單株生物量方差分析Table 2 Variance analysis of individual tree biomass in stands with different stand densities and spacing configurations
從圖6可見:林分總生物量和林分NSC總儲量受種植點配置的影響極顯著(P<0.01)。當林分密度為400和278 株·hm-2時,林分總生物量和總NSC儲量受林分密度的影響不顯著。相同林分密度下林分NSC總儲量均表現為正方形配置高于長方形配置。

圖6 不同密度和種植點配置對林分生物量和非結構性碳水化合物(NSC)儲量的影響Figure 6 Effect of different densities and spacing configurations on biomass and non-structural carbohydrate (NSC) reserves in the stand level
3 討論
3.1 生長季楊樹各器官 NSC 分配特征
碳資源是植物生長發育的代謝基礎,植物按照一定比例將碳分配至各個器官中[3]。楊樹的葉、枝、干、根4個器官中,可溶性糖從大到小依次為葉、根、枝、干,淀粉從大到小依次為干、根、枝、葉。糖類在器官間分配格局主要受“源—匯”關系和同化物利用的就近原則影響[9],在葉和根系等生長性強的器官中居多[25],其次是離生長中心較近的器官較多,這與劉萬德等[26]的結果相似。而淀粉作為暫時儲存物質,當光合產物的消耗大于合成時,淀粉被重新活化為可溶性糖,用于緩沖碳資源不足[27]。因此淀粉主要分布在干、根和枝等儲存器官中,葉中僅有少量淀粉存在。然而有研究表明:興安落葉松Larix gmelinii和紅松Pinus koraiensis的根具有較高的可溶性糖[28],這與本研究結果不一致。但是該地區蒙古櫟Quercus mongolica可溶性糖含量在器官間的分布與本研究結果相似。此外,蒙古櫟和興安落葉松根中的淀粉含量處于相對較高的水平值[28],這與本研究中樹干淀粉最高,根部淀粉次之的結果不一致,這可能由不同針闊樹種及氣候差異所引起。上述2個樹種所處樣地年均氣溫和降水量分別為3.1 ℃和629 mm,而本研究中樣地年均氣溫和降水量分別為14.4 ℃ 和973 mm。長期的低溫馴化驅動蒙古櫟和興安落葉松將淀粉更多地轉移并儲存于地下根系,當低溫來臨時轉化為可溶性糖增加根系細胞中的滲透物質,以避免根系因“根際低溫”而出現生長被抑制的現象[29]。相比之下,楊樹處于相對溫濕的環境中則不需要在根系中儲存大量的淀粉來應對低溫脅迫,因此淀粉被更多的儲存在樹干中。盡管根系和樹干均為主要的儲存器官[30],但還沒有明確的研究證明根系和樹干儲存的主次關系,儲存器官的優先級可能受到多種因素影響。本研究中蘇北地區楊樹在生長季將碳更多地分配到地上部分,把樹干作為主要的儲存器官,這也可能是楊樹生長于該地區所形成的一種適應性策略。
3.2 林分密度和種植點配置對楊樹個體與林分水平NSC影響
人工造林時,較高的碳資源儲量是苗木健康生長的重要保障[31]。碳水化合物的含量水平反映了林木的生長狀態。整體上,本研究中個體水平上僅葉器官中可溶性糖和總NSC質量分數在不同林分密度間存在顯著差異(P<0.05),且楊樹葉中NSC及可溶性糖均表現為高密度(400 株·hm-2)大于低密度(278 株·hm-2)。這可能是高密度林分提高了葉片光合作用,進而促使葉中具有較多的光合產物。研究表明:5年生楊樹種苗在較高的林分密度能有效地增加冠層的光攔截量,提高冠層的光合效率[32];并且在較低的林分密度中光能可能會被過多的耗散,降低了冠層對光能的利用效率[33]。林分水平的NSC儲量在不同林分密度間差異不顯著,這與TRUAX等[14]的研究結果一致,可能是研究區種植密度梯度差較小的緣故。
總的來說,在相同林分密度下不同種植點配置間楊樹個體和林分水平上的NSC含量及總儲量均表現為正方形配置大于長方形配置。與以往研究表明“寬行窄距”林分配置中個體具有更強生長潛力[34]的結論不一致,可能是由于當單株種植面積相似時,“寬行窄距”的配置使個體生長空間有所壓縮,個體在正方形配置的林分中具有更加均勻的生長空間,對水分和光能的利用率有所提高[35]。王琪等[18]研究表明:楊樹細根對正方形配置的土地空間利用度更高。然而種植點配置對個體的影響具有較大的變異性,當評估種植點配置對個體的效應時,應結合造林密度、造林樹種、林齡以及造林地環境條件等因素綜合考慮種植點配置對個體的影響。
綜上可知,較高的林分密度促進了楊樹葉中可溶性糖質量分數的升高。相同林分密度下不同配置間,正方形配置對楊樹葉、干和根中NSC質量分數具有促進作用,正方形種植點配置更有利于楊樹儲備碳資源,促進林分中個體的生長發育。未來進行營林活動時,相同林分密度下應優先考慮正方形的種植點配置,以促進個體積累更多的碳資源,提升個體及林分水平的固碳增匯潛力。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
