甘肅隴東大豆孢囊線蟲田間侵染動(dòng)態(tài)
2022-04-04 14:16:21羅寧李惠霞劉永剛李煥宇楊順義李金章石明明李金鴻
植物保護(hù) 2022年2期
關(guān)鍵詞:大豆
羅寧 李惠霞 劉永剛 李煥宇 楊順義 李金章 石明明 李金鴻


摘要 為探討甘肅隴東地區(qū)大豆孢囊線蟲田間侵染動(dòng)態(tài),于2017年-2019年對(duì)大豆根系和根際土壤定期定點(diǎn)取樣,采用漂浮法、貝曼漏斗法及酸性品紅染色法對(duì)土樣中和根組織內(nèi)線蟲進(jìn)行分離計(jì)數(shù),研究其繁殖數(shù)量與氣溫、降雨量的關(guān)系。結(jié)果表明,2017年土壤中2齡幼蟲和新鮮孢囊的數(shù)量有3個(gè)明顯高峰,大豆根組織內(nèi)2~4齡幼蟲均出現(xiàn)3個(gè)明顯高峰,即發(fā)生3代,平均世代歷期35 d。2018年-2019年土壤中2齡幼蟲和新鮮孢囊均有4個(gè)明顯高峰,大豆根組織內(nèi)2~4齡幼蟲亦出現(xiàn)4個(gè)明顯高峰,即發(fā)生4代。第1代侵染數(shù)量最多,是主要危害世代。最后1代侵染數(shù)量極少且發(fā)育緩慢。在2017年-2019年生長(zhǎng)季中,線蟲繁殖數(shù)量與生長(zhǎng)季平均溫度具有相關(guān)性,與累積降雨量無相關(guān)性。
關(guān)鍵詞 大豆; 大豆孢囊線蟲; 侵染特征; 世代循環(huán)
中圖分類號(hào): S435.651
文獻(xiàn)標(biāo)識(shí)碼: A
DOI: 10.16688/j.zwbh.2021019
Abstract In order to explore the infection dynamics of soybean cyst nematode in the soybean field in east of Gansu province, soil and root samples were periodically collected from one fixed soybean block from 2017 to 2019. The nematodes were separated and counted under microscope by floating, Baermann funnel and acid fuchsin staining method, and the relationship between the reproduction and temperature and humidity were investigated. The results showed that there were three distinct number peaks of 2nd stage juvenile and new cysts in the soil, and three peaks of the emergence of 2nd, 3rd and 4th juveniles in soybean roots, which occurred in three generations and the average period of one generation was 35 d in 2017. From 2018 to 2019, there were four distinct number peaks of 2nd stage juvenile and new cysts in the soil and four peaks of the emergence of 2nd, 3rd and 4th juveniles in soybean roots, indicating that four generations occurred. The first generation had the largest number of infections, which was the main damage period and nematodes developed slowly in last generation. During the growing season from 2017 to 2019, the number of nematodes reproduction was correlated with the average temperature, but not with the accumulated rainfall.
Key words soybean; Heterodera glycines; infection characteristics; generation cycle
大豆孢囊線蟲(soybean cyst nematode,SCN)屬側(cè)尾腺口綱墊刃目異皮科孢囊線蟲屬,是大豆生產(chǎn)中危害最嚴(yán)重的植物病原線蟲[1]。目前,在美國(guó)、加拿大、波多黎各、海地、阿根廷、巴西、巴拉圭、哥倫比亞、俄羅斯、比利時(shí)、英國(guó)、意大利、日本、韓國(guó)、朝鮮、伊朗、印度尼西亞和埃及等國(guó)家大豆種植區(qū)普遍發(fā)生[25]。我國(guó)23個(gè)省(自治區(qū),直轄市)已有大豆孢囊線蟲分布[611],其中在黃淮海和東北大豆產(chǎn)區(qū)危害尤為嚴(yán)重,每年危害面積超過267萬 hm2[12],隨著大豆連作年限增加,病情逐年加重,嚴(yán)重者減產(chǎn)80%以上,甚至絕收。
據(jù)報(bào)道,大豆孢囊線蟲理論上1年可以繁殖6代,具體世代數(shù)主要受土壤溫度、寄主品種、播種時(shí)間和播種期等因素影響[13]。美國(guó)明尼蘇達(dá)州大豆孢囊線蟲1年發(fā)生3~4代[14]。俄羅斯阿穆爾州地區(qū)1年發(fā)生3代,平均歷期27.3 d[15]。在我國(guó),黑龍江省1年發(fā)生3代,第1代發(fā)生量最大[16],在山西、江蘇和河南等省1年發(fā)生4代[6]。同一地區(qū)不同生長(zhǎng)季,大豆孢囊線蟲發(fā)生代數(shù)存在差異。研究顯示,大豆孢囊線蟲2013年在山東大豆上發(fā)生3代,2014年發(fā)生4代,第2代有明顯世代重疊現(xiàn)象[1314]。北京地區(qū),大豆孢囊線蟲在地黃上1年發(fā)生5~6代[17]。目前,甘肅省大豆孢囊線蟲的侵染動(dòng)態(tài)尚無研究。
甘肅省是西北地區(qū)大豆主產(chǎn)區(qū)之一,年種植面積維持在13.33萬~15.33萬hm2,其中隴東地區(qū)種植面積最大,達(dá)10萬hm2[18],本課題組調(diào)查發(fā)現(xiàn),隴東地區(qū)大豆孢囊線蟲發(fā)生嚴(yán)重,孢囊檢出率為87.2%,已成為該地區(qū)大豆生產(chǎn)的主要限制因素[19]。
在大豆生長(zhǎng)季,孢囊線蟲的侵染次數(shù)與數(shù)量受多種因素影響,如寄主品種、氣溫和降雨量等均不同程度地影響大豆孢囊線蟲群體繁殖[2022],另外耕作方法和土壤質(zhì)地也會(huì)導(dǎo)致大豆孢囊線蟲群體變化[2325]。我國(guó)幅員遼闊,大豆種植區(qū)跨度大,甘肅與黑龍江、江蘇及山東等大豆產(chǎn)區(qū)氣候條件、大豆播種期均不同,大豆孢囊線蟲侵染規(guī)律與其他地區(qū)是否存在差異尚不明確。因此,本研究在2017年-2019年春播大豆生育期內(nèi)對(duì)大豆孢囊線蟲群體變化動(dòng)態(tài)進(jìn)行系統(tǒng)研究,探究氣溫、降雨對(duì)其繁殖的影響,以期明確該地區(qū)大豆孢囊線蟲田間侵染規(guī)律,為大豆孢囊線蟲有效防治提供依據(jù)。
1 材料與方法
1.1 供試品種
大豆‘中黃30號(hào)’(市售),中感大豆孢囊線蟲病。
1.2 試驗(yàn)地概況
試驗(yàn)地位于甘肅省慶陽市寧縣良平鎮(zhèn)豐樂村(35°26′E,108°04′N),年平均氣溫8.7℃,無霜期168 d,年均降水量565.9 mm,降雨多集中在7月-9月,季節(jié)分配不均,屬于典型黃土高原半濕潤(rùn)偏干旱氣候類型區(qū)[26]。試驗(yàn)地連續(xù)多年種植大豆,孢囊線蟲基數(shù)較高,平均37.4個(gè)/100 g。分別于2017年5月1日、2018年5月2日和2019年5月5日播種大豆,播種后田間無人工灌溉,進(jìn)行正常鋤草和噴藥管理。
1.3 取樣時(shí)間和方法
2017年5月15日至9月23日內(nèi)共取樣14次,每間隔10 d左右取樣1次。2018年5月13日至9月21日內(nèi)共取樣19次,2019年5月15日至9月20日內(nèi)共取樣19次,每間隔6~8 d取樣1次。采用“ZigZag”取樣法,每次隨機(jī)選取10個(gè)樣點(diǎn),每個(gè)樣點(diǎn)挖采大豆根系2~3株,保持根系完整,裝入取樣袋。每個(gè)樣點(diǎn)挖取根圍深度0~20 cm土壤作為1個(gè)小樣,將10個(gè)小樣裝入塑料袋混合作為1個(gè)樣品,約1 kg,帶回實(shí)驗(yàn)室檢測(cè)。
1.4 線蟲的室內(nèi)檢測(cè)
1.4.1 土壤中線蟲的分離
將土樣混合均勻后稱取100 g,采用漂浮法和貝曼漏斗法對(duì)孢囊和2齡幼蟲(J2)進(jìn)行定量分離[27],在60目篩上收集白雌蟲和孢囊,解剖鏡和顯微鏡下對(duì)孢囊和J2進(jìn)行鏡檢和計(jì)數(shù)。
1.4.2 根組織中線蟲的染色和計(jì)數(shù)
將混合樣中的大豆根系清洗干凈剪成2~3 cm小段并稱重,采用酸性品紅對(duì)根組織進(jìn)行染色[4]。在顯微鏡下對(duì)染色的2齡、3齡(J3)、4齡(J4)幼蟲和根組織內(nèi)年輕雌蟲進(jìn)行鏡檢、計(jì)數(shù)和拍照。
1.4.3 孢囊線蟲世代的劃分標(biāo)準(zhǔn)
依據(jù)土壤中J2、新鮮孢囊出現(xiàn)的高峰數(shù)及其時(shí)間間隔長(zhǎng)短,同時(shí)參考根組織中J2、J3、J4和年輕雌蟲出現(xiàn)情況來確定大豆孢囊線蟲的世代。
1.5 數(shù)據(jù)統(tǒng)計(jì)與分析
統(tǒng)計(jì)每100 g土壤中孢囊(白雌蟲和新鮮孢囊)和J2的數(shù)量,以及1 g根組織內(nèi)J2、J3、J4和年輕雌蟲的數(shù)量,統(tǒng)計(jì)各蟲態(tài)發(fā)育動(dòng)態(tài)。從當(dāng)?shù)貧庀缶肢@取當(dāng)年氣象數(shù)據(jù),統(tǒng)計(jì)每個(gè)取樣周期間平均氣溫、平均最低氣溫、平均最高氣溫和累積降雨量。采用Excel 2010和SPSS軟件對(duì)各齡期群體動(dòng)態(tài)、氣溫和降雨信息進(jìn)行統(tǒng)計(jì)分析。
2 結(jié)果與分析
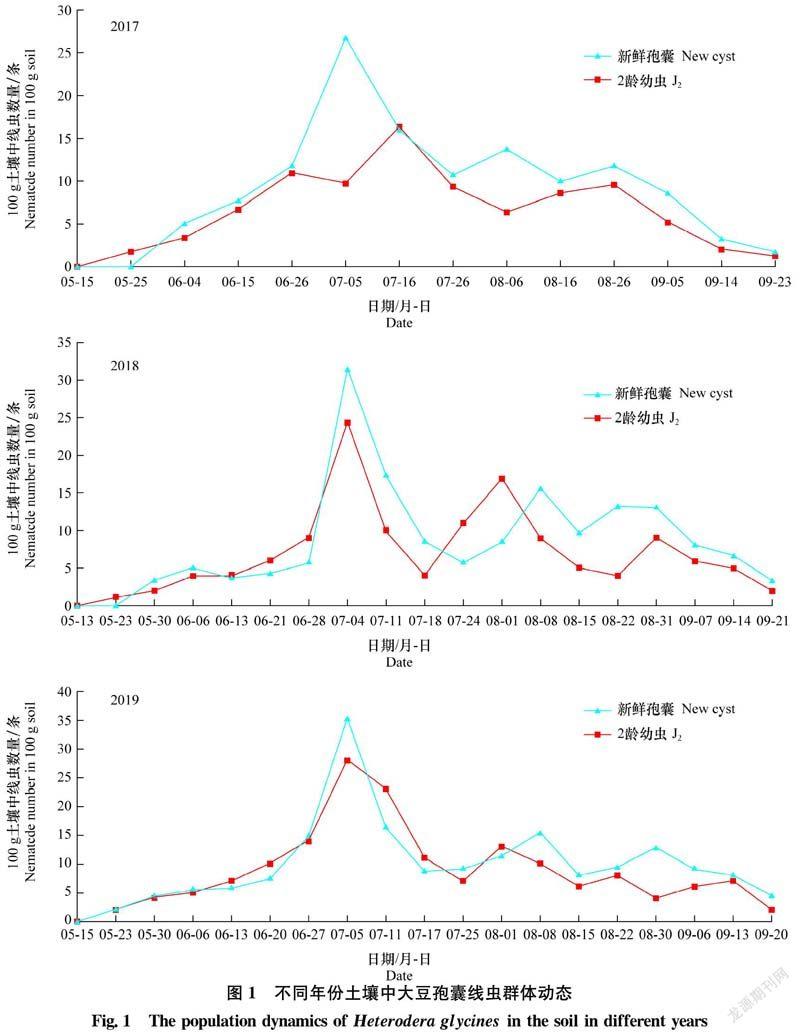
2.1 土壤中大豆孢囊線蟲群體動(dòng)態(tài)
2017年-2019年3個(gè)大豆生長(zhǎng)季對(duì)土壤中孢囊線蟲J2和新鮮孢囊群體密度變化情況的觀察表明,100 g土壤中新鮮孢囊和J2數(shù)量的變化趨勢(shì)較為吻合,有輕微起伏(圖1)。2017年新鮮孢囊出現(xiàn)3個(gè)高峰,2018年-2019年新鮮孢囊出現(xiàn)3個(gè)明顯高峰和1個(gè)不明顯高峰。2017年-2019年新鮮孢囊數(shù)最高值分別出現(xiàn)在7月5日、7月4日和7月5日,100 g土中分別有26.7、31.0個(gè)和35.3個(gè)。9月中旬,新鮮孢囊數(shù)量持續(xù)減少,孢囊數(shù)量為1~5個(gè)。此外,在2018年6月6日發(fā)生高峰不明顯,孢囊數(shù)僅為5個(gè)。
2017年-2019年,土壤中J2群體數(shù)量變化見圖1。2017年土壤中J2數(shù)量共出現(xiàn)3個(gè)明顯峰值,時(shí)間為6月26日、7月16日和8月26日。7月16日J(rèn)2密度最大,100 g土中有16.3條。生長(zhǎng)季后期,J2群體密度逐漸降低,數(shù)量極少,100 g土只有2條。2018年-2019年,土壤中J2數(shù)量均出現(xiàn)4個(gè)明顯峰值,J2全年密度最大出現(xiàn)在7月4日和7月5日,為24.3條和28.0條,9月中旬土壤中J2發(fā)生水平低,僅為1~2條。
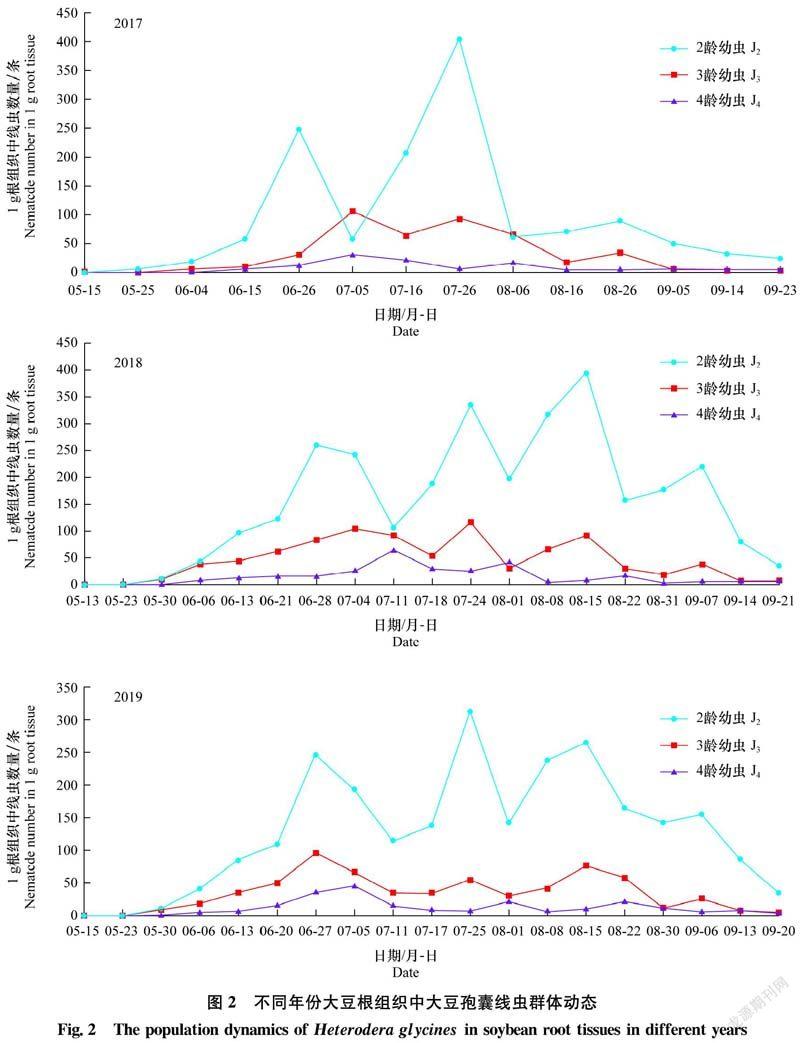
2.2 根組織中大豆孢囊線蟲群體動(dòng)態(tài)
2017年-2019年大豆根組織中大豆孢囊線蟲 J2、J3和J4群體密度變化情況見圖2。2017年J2密度有3個(gè)明顯高峰,其中7月26日密度最大,1 g根系中有404條。J3和J4亦有3個(gè)明顯高峰,密度最高為1 g根中有105條、31條。9月5日-14日根組織中J3和J4密度極小,1 g根中僅有3條。2018年和2019年,J2侵染數(shù)量均有4個(gè)明顯高峰,分別在8月15日和7月25日J(rèn)2數(shù)量最多,1 g根中分別有394條和312條。J3數(shù)量有4個(gè)明顯高峰,在7月24日和6月27日數(shù)量最多,1 g根中分別有115條和95條,持續(xù)時(shí)間較短。J4數(shù)量有4個(gè)明顯高峰,分別在7月11日和7月5日數(shù)量最多,分別為64條和45條。9月上旬以后根組織內(nèi)雖然再次出現(xiàn)J3和J4,但密度極小,僅有2~4條。
2.3 大豆孢囊線蟲的世代循環(huán)
2017年-2019年,從6月上旬至7月上旬,土壤中J2與新鮮孢囊發(fā)生水平低,結(jié)合根組織中J2侵入量、J3和J4(圖3,b~e)形成量大幅增加可以推測(cè)基本完成第1代。第1代J2侵入速度慢,根組織中各齡幼蟲發(fā)育遲緩,數(shù)量多,世代周期長(zhǎng),約42 d。
2017年7月5日至9月6日,根組織中J2、J3和J4均出現(xiàn)2個(gè)高峰,表明基本完成第2、3代,歷期分別為32 d和30 d。第1代和第2代內(nèi)各蟲態(tài)發(fā)生高峰順序與土壤中J2→根組織中J2、J3、J4→新鮮孢囊不相符,表明前2代存在較為明顯的世代重疊現(xiàn)象。第3代根內(nèi)J3、J4發(fā)育緩慢且數(shù)量極少,土壤中J2與新鮮孢囊數(shù)量呈下降趨勢(shì),說明第3代發(fā)育不完全。因此,2017年大豆孢囊線蟲發(fā)生3代。
2018年和2019年,從7月上旬至8月上旬,土壤中J2、根組織中各齡幼蟲與新鮮孢囊在不同時(shí)期發(fā)生不同程度的高峰,表明基本完成第2代,歷期28 d,根組織中J2、J3、J4發(fā)育較慢、世代周期較長(zhǎng)。第1代和第2代內(nèi)各蟲態(tài)發(fā)生高峰的順序并不規(guī)律,表明前2代存在較為明顯的世代重疊現(xiàn)象。第3代于8月下旬土壤中J2,根組織內(nèi)J2、J3、J4數(shù)量達(dá)到高峰,且土壤中產(chǎn)生大量新鮮孢囊,該世代歷期21 d基本完成,J2侵入速度快,根組織中幼蟲發(fā)育快且世代發(fā)生比較整齊。直至9月中旬根組織內(nèi)J2侵染數(shù)量達(dá)到高峰,但根內(nèi)J3、J4發(fā)育減慢,數(shù)量極少,土壤中J2出現(xiàn)峰值,完成第4代,歷期23 d。之后雖然在土壤中能檢測(cè)出少數(shù)J2,但對(duì)根組織的侵染基本停止。因此,2018年和2019年大豆孢囊線蟲發(fā)生4代。
每代根組織內(nèi)J4膨大突出根部表皮,進(jìn)一步發(fā)育為成熟雌蟲(圖3,f~g),雌蟲在體內(nèi)產(chǎn)卵。 后期雌蟲死亡,體壁漸漸增厚,形成褐色、檸檬形的孢囊(圖3h)脫落到土壤中。孢囊中的卵粒(圖3a)存活時(shí)間長(zhǎng)。由此可見,大豆孢囊線蟲每年第1代侵染時(shí)間基本一致,但在6月下旬后,因田間溫度和降雨量影響,大豆孢囊線蟲侵入和發(fā)育速率發(fā)生變化,2018年和2019年第2、3、4代歷期較短。
2.4 溫度和降雨對(duì)大豆孢囊線蟲繁殖的影響分析
該地區(qū)在大豆整個(gè)生長(zhǎng)季無人工灌溉,因此氣溫是影響大豆孢囊線蟲繁殖的主要因素。用SPSS軟件做Spearman相關(guān)分析,結(jié)果表明,平均溫度與J2、J3、J4數(shù)量的Spearman相關(guān)系數(shù)分別為0.747、0.737、0.608 (P<0.001),說明平均溫度(℃)和線蟲數(shù)量(條)存在正相關(guān)關(guān)系,即平均溫度升高與線蟲繁殖數(shù)量有關(guān)(表1)。而累積降雨量與J2、J3、J4數(shù)量的Spearman相關(guān)均為P>0.05,說明累積降雨量(mm)和線蟲數(shù)量(條)不存在相關(guān)關(guān)系,可能和降雨量不均勻有關(guān)。
3 討論
大豆孢囊線蟲在20~28℃內(nèi)隨溫度升高而發(fā)育速度加快,28℃為最適發(fā)育溫度,高于33℃幾乎不能發(fā)育[27]。在甘肅隴東,氣象數(shù)據(jù)表明,2017年7月份最高氣溫超過30℃的高溫日數(shù)多達(dá)22 d,平均最高氣溫29.0℃(21.8~38.5℃),持續(xù)時(shí)間較長(zhǎng),降雨量少,土壤呈半干旱狀況,土壤中J2數(shù)量顯著減少。2018年和2019年7月份最高氣溫超過30℃的高溫日有8 d,平均最高氣溫27.5℃(20.1~32.0℃),降雨量較2017年適中,土壤中J2數(shù)量明顯增加,這表明持續(xù)過高的溫度和過低的土壤含水量抑制大豆孢囊線蟲的侵染與生長(zhǎng)發(fā)育。通過Spearman相關(guān)分析表明,線蟲的繁殖數(shù)量與平均溫度正相關(guān),與降雨量無相關(guān)性,這與上述結(jié)果不一致,可能和最高氣溫持續(xù)期不同、降雨量分布不均勻有關(guān),土壤中含水量過低或過高都會(huì)影響線蟲的生長(zhǎng),導(dǎo)致線蟲代數(shù)不完全和世代重疊現(xiàn)象出現(xiàn),從而表現(xiàn)無相關(guān)性。降雨量和土壤含水量對(duì)大豆孢囊線蟲影響復(fù)雜,除了干旱和潮濕的影響,還應(yīng)分析水分脅迫強(qiáng)度及時(shí)間等[28],此部分內(nèi)容有待更深層次研究。此外,每年9月中旬基本完成最后1代,根組織內(nèi)各齡期幼蟲數(shù)量少,土壤中成熟孢囊數(shù)量少、個(gè)體小,這可能是大豆生長(zhǎng)季后期植株衰老、根組織木栓化和供給營(yíng)養(yǎng)少所致。
劉維志[6]指出,在中國(guó),大豆孢囊線蟲1年可發(fā)生3~4代或6代。趙洪海等[13]和齊娜偉等[14]的研究表明,山東省大豆孢囊線蟲在2013年發(fā)生3代,2014年發(fā)生4代,各世代平均歷期30 d。美國(guó)明尼蘇達(dá)州南部大豆孢囊線蟲1年發(fā)生3~4代,從7月到11月,土壤中孢囊數(shù)量呈上升趨勢(shì)。本研究表明,2017年甘肅隴東地區(qū)大豆孢囊線蟲發(fā)生3代,各代平均歷期較長(zhǎng),為35 d。2018年和2019年各代發(fā)育歷期較短,平均為28 d,發(fā)生4代。本文報(bào)道大豆孢囊線蟲世代平均歷期和世代數(shù)與上述報(bào)道略有差異,說明大豆孢囊線蟲世代歷期與當(dāng)?shù)販囟取⑵贩N抗性、土壤狀況和田間管理措施密切相關(guān)[12, 29]。本研究發(fā)現(xiàn),土壤中J2侵入點(diǎn)是新生伸長(zhǎng)組織,在側(cè)根和新生側(cè)根中各齡期線蟲數(shù)量多,主根中線蟲數(shù)量隨主根的生長(zhǎng)而減少。在線蟲大量孵化時(shí)期,一條新生須根中線蟲侵染量最多達(dá)53條。
大豆孢囊線蟲屬于土傳線蟲病害,危害程度重,傳播途徑多,發(fā)病區(qū)域廣。孢囊抗逆性較強(qiáng),能夠在干旱和低溫條件下長(zhǎng)期存活,防治極其困難[20, 30]。本研究結(jié)果表明,大豆孢囊線蟲第1代發(fā)生量最大,是主要危害期,此時(shí)大豆根系脆弱、抵抗力差,容易被線蟲侵染,因此應(yīng)以防治第1代發(fā)生為主,控制第2、3代侵染為輔。本課題組田間試驗(yàn)表明,播種前,溝施0.5%阿維菌素顆粒劑和采用生物種衣劑菌線克SN101、SN102進(jìn)行種子包衣處理均有一定防效[31]。大豆收獲后改種非寄主植物小麥、玉米、油菜和苜蓿等3~4年,每年可使土壤中孢囊基數(shù)減少20%左右[32]。
參考文獻(xiàn)
[1] JONES J T, HAEGEMAN A, DANCHINE C J, et al. Top 10 plant parasitic nematodes in molecular plant pathology [J]. Molecular Plant Pathology, 2013, 14(9): 946961.
[2] 孫玉秋, 許艷麗, 李春杰, 等. 黑龍江省大豆胞囊線蟲種群分布和卵密度研究[J]. 大豆科學(xué), 2011, 30(2): 250253.
[3] MAAFI Z T, SALATI M, RIGGS R D. Distribution, population density, race and type determination of soybean cyst nematode, Heterodera glycines, in Iran [J]. Nematology, 2008, 10(6): 919924.
[4] 段玉璽. 植物線蟲學(xué)[M]. 北京: 科學(xué)出版社, 2011.
[5] 陳井生. 黑龍江省大豆胞囊線蟲毒力類型分析及品種抗性研究[D]. 沈陽: 沈陽農(nóng)業(yè)大學(xué), 2019.
[6] 劉維志. 植物病原線蟲學(xué)[M]. 北京: 中國(guó)農(nóng)業(yè)出版社, 2000.
[7] PENG Deliang, PENG Huan, WU Duqing, et al. First report of soybean cyst nematode (Heterodera glycines) on soybean from Gansu and Ningxia China [J]. Plant Disease, 2016, 100(1): 229.
[8] WANG Dong, DUAN Yuxi, WANG Yuanyuan, et al. First report of soybean cyst nematode, Heterodera glycines, on soybean from Guangxi, Guizhou and Jiangxi provinces, China [J]. Plant Disease, 2015, 99(6): 893.
[9] SHI Hongli, ZHENG Jingwu. First report of soybean cyst nematode (Heterodera glycines) on tobacco in Henan, Central China [J]. Plant Disease, 2013, 97(6): 852.
[10]ZHENG Jingwu, ZHANG Yan, CHEN Senyu, et al. First report of the soybean cyst nematode, Heterodera glycines, on soybean in Zhejiang, Eastern China [J]. Plant Disease, 2009, 93(3): 319.
[11]李克梅, 日孜旺古麗, 董艷秋. 新疆大豆孢囊線蟲病的初步研究[J]. 植物保護(hù), 2014, 40(2): 132134.
[12]宋美靜, 朱曉峰, 王東, 等. 我國(guó)大豆主產(chǎn)區(qū)大豆胞囊線蟲群體分布及致病性分化研究[J]. 大豆科學(xué), 2016, 35(4): 630636.
[13]趙洪海, 丁海燕, 王鳳龍. 大豆和煙草上大豆孢囊線蟲田間侵染特征比較分析[J]. 中國(guó)農(nóng)業(yè)科學(xué), 2014, 47(22): 44174425.
[14]齊娜偉, 趙洪海. 2014年氣象條件對(duì)大豆孢囊線蟲田間侵染進(jìn)程的影響分析[J]. 山東農(nóng)業(yè)科學(xué), 2015, 47(5): 8185.
[15]項(xiàng)鵬, 張武, 李寶華, 等. 俄羅斯阿穆爾州大豆胞囊線蟲研究現(xiàn)狀[J]. 黑龍江農(nóng)業(yè)科學(xué), 2018(8): 126127.
[16]劉曉英. 黑龍江省東部大豆孢囊線蟲發(fā)生規(guī)律與防治技術(shù)研究[D]. 哈爾濱: 東北農(nóng)業(yè)大學(xué), 2003.
[17]陳金堂, 李知. 為害地黃的大豆孢囊線蟲的初步研究[J]. 植物病理學(xué)報(bào), 1981, 11(1): 3744.
[18]楊茹萍, 張國(guó)宏, 王立明, 等. 甘肅省大豆主產(chǎn)區(qū)產(chǎn)量性狀及品質(zhì)分析[J]. 大豆科學(xué), 2013, 32(1): 5055.
[19]羅寧, 李惠霞, 郭靜, 等. 甘肅省隴東南大豆孢囊線蟲的發(fā)生和分布[J]. 植物保護(hù), 2019, 43(3): 165169.
[20]李秀花, 馬娟, 陳書龍, 等. 不同植物對(duì)大豆孢囊線蟲種群的影響[J]. 河北農(nóng)業(yè)科學(xué), 2010, 14(8): 109111.
[21]孫晶雙, 鄭雅楠, 段玉璽, 等. 溫度、光照和土壤含水量對(duì)大豆胞囊線蟲休眠卵孵化的影響[J]. 大豆科學(xué), 2009, 28(4): 690692.
[22]董秋月, 鄭玉華, 趙洪海. 土壤水分對(duì)煙草孢囊線蟲侵染和煙堿含量的影響[J]. 中國(guó)煙草科學(xué),2015, 36(5): 6468.
[23]朱艷, 陳立杰, 段玉璽. 不同耕作方式對(duì)大豆胞囊線蟲群體數(shù)量的影響[J]. 大豆科學(xué), 2007, 26(2): 208212.
[24]靳學(xué)慧, 辛惠普, 鄭雯, 等. 長(zhǎng)期輪作和連作對(duì)土壤中大豆胞囊線蟲數(shù)量的影響[J]. 中國(guó)油料作物學(xué)報(bào), 2006, 28(2): 189193.
[25]許艷麗, 李春杰, 劉金波, 等. 東北黑土區(qū)重迎茬對(duì)大豆病蟲害發(fā)生的影響[J]. 大豆科學(xué), 2008, 27(3): 471474.
[26]路亞奇, 焦美齡, 曹彥超. 近35年甘肅慶陽黃土高原地帶氣候特征分析[J]. 中國(guó)農(nóng)學(xué)通報(bào), 2015, 31(29): 181190.
[27]劉維志. 植物線蟲研究技術(shù)[M]. 沈陽: 遼寧科學(xué)技術(shù)出版社,1995.
[28]趙洪海, 梁晨, 許冠堂, 等. 土壤濕度對(duì)大豆孢囊線蟲侵染和繁殖的影響[J]. 青島農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版), 2017, 34(1): 912.
[29]DUAN Yuxi, ZHENG Yanan, CHEN Lijie, et al. Effects of abiotic environmental factors on soybean cyst nematode [J]. Agricultural Sciences in China, 2009, 8(3): 317325.
[30]吳海燕, 段玉璽, 李秀俠. 大豆孢囊線蟲的休眠[J]. 動(dòng)物學(xué)報(bào), 2006, 52(3): 498503.
[31]羅寧, 劉永剛, 李惠霞, 等. 不同殺線劑對(duì)大豆孢囊線蟲病的防治效果[J]. 陜西農(nóng)業(yè)科學(xué), 2020, 66 (1): 1014.
[32]袁明. 黑龍江省西部大豆胞囊線蟲病發(fā)生動(dòng)態(tài)及防治對(duì)策[J]. 黑龍江農(nóng)業(yè)科學(xué), 2011(5): 4748.
(責(zé)任編輯:田 喆)
猜你喜歡
農(nóng)業(yè)科技通訊(2023年1期)2023-02-12 07:09:18
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國(guó)化肥信息(2022年7期)2022-08-31 01:29:28
中國(guó)化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農(nóng)產(chǎn)品市場(chǎng)周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報(bào)·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
