甘肅省部分地區(qū)馬鈴薯晚疫病菌交配型及線粒體DNA單倍型分析
2022-04-04 12:55:33惠娜娜王立李繼平鄭果李培玲馬生彪呂昭龍
植物保護(hù) 2022年2期
惠娜娜 王立 李繼平 鄭果 李培玲 馬生彪 呂昭龍

摘要 由致病疫霉Phytophthora infestans (Mont.) de Bary侵染引起的馬鈴薯晚疫病是一種世界范圍內(nèi)的毀滅性病害。為了明確甘肅省馬鈴薯晚疫病菌的遺傳結(jié)構(gòu),對(duì)2018年甘肅省定西市、張掖市和隴南市采集的130株晚疫病菌進(jìn)行了交配型和線粒體DNA (mtDNA)單倍型測(cè)定。結(jié)果表明:從130株晚疫病菌中檢測(cè)到A1和A2兩種交配型,A1交配型為14株(10.77%),A2交配型為116株(89.23%);檢測(cè)到Ⅰa、Ⅱa和Ⅱb 3種線粒體 DNA單倍型,發(fā)生頻率分別為88.46%、3.85%和7.69%。Ⅰa單倍型菌株的交配型為A1和A2,Ⅱa單倍型菌株交配型為A1和A2,Ⅱb單倍型菌株交配型為A1。該結(jié)果表明甘肅省部分地區(qū)馬鈴薯晚疫病菌以A2交配型,Ⅰa mtDNA單倍型為主。甘肅省各采集地晚疫病菌群體結(jié)構(gòu)或與當(dāng)?shù)胤N薯繁育、調(diào)運(yùn)有關(guān)。
關(guān)鍵詞 馬鈴薯; 晚疫病菌; 交配型; 線粒體DNA單倍型
中圖分類號(hào): S435.32
文獻(xiàn)標(biāo)識(shí)碼: A
DOI: 10.16688/j.zwbh.2021037
Abstract Potato late blight caused by Phytophthora infestans (Mont.) de Bary is a destructive disease in the world. In order to clarify the genetic structure of P.infestans in Gansu province, the mating type and mtDNA haplotype of 130 P.infestans strains collected in Dingxi city, Zhangye city and Longnan city in 2018 were determined. The results showed that A1 and A2 mating types were detected from 130 strains, among which 14 strains (10.77%) were the A1 mating type and 116 strains (89.23%) were A2. Three mtDNA haplotypes of Ⅰa, Ⅱa and Ⅱb were detected from 130 strains, and the occurrence frequencies were 88.46%, 3.85% and 7.69%, respectively. The mating types of Ⅰa haplotype strains were A1 and A2, Ⅱa haplotype strains were A1 and A2, and Ⅱb haplotype strains were A1. The results showed that the main pathogens of P.infestans in Gansu province were A2 mating type and Ⅰa mtDNA haplotype. The population structure of P.infestans in different collection sites in Gansu province may be related to the breeding and transportation of local seed potatoes.
Key words potato; Phytophthora infestans; mating type; mtDNA haplotype
由致病疫霉Phytophthora infestans (Mont.) de Bary侵染引起的馬鈴薯晚疫病在甘肅省的發(fā)生日趨嚴(yán)重,是當(dāng)前制約馬鈴薯產(chǎn)業(yè)健康可持續(xù)發(fā)展的最為嚴(yán)重的病害之一,其危害性高、防治難度大,對(duì)社會(huì)造成嚴(yán)重影響,現(xiàn)已被視為全球第一大作物病害[12]。馬鈴薯晚疫病菌是有A1和A2兩種交配型的異宗配合卵菌,不同交配型菌株可進(jìn)行有性生殖產(chǎn)生卵孢子。自1956年墨西哥首次發(fā)現(xiàn)A2交配型,世界各地陸續(xù)發(fā)現(xiàn)了A2交配型。張志銘等[3]1996年首次報(bào)道在我國山西和內(nèi)蒙古檢測(cè)到A2交配型,隨后在河北、四川、云南等地相繼檢測(cè)到了A2交配型[45]。A2交配型菌株的存在大大增強(qiáng)了晚疫病菌的適應(yīng)性和變異性[3]。
馬鈴薯晚疫病菌群體結(jié)構(gòu)的組成與變化對(duì)馬鈴薯晚疫病的發(fā)生、流行具有重要作用[6]。致病疫霉mtDNA為伴母性遺傳,序列簡單、相對(duì)保守,是研究致病疫霉起源進(jìn)化以及系統(tǒng)發(fā)育的理想對(duì)象[7]。國外研究表明晚疫病菌群體mtDNA單倍型可劃分為4種,即Ⅰa,Ⅱa,Ⅰb和Ⅱb[8]。Fry等[910]及Goodwin等[1112]研究認(rèn)為Ⅰb代表致病疫霉在第二次全球遷移前與US-1無性繁殖譜系密切相關(guān)的“舊”群體,而Ⅰa、Ⅱa和Ⅱb為致病疫霉第二次全球遷移發(fā)生后在墨西哥以外出現(xiàn)的“新”群體。研究晚疫病菌mtDNA單倍型可以揭示晚疫病菌遺傳結(jié)構(gòu)的演變,監(jiān)控晚疫病菌群體的變異情況。
馬鈴薯是我國西北地區(qū)農(nóng)民賴以生存的重要糧食作物和經(jīng)濟(jì)作物。甘肅省是我國重要的馬鈴薯種薯和商品薯繁殖基地及淀粉加工基地[13],2020年甘肅省馬鈴薯種植面積已超68.4萬hm2[14]。近年來由于種植結(jié)構(gòu)調(diào)整、氣候變化等因素的影響,馬鈴薯晚疫病頻繁流行,給馬鈴薯產(chǎn)業(yè)造成極大的損失,威脅甘肅省馬鈴薯產(chǎn)業(yè)的綠色健康可持續(xù)發(fā)展。本文對(duì)甘肅省馬鈴薯晚疫病菌的交配型和mtDNA單倍型開展相關(guān)研究,明確晚疫病菌交配型、mtDNA單倍型的組成與分布,初步探明了馬鈴薯晚疫病菌群體結(jié)構(gòu)的變化,為進(jìn)一步系統(tǒng)研究甘肅省晚疫病菌的群體結(jié)構(gòu)提供了技術(shù)支撐,同時(shí)為甘肅省馬鈴薯晚疫病病害綜合防控策略的制定提供參考。
1 材料與方法
1.1 試驗(yàn)材料
供試菌株:2018年采集自甘肅省定西市、張掖市和隴南市等地縣的馬鈴薯種植生產(chǎn)田新發(fā)病田塊,采集具有典型晚疫病癥狀的新鮮葉片,裝入紙袋置于冰盒中,帶回實(shí)驗(yàn)室,通過單孢分離[15]培養(yǎng)獲得的130株晚疫病菌。
供試培養(yǎng)基為黑麥番茄培養(yǎng)基[15](60 g黑麥加500 mL蒸餾水煮沸1 h,過濾,濾液加100 mL 番茄汁,0.4 g CaCO3,20 g蔗糖,11 g 瓊脂粉,蒸餾水定容至1 000 mL,分裝,121℃滅菌)。
1.2 馬鈴薯晚疫病菌交配型測(cè)定
采用皿內(nèi)對(duì)峙培養(yǎng)的方法。將待測(cè)菌株分別與標(biāo)準(zhǔn)菌株對(duì)峙接種在直徑9 cm的黑麥番茄汁培養(yǎng)基平板上,菌餅之間距離1.5 cm,在18℃黑暗條件下培養(yǎng)14 d左右,用顯微鏡檢查卵孢子的產(chǎn)生。同時(shí)將待測(cè)菌株單獨(dú)接于培養(yǎng)基上,同等條件下培養(yǎng),顯微鏡檢查。待測(cè)菌株與A1標(biāo)準(zhǔn)菌株產(chǎn)生卵孢子的為A2交配型;與A2標(biāo)準(zhǔn)菌株產(chǎn)生卵孢子的為A1交配型;與A1、A2標(biāo)準(zhǔn)菌株均能產(chǎn)生卵孢子的為A1A2交配型;不需交配即能產(chǎn)生卵孢子的為自育型菌株。標(biāo)準(zhǔn)菌株A1(NL80029)和A2(NL88133)由西北農(nóng)林科技大學(xué)單衛(wèi)星教授提供。
1.3 馬鈴薯晚疫病菌線粒體 DNA單倍型測(cè)定
1.3.1 基因組DNA的提取
采用OMEGA試劑盒提取基因組DNA。取純化培養(yǎng)7 d的致病疫霉菌株平板,刮下菌絲置于干凈的離心管-20℃冷凍保存。將保存的冷凍菌絲倒入加有液氮的研缽充分研磨成粉末,利用試劑盒提取DNA,所得DNA經(jīng)1%瓊脂糖凝膠電泳檢測(cè)。利用Q5000超微量核酸蛋白測(cè)定儀,檢測(cè)DNA的純度和濃度。
1.3.2 引物序列
利用Griffith等[8]的PCR-RFLP方法分析線粒體DNA單倍型。引物序列F2:5′-TCCCTTTGTCCTCTACCHAT-3′;R2:5′-TACGGCGGTTTAGCACATACA-3′;F4:5′-GGTCATCCAGAGGTTTATGTT-3′;R4:5′-CGATACCGATACCAGCACCAA-3′。引物由西安擎科生物工程有限公司合成。
1.3.3 PCR擴(kuò)增體系和程序
PCR擴(kuò)增體系(25 μL):2×TsingKe Master Mix 12.5 μL,100 μmol/L F2/R2或F4/R4引物各1 μL, 50 ng/μL 的基因組DNA 1 μL,用超純水定容至25 μL。PCR擴(kuò)增程序?yàn)椋?4℃預(yù)變性90 s;94℃變性40 s,55℃退火60 s,72℃延伸90 s,40個(gè)循環(huán);72℃下延伸120 s。PCR產(chǎn)物于1.2%的瓊脂糖凝膠中電泳檢測(cè),凝膠成像儀中拍照。本試驗(yàn)所用2×TsingKe Master Mix購于西安擎科生物工程有限公司。
1.3.4 酶切分析
利用Msp Ⅰ 和EcoR Ⅰ 分別對(duì)F2/R2和F4/R4兩對(duì)引物PCR產(chǎn)物(P2、P4)進(jìn)行酶切4 h以上(37℃)。酶切體系為50 μL:PCR產(chǎn)物10 μL,Buffer 5 μL,限制性內(nèi)切酶 1 μL,超純水補(bǔ)足至50 μL。酶切產(chǎn)物于2%的瓊脂糖凝膠電泳中檢測(cè),凝膠成像儀中拍照。根據(jù)酶切條帶大小判斷屬于何種基因型。本試驗(yàn)所用限制性內(nèi)切酶Msp Ⅰ和EcoR Ⅰ均購自生工生物工程(上海)股份有限公司。
1.4 數(shù)據(jù)分析
采用Microsoft Excel 2010進(jìn)行數(shù)據(jù)統(tǒng)計(jì)分析。
2 結(jié)果與分析
2.1 馬鈴薯晚疫病菌交配型組成分析
交配型測(cè)定結(jié)果表明(表1),130株晚疫病菌中有10.77%為A1交配型菌株(14株),89.23%為A2交配型菌株(116株)。其中A1交配型均來自張掖市,A2交配型來自定西市和隴南市,各采集地菌株交配型結(jié)構(gòu)相對(duì)單一。
寄主品種分析結(jié)果顯示:A1交配型菌株寄主為‘荷蘭15號(hào)’和‘克新13號(hào)’。A2交配型菌株寄主為‘隴薯6號(hào)’‘冀張薯12號(hào)’‘隴薯10號(hào)’‘隴薯7號(hào)’‘隴薯3號(hào)’‘青薯9號(hào)’和‘隴薯14號(hào)’。
2.2 馬鈴薯晚疫病菌線粒體DNA單倍型組成分析
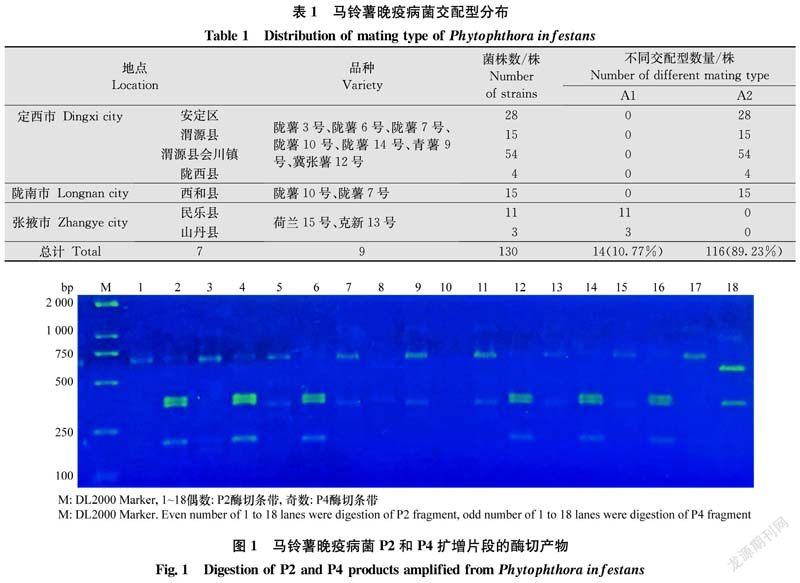
線粒體單倍型測(cè)定結(jié)果表明(圖1、表2):130株晚疫病菌中檢測(cè)到Ⅰa、Ⅱa和Ⅱb 3種mtDNA單倍型,Ⅰa單倍型為115株,Ⅱa單倍型為5株,Ⅱb單倍型為10株,頻率分別為88.46%、3.85%和7.69%。
從菌株來源分析,定西市采集分離的101株菌株包括Ⅰa和Ⅱa兩種單倍型,分別為98株(97.03%)和3株(2.97%,安定區(qū)),隴南市采集分離的15株菌株均為Ⅰa單倍型,張掖市采集分離的14株菌株共鑒定出Ⅰa、Ⅱa和Ⅱb 3種單倍型,分別為2株(14.28%)、2株(14.28%)和10株(71.44%),上述結(jié)果表明甘肅省的晚疫病菌mtDNA結(jié)構(gòu)較為復(fù)雜,不同地區(qū)晚疫病菌mtDNA單倍型組成差異較大。
2.3 馬鈴薯晚疫病菌交配型與線粒體 DNA單倍型相關(guān)性分析
晚疫病菌交配型和mtDNA單倍型測(cè)定結(jié)果表明:115株為Ⅰa單倍型,其所對(duì)應(yīng)的交配型有2株為A1交配型(1.74%),分布于張掖市民樂縣,113株為A2交配型(98.26%),分布于定西市、隴南市;5株菌株為Ⅱa單倍型,其所對(duì)應(yīng)的交配型有2株為A1交配型(40.00%),分布于張掖市山丹縣,3株為A2交配型(60.00%),分布于定西市安定區(qū);10株菌株為Ⅱb單倍型,其所對(duì)應(yīng)的交配型均為A1交配型,分布于張掖市民樂縣。
3 結(jié)論與討論
Niederhauser等[16]1956年在墨西哥中部首次發(fā)現(xiàn)了A2交配型,Hohl 1984年報(bào)道在瑞士發(fā)現(xiàn)A2交配型菌株,隨后在美洲、歐洲、非洲和亞洲馬鈴薯主產(chǎn)區(qū)陸續(xù)報(bào)道了A2交配型菌株[6]。Vargas 等[17] 2009 年報(bào)道了哥倫比亞首次發(fā)現(xiàn) A2交配型菌株,并測(cè)定出 A2 交配型菌株屬于Ⅰa 型線粒體 DNA 單倍型,而 A1 交配型菌株為Ⅱa 型。Casa-coila等[18]研究表明巴西南部馬鈴薯晚疫病菌存在A1、A2、A1A2和自育型交配型菌株,A2為59.73%。國內(nèi)學(xué)者研究表明中國北方地區(qū)(河北省、山東省、黑龍江省、內(nèi)蒙古)主要以A1交配型、無性繁殖為主[1924],南方地區(qū)(湖北省、貴州省、云南省)以A2交配型為主[24]。部分學(xué)者在2003年-2015年對(duì)甘肅省馬鈴薯晚疫病菌交配型進(jìn)行了相關(guān)研究[2233],檢測(cè)到了A1、A2、A1A2和自育型,不同年份不同采集地交配型組成存在明顯差異。本研究對(duì)2018年采集自甘肅省定西市、張掖市和隴南市的130株晚疫病菌交配型分析表明,甘肅省馬鈴薯晚疫病菌以A2交配型為主,各地區(qū)交配型組成相對(duì)單一,該結(jié)果說明甘肅省馬鈴薯主產(chǎn)區(qū)晚疫病菌主要以無性繁殖為主,這一結(jié)果與國內(nèi)學(xué)者對(duì)甘肅地區(qū)馬鈴薯晚疫病菌交配型研究結(jié)果基本一致[2932]。李繼平[31]2013年報(bào)道了2007年-2012年甘肅省晚疫病菌交配型存在A1、A2、A1A2和自育型,2008年發(fā)現(xiàn)A2交配型,之后A2交配型菌株比例上升,自育型菌株在2007年和2008年檢測(cè)到,A1A2菌株在2011年和2012年發(fā)現(xiàn),本研究未檢測(cè)到A1A2和自育型菌株,這或與采樣點(diǎn)有關(guān)。不同年度間甘肅省馬鈴薯晚疫病菌交配型組成不同,以定西地區(qū)為例,2007年-2012年以A1交配型為主(73.08%),2018年均為A2交配型,A2交配型菌株的增多可能意味著馬鈴薯晚疫病菌容易發(fā)生基因重組,可能形成致病力更強(qiáng)的菌系,從而增加抗性菌株、新生理小種出現(xiàn)的頻率,馬鈴薯品種抗性喪失等等問題,造成馬鈴薯晚疫病的發(fā)生日趨嚴(yán)重。因此,應(yīng)對(duì)甘肅省馬鈴薯晚疫病菌持續(xù)開展交配型、抗藥性、生理小種等監(jiān)測(cè)工作。
由于致病疫霉線粒體DNA結(jié)構(gòu)相對(duì)簡單且穩(wěn)定性良好,因此被廣泛用于致病疫霉起源和演化的研究。Day等[34]研究表明1995年-1998年蘇格蘭、英格蘭、威爾士等地馬鈴薯晚疫病菌為Ⅰa和Ⅱa單倍型。Winton等[35]表明美國阿拉斯加晚疫病菌為Ⅱa單倍型。趙志堅(jiān)等[36]發(fā)現(xiàn)2001年云南馬鈴薯晚疫病菌為Ⅰa單倍型。郭軍等[37]研究表明內(nèi)蒙古馬鈴薯晚疫病菌交配型為A1,群體mtDNA單倍型均為Ⅱa型。馬云芳[38]研究表明2009年采集自甘肅的菌株中發(fā)現(xiàn)Ⅰa、Ⅱa單倍型,寧夏的菌株中發(fā)現(xiàn)Ⅰa、Ⅱa和Ⅱb,陜西的菌株中發(fā)現(xiàn)Ⅱa;2010年甘肅菌株為Ⅰa單倍型,寧夏的菌株中發(fā)現(xiàn)Ⅰa、Ⅱa和Ⅱb單倍型,陜西的菌株中發(fā)現(xiàn)Ⅱa。楊策[39]研究發(fā)現(xiàn)2010年-2012年貴州菌株發(fā)現(xiàn)Ⅰa和Ⅱb單倍型,云南菌株發(fā)現(xiàn)Ⅱa,福建、武漢菌株發(fā)現(xiàn)Ⅰa、Ⅱa和Ⅱb單倍型,寧夏菌株中發(fā)現(xiàn)Ⅰa和Ⅱa,甘肅菌株為Ⅰa型。Tian等[40]2016年報(bào)道了甘肅省馬鈴薯晚疫病菌的mtDNA單倍型為Ⅰa和Ⅱa單倍型,其中Ⅰa為優(yōu)勢(shì)基因型。李璐[23]研究表明黑龍江菌株為Ⅰa和Ⅱa單倍型,內(nèi)蒙古菌株為Ⅱa單倍型。惠娜娜[41]對(duì)2007年-2012年采集自甘肅的45株晚疫病菌進(jìn)行mtDNA單倍型檢測(cè),發(fā)現(xiàn)Ⅰa和Ⅱa 單倍型,其中Ⅰa比例高。本研究表明甘肅省馬鈴薯晚疫病菌檢測(cè)到了Ⅰa、Ⅱa和Ⅱb 3種mtDNA單倍型,其中Ⅰa為優(yōu)勢(shì)基因型,在張掖市民樂縣發(fā)現(xiàn)了Ⅱb單倍型。這一結(jié)果與國內(nèi)學(xué)者對(duì)西北地區(qū)致病疫霉群體單倍型結(jié)構(gòu)研究結(jié)果略有不同。據(jù)Fry等及Goodwin等的假說,致病疫霉“新”群體已成為甘肅省馬鈴薯產(chǎn)區(qū)的主導(dǎo)者,“新”群體對(duì)晚疫病的防治帶來新的挑戰(zhàn)。
甘肅省馬鈴薯晚疫病菌群體遺傳結(jié)果或與種薯調(diào)運(yùn)有關(guān)。定西市和隴南市等地以定西地區(qū)(渭源、安定)繁育種薯為主,各地晚疫病菌交配型和mt DNA單倍型相對(duì)一致,該地晚疫病菌以A2交配型,Ⅰa單倍型菌株為主,這與之前甘肅定西地區(qū)菌株交配型的結(jié)果略有不同,但mtDNA單倍型組成與之前結(jié)果基本一致;河西地區(qū)(張掖市)以自繁、內(nèi)蒙古等地調(diào)種為主,該地晚疫病菌以A1交配型,Ⅰa、Ⅱ a和Ⅱb單倍型菌株組成,山丹縣采集的菌株與郭軍等[20,37]、李璐[23]對(duì)內(nèi)蒙古馬鈴薯晚疫病菌(以A1交配型,Ⅱ a單倍型為主)的研究結(jié)果一致,mt DNAⅡb單倍型菌株采自于當(dāng)?shù)刈苑狈N薯品種。本研究結(jié)果表明甘肅省部分馬鈴薯產(chǎn)區(qū)馬鈴薯晚疫病主要以種薯帶菌和種薯調(diào)運(yùn)為主要初侵染源和傳播途徑。因此,甘肅省對(duì)馬鈴薯晚疫病的防控應(yīng)以種薯處理為主,通過對(duì)種薯進(jìn)行藥劑處理來消除或減少病原,以達(dá)到控制晚疫病的發(fā)生和流行,同時(shí)今后需要擴(kuò)大晚疫病菌采集點(diǎn),加強(qiáng)對(duì)不同生態(tài)區(qū)晚疫病菌群體遺傳結(jié)構(gòu)的監(jiān)測(cè),為下一步晚疫病的防控提供理論支撐。
參考文獻(xiàn)
[1] JAMES W C. Estimated losses of crops from plant pathogens [M]∥ PIMENTEL D. CRC handbook of pest management in agriculture (USA), 1981.
[2] ERWIN D C, ROBERIO K. Phytophthora diseases worldwide [M]. Saint Paul, MN, USA: APS Press, 1996: 562.
[3] 張志銘, 李玉琴, 田世民, 等. 中國發(fā)生馬鈴薯晚疫病菌(Phytophthora infestans) A2交配型[J]. 河北農(nóng)業(yè)大學(xué)學(xué)報(bào), 1996, 19(4): 6265.
[4] 朱杰華, 張志銘, 李玉琴. 馬鈴薯晚疫病菌(Phytophthora infestans)A2交配型的分布[J]. 植物病理學(xué)報(bào), 2000, 30(4): 375.
[5] 閔凡祥, 王曉丹, 胡林雙, 等. 黑龍江省馬鈴薯晚疫病菌交配型的研究[J]. 中國馬鈴薯, 2010, 24(1): 4749.
[6] 朱杰華. 中國馬鈴薯晚疫病菌群體遺傳結(jié)構(gòu)研究[D]. 保定: 河北農(nóng)業(yè)大學(xué), 2004.
[7] 趙志堅(jiān), 曹繼芬, 李燦輝, 等. 云南致病疫霉交配型、甲霜靈敏感性、mtDNA單倍型及其群體演替研究[J]. 中國農(nóng)業(yè)科學(xué), 2007, 40(4): 727734.
[8] GRIFFITH G W, SHAW D S. Polymorphisms in Phytophthora infestans: Four mitochondrial haplotypes are detected after PCR amplification of DNA from pure cultures or from host lesions [J]. Applied and Environmental Microbiology, 1998, 64(10): 40074014.
[9] FRY W E, GOODWIN S B. Resurgence of the Irish potato famine fungus [J]. Bioscience, 1997, 47(6): 363371.
[10]FRY W E, GOODWIN S B, DYER A T, et al. Historical and recent migrations of Phytophthora infestans: chronology, pathways, and implications [J]. Glia, 1993, 45(2):155169.
[11]GOODWIN S B. The population genetics of Phytophthora [J]. Phytopathology, 1997, 87(4): 462473.
[12]GOODWIN S B, COHEN B A, FRY W E. Panglobal distribution of a single clonal lineage of the Irish potato famine fungus [J]. Proceedings of the National Academy of Sciences of the United States of America, 1994, 91(24): 1159111595.
[13]何三信, 文國宏, 王一航, 等. 甘肅省馬鈴薯產(chǎn)業(yè)現(xiàn)狀及提升措施建議[J]. 中國馬鈴薯, 2010, 24(1): 54 57.
[14]趙記軍, 吳正強(qiáng), 董博.甘肅馬鈴薯產(chǎn)業(yè)現(xiàn)狀與發(fā)展對(duì)策[J].甘肅農(nóng)業(yè)科技, 2021, 52(4):7782.
[15]GUO Liyun, YANG Yanli, LUO Wenfu. Mating type and biological characteristics of Phytophthora infestans isolates from Yunnan, China [J]. 植物病理學(xué)報(bào), 2002, 32 (1): 4954.
[16]NIEDERHAUSER J S. Division of mycology: the blight, the blighter, and the blighted [J]. Transactions of the New York Academy of Sciences, 1956, 19(1):5563.
[17]VARGAS A M, OCAMPO L M Q, CSPEDES M C, et al. Characterization of Phytophthora infestans populations in Colombia: first report of the A2 mating type [J]. Phytopathology, 2009, 99 (1):8288.
[18]CASA-COILA V H, GOMES C B, LIMA-MEDINA I, et al. Characterization of mating type and the diversity of pathotypes of Phytophthora infestans isolates from Southern Brazil [J]. Journal of Plant Diseases and Protection, 2020, 127(1): 4354.
[19]LI Benjin, CHEN Qinghe, LV Xinin, et al. Phenotypic and genotypic characterization of Phytophthora infestans isolates from China [J]. Journal of Phytopathology, 2009, 157(9):558567.
[20]郭軍, 屈冬玉, 鞏秀峰, 等. 內(nèi)蒙古馬鈴薯晚疫病菌交配型和生理小種研究[J]. 西北農(nóng)林科技大學(xué)學(xué)報(bào)(自然科學(xué)版), 2007, 35(11): 120124.
[21]陳思慧. 中國馬鈴薯主產(chǎn)區(qū)晚疫病菌遺傳變異研究[D]. 呼和浩特:內(nèi)蒙古大學(xué), 2017.
[22]敖翔. 黑龍江省馬鈴薯晚疫病菌群體結(jié)構(gòu)及塊莖晚疫病抗性分析[D].哈爾濱:東北農(nóng)業(yè)大學(xué),2018.
[23]李璐. 致病疫霉群體結(jié)構(gòu)及誘導(dǎo)劑與殺菌劑對(duì)晚疫病防效研究[D].哈爾濱:東北農(nóng)業(yè)大學(xué),2019.
[24]張治斌. 中國馬鈴薯主產(chǎn)區(qū)晚疫病菌表型和SSR基因型分析[D].呼和浩特:內(nèi)蒙古大學(xué),2020.
[25]王英華, 國立耘, 梁德霖, 等. 馬鈴薯晚疫病菌在內(nèi)蒙古和甘肅的交配型分布及對(duì)幾種殺菌劑的敏感性[J]. 中國農(nóng)業(yè)大學(xué)學(xué)報(bào), 2003, 8(1): 7882.
[26]孫銀銀. 西北部分地區(qū)致病疫霉菌的交配型分布和遺傳多樣性初步分析[D]. 楊凌: 西北農(nóng)林科技大學(xué), 2010.
[27]惠娜娜, 李繼平, 李青青, 等. 甘肅馬鈴薯晚疫病病菌交配型及對(duì)甲霜靈的敏感性[J]. 甘肅農(nóng)業(yè)科技, 2010(10): 810.
[28]王生榮, 呂偉, 李惠霞. 甘肅馬鈴薯晚疫病菌交配型和對(duì)霜脲氰的敏感性測(cè)定 [C]∥ 2010年中國菌物學(xué)會(huì)學(xué)術(shù)年會(huì)論文摘要集. 長沙: 中國菌物學(xué)會(huì), 2010: 89.
[29]楊海萍. 甘肅省馬鈴薯晚疫病菌的群體遺傳多樣性分析[D]. 楊凌: 西北農(nóng)林科技大學(xué), 2011.
[30]李繼平. 甘肅馬鈴薯晚疫病菌群體結(jié)構(gòu)及病害治理技術(shù)研究[D]. 蘭州: 甘肅農(nóng)業(yè)大學(xué), 2013.
[31]李會(huì)平. 抗病性鑒定圃馬鈴薯晚疫病菌群體遺傳多樣性分析[D]. 楊凌: 西北農(nóng)林科技大學(xué), 2014.
[32]張大為, 惠娜娜, 王立, 等. 甘肅省馬鈴薯致病疫霉交配型組成及其對(duì)甲霜靈的抗藥性[J]. 西北農(nóng)業(yè)學(xué)報(bào), 2014, 23(6): 184188.
[33]方治國. 中國馬鈴薯晚疫病菌交配型與無毒基因多樣性分析[D].福州:福建農(nóng)林大學(xué), 2013.
[34]DAY J P,WATTIER R A M, SHAW D S, et al. Phenotypic and genotypic diversity in Phytophthora infestans on potato in Great Britain, 1995-1998 [J].Plant Pathology, 2004, 53(3):303315.
[35]WINTON L M, LEINER R H, KROHN A L, et al. Occurrence of late blight caused by Phytophthora infestans on potato and tomato in Alaska [J]. Plant Disease, 2007, 91(5):634.
[36]趙志堅(jiān),李先平,李成云,等.致病疫霉線粒體DNA單倍型的分子鑒別[J].云南農(nóng)業(yè)大學(xué)學(xué)報(bào), 2002(4):434435.
[37]郭軍, 屈冬玉, 鞏秀峰.內(nèi)蒙古馬鈴薯晚疫病菌基因型多樣性分析[J]. 西北農(nóng)林科技大學(xué)學(xué)報(bào)(自然科學(xué)版), 2007, 35(4):120124.
[38]馬云芳. 西北馬鈴薯主產(chǎn)區(qū)致病疫霉菌線粒體DNA單倍型分析[D]. 楊凌: 西北農(nóng)林科技大學(xué), 2012.
[39]楊策. 中國馬鈴薯晚疫病菌線粒體單倍型與溫度適應(yīng)性分析[D]. 福州: 福建農(nóng)林大學(xué), 2013.
[40]TIAN Y E, YIN J L, SUN J P, et al. Population genetic analysis of Phytophthora infestans in northwestern China [J]. Plant Pathology, 2016, 65 (1): 1725.
[41]惠娜娜. 甘肅省馬鈴薯晚疫病菌線粒體DNA單倍體型組成與分析[C]∥ 中國作物學(xué)會(huì)馬鈴薯專業(yè)委員會(huì). 2015年中國馬鈴薯大會(huì): 馬鈴薯產(chǎn)業(yè)與現(xiàn)代可持續(xù)農(nóng)業(yè), 2015: 426430.
(責(zé)任編輯:田 喆)
