馬唐種子萌發及幼苗建成對不同環境因子的響應
2022-04-04 12:55:33郭文磊張泰劼張純田興山
植物保護 2022年2期
郭文磊 張泰劼 張純 田興山
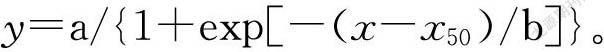

摘要 馬唐是我國常見的旱田雜草之一,然而近年來在部分地區直播稻田危害十分嚴重。本研究測定了馬唐種子萌發和幼苗建成過程中對不同環境因子的響應特征,以期為直播稻田馬唐綜合防控提供科學依據。結果表明:馬唐在L∥D=12 h∥12 h,光照/黑暗階段溫度分別為40℃/30℃、35℃/25℃、30℃/20℃、25℃/15℃、20℃/10℃的變溫條件下均可萌發,最適萌發溫度組合為30℃/20℃;光照不是馬唐種子萌發的必需條件,但對其萌發有一定的刺激作用;馬唐種子在pH 4~10條件下均可萌發,pH 7~10時萌發率均在90%以上;滲透勢為-0.6~0 MPa時馬唐種子萌發率在41.7%~98.3%,滲透勢為-0.8 MPa及以下時基本無法萌發;馬唐對鹽分脅迫的耐受能力較強,NaCl濃度為240 mmol/L時萌發率達20%;馬唐種子在土壤表面時出苗率最高(90%),埋藏深度6 cm時無法出苗;在水稻1~2葉期或2~3葉期保持2~3 cm水層并持續7 d以上,可使馬唐成苗率降至20%以下,與對照相比干重抑制率在95%以上,且對水稻生長無明顯影響。馬唐對不同環境因子具有較強的耐受能力,適度翻耕及建立合適的水層可能有助于減輕馬唐在直播稻田的危害。
關鍵詞 馬唐; 萌發; 幼苗建成; 水層; 環境因子
中圖分類號: S451.1
文獻標識碼: A
DOI: 10.16688/j.zwbh.2021007
Abstract Digitaria sanguinalis is one of the common weeds in upland fields in China. However, it has recently heavily infested some direct-sowing rice fields in parts of China. To provide scientific basis for integrated management of D.sanguinalis in direct-sowing rice fields, we analyzed and determined the responses of seed germination and seedling establishment to different environmental factors. The results showed that D.sanguinalis could germinate under the alternating day/night temperature of 40℃/30℃, 35℃/25℃, 30℃/20℃, 25℃/15℃, 20℃/10℃, and the maximum germination rate occurred at 30℃/20℃. Light was not an essential condition for D.sanguinalis germination, but it has a certain stimulating effect on germination of D.sanguinalis. Seed could germinate over a pH range of four to ten, and more than 90% germination rate was observed at pH values ranging from seven to ten. Seed germination rate was 41.7% to 98.3% when osmotic potential was -0.6-0 MPa, but it was almost completely inhibited at osmotic potential of -0.8 MPa. D.sanguinalis was tolerant to salt stress, and 20% germination rate occurred at 240 mmol/L NaCl. Seedling emergence rate was highest (90%) when burial depth was 0 cm and no seedlings emerged when burial depth was 6 cm. When water layer was established at one- to two-leaf or two- to three-leaf stage of Oryza sativa with 2-3 cm water layer depth for a continuous period of 7 d or longer, seedling survival rates were lower than 20% and dry weights of aerial parts of D.sanguinalis reduced by more than 95%, while O.sativa was not significantly affected. Overall, D.sanguinalis exhibited strong tolerance to different environmental factors, and appropriate tillage or water layer treatment might be helpful for reducing D.sanguinalis damage in direct-sowing rice fields.
Key words Digitaria sanguinalis; germination; seedling establishment; water layer; environmental factors
馬唐Digitaria sanguinalis是一年生禾本科植物,廣泛分布于全球熱帶、亞熱帶及溫帶地區,是我國玉米田、蔬菜田、棉花田、果園等作物田惡性雜草之一[12]。近年來,隨著水稻直播種植方式的不斷推廣,馬唐大量侵入直播稻田,危害逐漸加重并上升為部分地區稻田優勢雜草,如揚州市江都區水稻田馬唐發生密度占雜草總密度的22%以上[3]。馬唐屬雜草早期生長速度快,競爭力強,在低密度下也可帶來嚴重的產量損失,單株雜草可產生數萬粒種子,可通過風、水流、機械攜帶等方式快速傳播[4]。馬唐在華南地區可周年發生,對水田、旱田等不同田間環境均有較強適應能力,在上述環境下均可萌發并完成出苗,但目前有關不同環境因子下馬唐種子萌發及幼苗建成的系統研究較少。
化學除草是當前我國稻田最主要的除草方式,具有快速、高效、成本低等優點,但長期依賴化學除草也為稻田雜草防控帶來了新的問題。首先,栽培方式、除草劑使用等因素導致稻田雜草群落發生改變,馬唐等雜草在直播稻田大量發生。對于該類新形成的稻田優勢雜草,除草劑防控產品和技術較為缺乏[56]。王紅春等[7]調查發現,在水稻分蘗末期,隨著前期施用的土壤封閉除草劑藥效下降,馬唐仍可大量出苗,且生長迅速,水稻成熟前可完成生活史,對水稻產量和品質造成嚴重影響。筆者前期研究發現,水稻田常用禾本科雜草除草劑二氯喹啉酸、五氟磺草胺、雙草醚等對馬唐防效較差,僅乙酰輔酶A羧化酶(ACCase)抑制劑類除草劑氰氟草酯和噁唑酰草胺對2~3葉齡的馬唐有較好效果[8]。其次,ACCase抑制劑類除草劑作用位點單一,長期使用容易導致雜草抗藥性的產生。全球范圍內已報道49種雜草對ACCase抑制劑類除草劑產生了抗性[9],其中我國江蘇等地區的直播稻田馬唐種群已對該類除草劑產生了抗性[3]。通常雜草對ACCase類除草劑具有交互抗性[10],這進一步增加了稻田馬唐種群的防控難度,在缺乏新作用方式除草劑的背景下,豐富馬唐等稻田新形成優勢雜草的防控措施,減輕對化學除草劑的依賴,延長現有除草劑的使用周期,成為水稻直播種植模式中面臨的現實問題。對雜草萌發、幼苗建成等生物學特性進行研究有助于針對性地制定防控措施。
種子萌發并形成幼苗是植物適應自然環境形成種群的基礎,是雜草在田間造成危害的前提,同時也是雜草防控過程中最關鍵的階段[1112]。種子萌發受到多種環境因子的影響,如溫度、光照、pH、鹽濃度及滲透勢等[13]。例如,光照可刺激光頭稗Echinochloa colona等種子的萌發,因此其在土壤表層萌發率通常較高,在免/少耕田很可能危害更重[14]。有些雜草種子在干旱及高鹽土壤中仍可萌發,因此相比其他植物可獲得競爭優勢。在不同深度的土層中,濕度、溫度等存在差異,對雜草種子萌發和出苗有顯著影響[13]。水層是稻田控制雜草的重要手段,但直播稻田早期建立水層時間、水層深度、持續周期等因素對水稻和馬唐的不同影響尚未見報道。因此,本論文系統研究溫度、光照、pH、鹽濃度、滲透勢、埋藏深度、水層等環境因子對馬唐種子萌發和幼苗建成的影響,以期為直播稻田馬唐綜合防控提供科學依據。
1 材料與方法
1.1 材料
供試馬唐種子于2018年10月采自廣州市花都區花山鎮旱直播稻田,稻田總面積在3 hm2以上,在田間隨機采集至少500株的種子,采集后帶回實驗室自然條件下晾干備用。
1.2 方法
1.2.1 馬唐種子萌發對溫度的響應
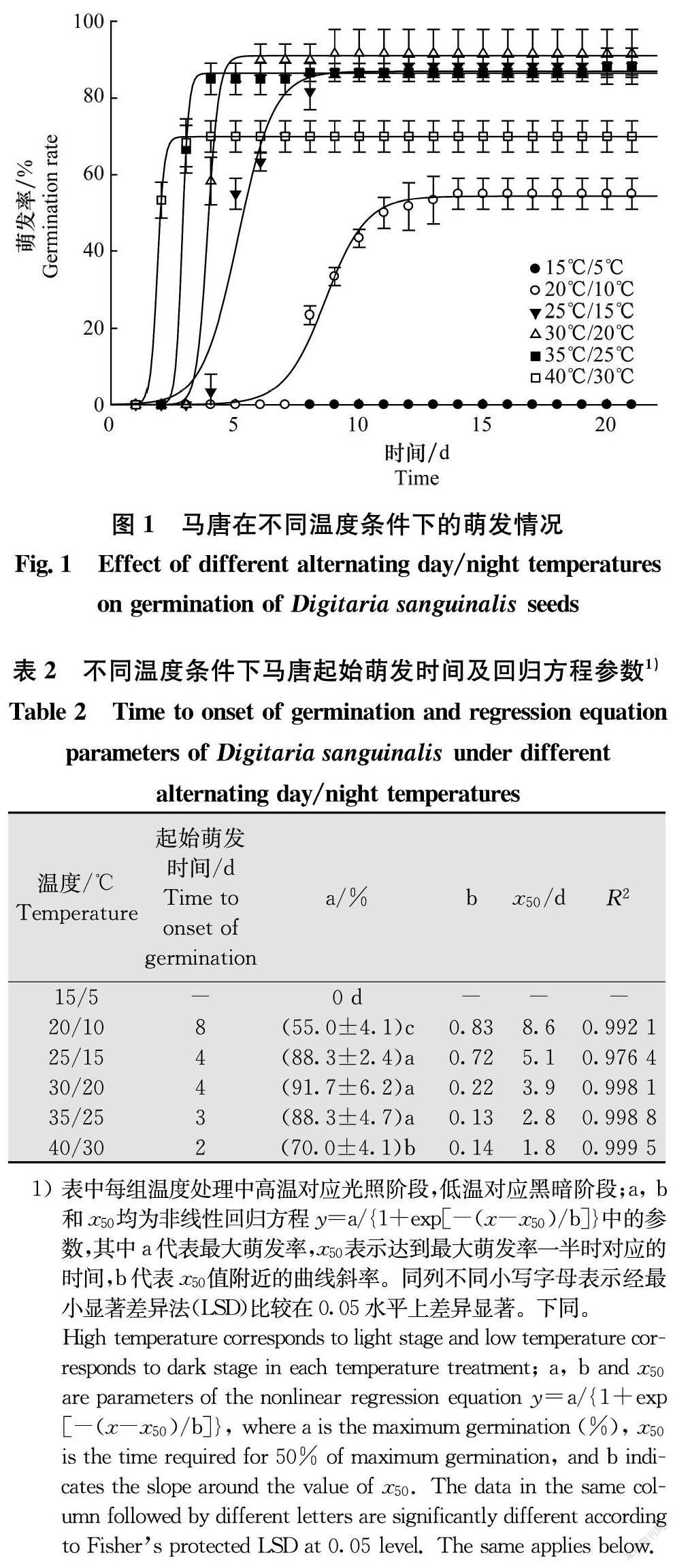
在直徑9 cm的培養皿中放入2層濾紙,加入去離子水5 mL,然后將20粒大小均勻、籽粒飽滿的馬唐種子均勻擺放在濾紙上,用保鮮膜將培養皿封口后放入光照培養箱(GXZ-280型,寧波江南儀器廠)中。光周期L∥D=12 h∥12 h,設置6組變溫處理,即40℃/30℃、35℃/25℃、30℃/20℃、25℃/15℃、20℃/10℃、15℃/5℃,光照階段與高溫相對應,光照強度為200 μmol/(m2·s)。每處理3次重復,每24 h統計1次發芽數,共統計21 d,胚根伸出種皮3 mm視為萌發[15]。
1.2.2 馬唐種子萌發對光照的響應
為明確光照對馬唐種子萌發的影響,以24 h為1個周期,設置3組處理:光照24 h/d、光照12 h/d、光照0 h/d。為保證無光照的環境,將光照0 h/d處理組的培養皿用3層鋁箔紙包裹。溫度條件為30℃/20℃變溫處理,其他條件同1.2.1。處理后21 d檢查萌發數。
1.2.3 馬唐種子萌發對pH的響應
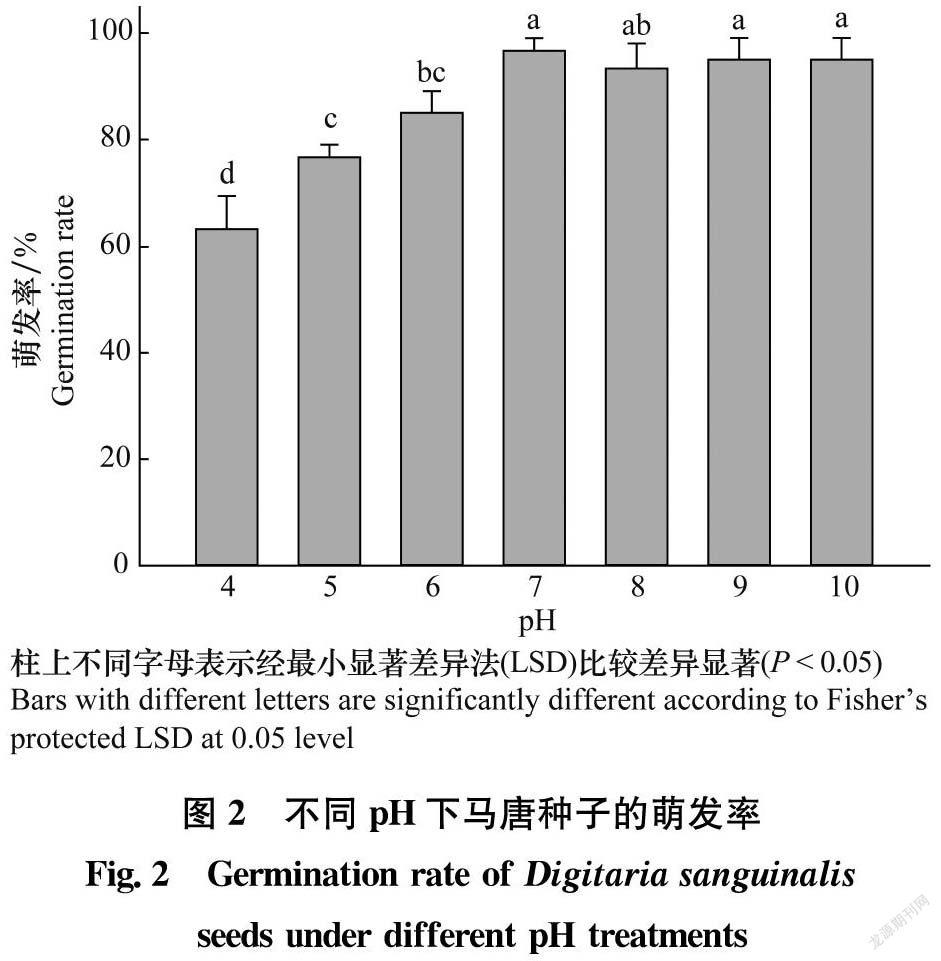
用不同pH的緩沖液代替去離子水進行馬唐種子萌發試驗。配制方法參考Chachalis等的方法[16],使用1 mol/L的HCl溶液調節2 mmol/L的鄰苯二甲酸氫鉀緩沖液至pH為4.0;用1 mol/L的NaOH溶液調節2 mmol/L的MES[2-(N-嗎啉)乙磺酸]緩沖液至pH為5.0和6.0;用1 mol/L的NaOH溶液調節2 mmol/L的HEPES[N-(2-羥甲基)哌嗪-N′-(2-乙磺酸)] 緩沖液至pH為7.0和8.0;用1 mol/L的NaOH溶液調節2 mmol/L的三甲基甘氨酸緩沖液至pH為9.0和10.0。溫度條件為30℃/20℃變溫處理,其他條件同1.2.1。處理后21 d檢查萌發數。
1.2.4 馬唐種子萌發對滲透勢的響應
參考Michel等的方法[17]。分別稱取0、72.5、112.2、143.2、169.4、192.6、213.6、251.0、284.0、299.3 g的PEG 8000溶于1 L去離子水中,即配制成滲透勢分別為0、-0.1、-0.2、-0.3、-0.4、-0.5、-0.6、-0.8、-1.0、-1.1的水溶液。用不同滲透勢的水溶液進行馬唐種子萌發試驗,溫度條件為30℃/20℃變溫處理,其他條件同1.2.1。處理后21 d檢查萌發數。
1.2.5 馬唐種子萌發對鹽分脅迫的響應
分別配制濃度為0、20、40、80、120、160、200、240、280 mmol/L和320 mmol/L的NaCl溶液,用于馬唐種子萌發試驗,溫度條件為30℃/20℃變溫處理,其他條件同1.2.1。處理后21 d檢查萌發數。
1.2.6 馬唐出苗對埋藏深度的響應
將取自當地農田的表層土壤(pH 6.25,有機質2.75%,速效氮160 mg/kg,速效磷146 mg/kg,速效鉀425 mg/kg)在170℃的烘箱中烘干3 h,過20目篩后裝入底部有孔的直徑16 cm、高13 cm的花盆中。在距花盆底部5 cm的土層處均勻擺放20粒馬唐種子,然后分別覆土0、0.5、1、2、3、4、5 cm和6 cm,將花盆放入淺盤中,采用底部滲灌的方式澆水,保持濕潤,置于溫度25~35℃、相對濕度70%~85%、自然光照的玻璃溫室中培養。每處理3次重復,每24 h統計1次出苗數,共統計21 d,胚芽鞘伸出土壤表層可見時視為出苗[18]。
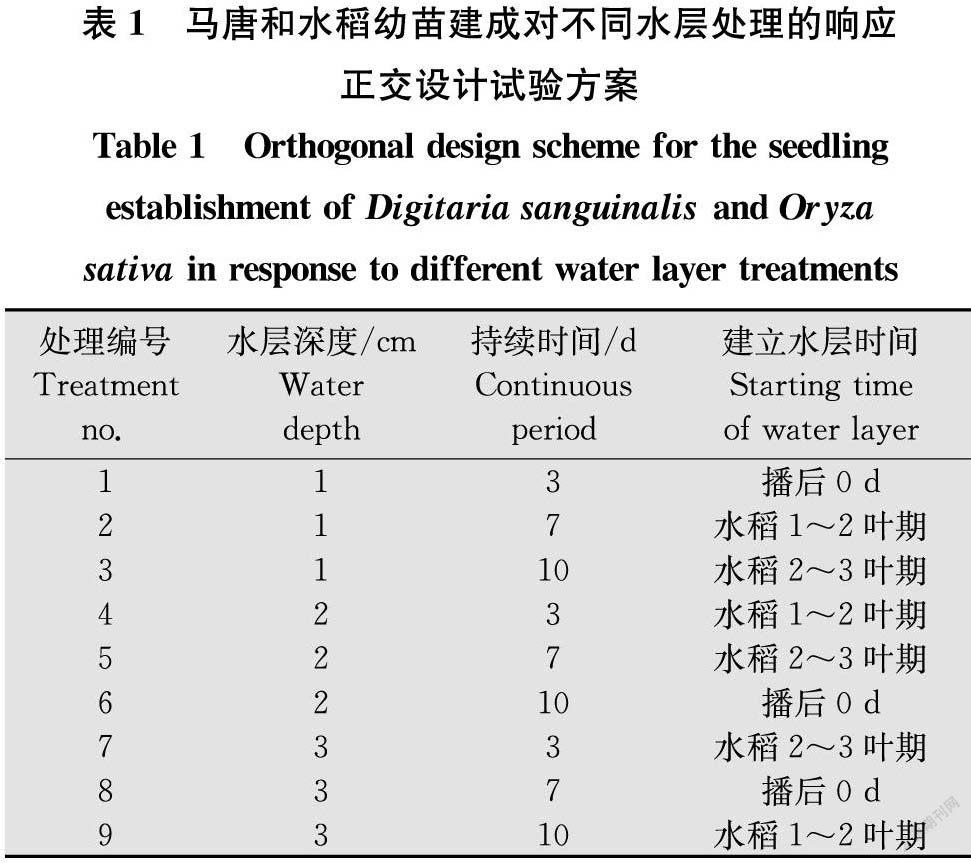
1.2.7 馬唐和水稻幼苗建成對水層的響應
在長方形盤(長、寬、高分別為22、15、7 cm)中裝入1.2.6中所述土壤,土層深度3 cm,每盆播種水稻和馬唐種子10粒。水稻品種為‘粵農絲苗’,來自廣東省農業科學院水稻研究所。采用正交設計法,設定3個因素(自變量),即水層深度、持續時間和建立水層時間,每個因素均為3個水平,具體設計見表1。非試驗處理周期內,土壤保持充分濕潤但不建立水層。另設一全程保持土壤充分濕潤但不建立水層處理作為對照。將材料放在1.2.6中的玻璃溫室中培養,環境條件同1.2.6,每天補充水分至目標水層深度。最后1次試驗處理結束后20 d檢查結果,以成苗率及地上部干重作為考察指標,每處理3次重復。
1.3 數據統計
采用SPSS 21.0軟件對同一試驗中不同處理的試驗結果進行單因素方差分析(One-way ANOVA),采用最小顯著差異法(LSD)比較不同處理間的差異顯著性。采用SigmaPlot 12.5軟件作圖,并對種子萌發率或出苗率與滲透勢、NaCl濃度和埋藏深度等的關系采用以下三參數非線性回歸方程進行曲線擬合[15, 19]:
式中,y表示某一時間或環境因子下的萌發率或出苗率,a代表最大萌發率或出苗率,x表示時間或NaCl濃度、滲透勢、埋藏深度等,x50表示達到最大萌發率或出苗率一半時對應的時間或NaCl濃度、滲透勢、埋藏深度等,b代表x50值附近的曲線斜率。
2 結果與分析
2.1 馬唐種子萌發對溫度的響應
不同溫度條件下,馬唐種子萌發率具有較大差異(圖1,表2)。在15℃/5℃變溫處理下,馬唐種子無法萌發;25℃/15℃、30℃/20℃和35℃/25℃條件適宜馬唐種子萌發,萌發率分別為88.3%、91.7%和88.3%;與上述溫度條件相比,光照階段溫度達到40℃,或黑暗階段溫度降低到10℃馬唐種子萌發均受到抑制。馬唐在20℃/10℃和40℃/30℃溫度條件下萌發率分別為55.0%和70.0%。除最終萌發率外,不同溫度條件下馬唐種子起始萌發時間和達到最終萌發率一半時所需時間(x50)也存在差異(表2),在20℃/10℃至40℃/30℃范圍內,馬唐起始萌發時間和x50值逐漸減小,在40℃/30℃條件下第2天即可萌發,有助于其在高溫環境中快速搶占生態位。
2.2 馬唐種子萌發對光照的響應
馬唐種子在無光照條件下仍可萌發,但萌發率顯著低于光照24 h/d和光照12 h/d處理組,光照24 h/d和光照12 h/d處理組之間馬唐種子萌發率無顯著差異(表3)。因此,光照對馬唐種子萌發具有一定的刺激作用,但并非其萌發的必要條件,遮蔭處理可能有助于減少馬唐在田間的萌發出苗數。
2.3 馬唐種子萌發對pH的響應
馬唐種子在pH 4~10的溶液中均可萌發(圖2)。在pH為7~10時,馬唐種子萌發率在93.3%~96.7%,不同處理間差異不顯著(P>0.05);在pH為4~6時,馬唐種子萌發率顯著低于其他處理(P<0.05),但萌發率仍在63.3%~85.0%。上述結果表明,馬唐種子萌發對pH適應范圍廣,可以在多數農田土壤環境中萌發。
2.4 馬唐種子萌發對滲透勢的響應
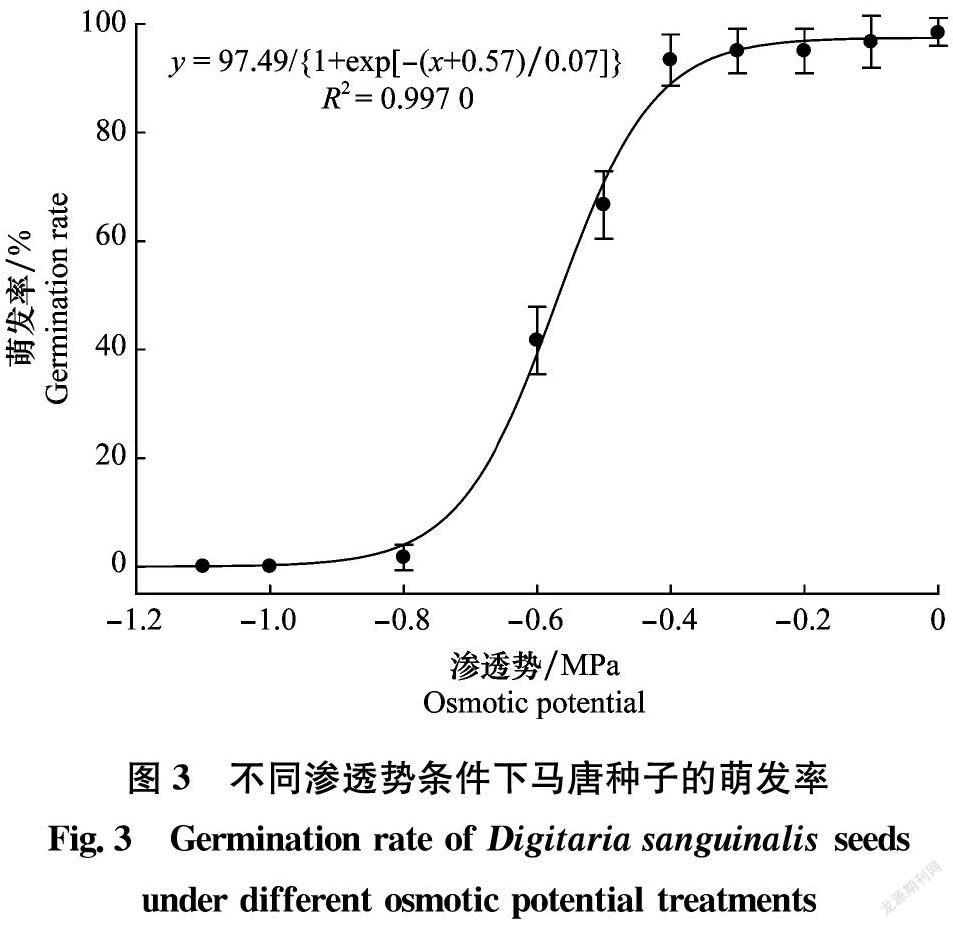
馬唐種子萌發率與滲透勢的關系符合三參數非線性回歸方程y=97.49/{1+exp[-(x+0.57)/0.07]}(R2=0.997 0)(圖3)。滲透勢為-0.4~0 MPa時,馬唐種子萌發率達93.3%~98.3%;滲透勢為-0.5 MPa和-0.6 MPa時,馬唐種子萌發率迅速下降至66.7%和41.7%;當滲透勢為-0.8 MPa及以下時,馬唐種子基本無法萌發,萌發率低于1.7%(圖3)。上述結果表明,馬唐對滲透勢的適應范圍較廣,在多種土壤濕度條件下均可能萌發。
2.5 馬唐種子萌發對鹽分脅迫的響應
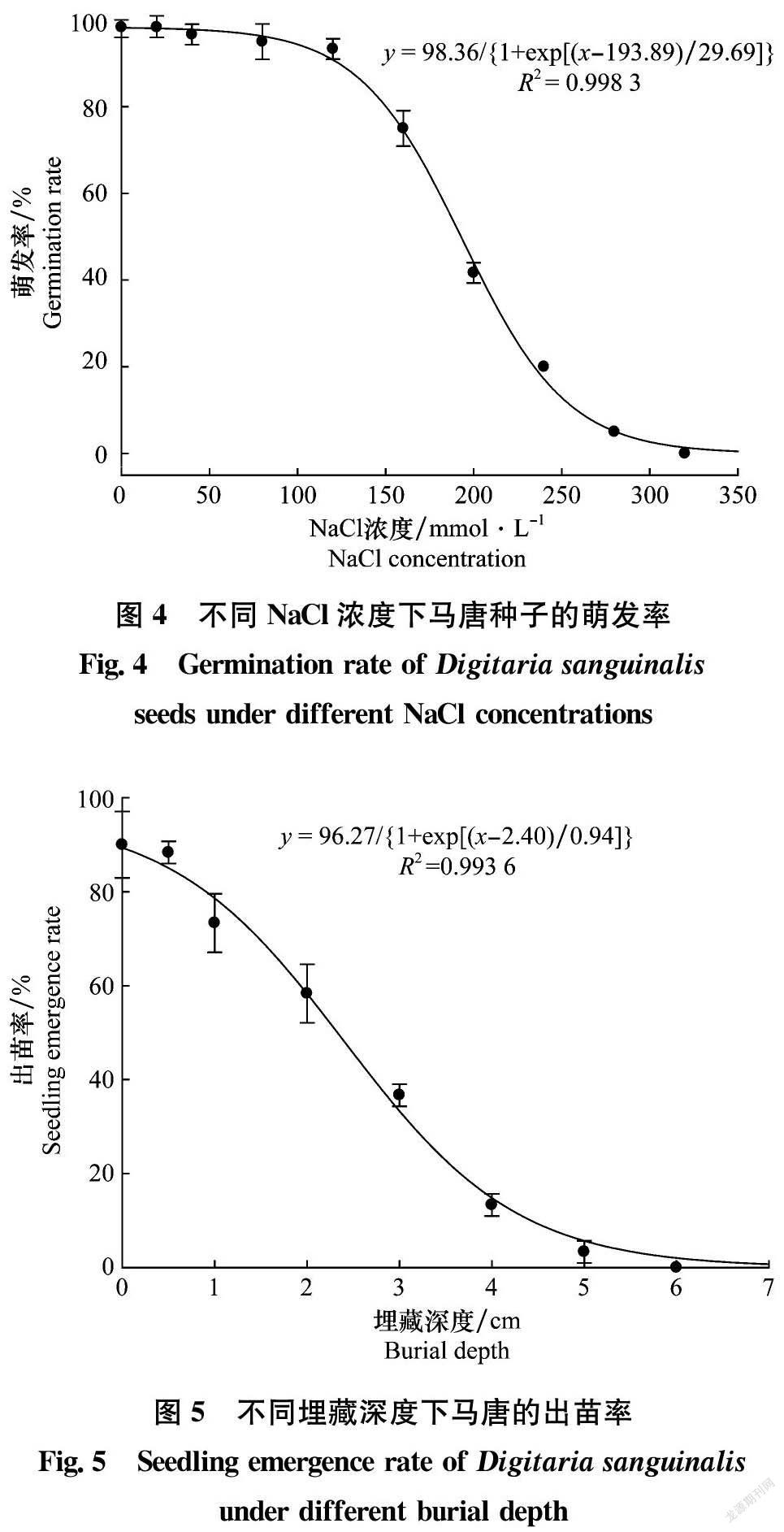
馬唐種子萌發率與NaCl濃度的關系符合三參數非線性回歸方程y=98.36/{1+exp[(x-193.89)/29.69]}(R2=0.998 3)(圖4)。當NaCl濃度為0 mmol/L和20 mmol/L時,馬唐種子萌發率均為98.3%;隨著NaCl濃度增大,馬唐種子萌發率逐漸降低,當NaCl濃度為120 mmol/L時萌發率高達93.3%。根據上述方程計算,抑制最大萌發率50%時的NaCl濃度為193 mmol/L。值得注意的是, NaCl濃度為240 mmol/L時仍有20%的馬唐種子萌發。上述結果表明,馬唐對鹽分脅迫具有較強的耐受能力,可適應大多數農田土壤環境條件。
2.6 馬唐出苗對埋藏深度的響應
馬唐出苗率與埋藏深度的關系符合三參數非線性回歸方程y=96.27/{1+exp[(x-2.40)/0.94]}(R2=0.993 6)(圖5)。當埋藏深度為0 cm時,馬唐出苗率最高,達90.0%,埋藏深度0.5 cm時出苗率為88.3%,與0 cm時無顯著性差異(P>0.05)。隨著埋藏深度進一步增加,馬唐出苗率迅速下降,埋藏深度5 cm時出苗率僅為3.3%,埋藏深度為6 cm時馬唐無法完成出苗。
2.7 馬唐和水稻幼苗建成對水層的響應
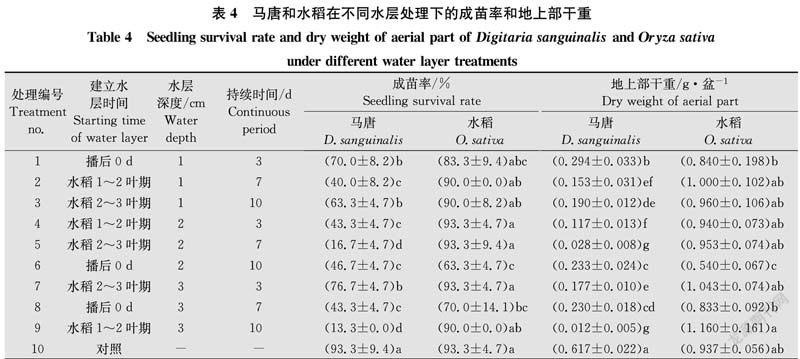
水層深度、水層持續周期及建立水層時間對馬唐和水稻的成苗率、地上部干重均具有一定程度的影響。從成苗率看,播后0 d建立2 cm水層持續10 d與播后0 d建立3 cm水層持續7 d水稻成苗率分別為63.3%和70.0%,顯著低于對照(93.3%)及其他水層處理(83.3%~93.3%)(P<0.05);水稻2~3葉期建立2 cm水層持續7 d和水稻1~2葉期建立3 cm水層持續10 d馬唐成苗率分別為16.7%和13.3%,與其他處理差異顯著(P<0.05),其他水層處理馬唐成苗率在40.0%~76.7%,對照組馬唐成苗率高達93.3%(表4)。從地上部干重看,播后0 d建立1 cm水層持續3 d,建立2 cm水層持續10 d和建立3 cm水層持續7 d水稻地上部干重分別為0.840、0.540 g和0.833 g,顯著低于水稻1~2葉期建立3 cm水層持續10 d(P<0.05),其他處理與水稻1~2葉期建立3 cm水層持續10 d水稻地上部干重差異不顯著(P>0.05)(表4);對照組馬唐地上部干重為0.617 g,水層處理馬唐地上部干重均顯著低于對照(P<0.05),其中水稻2~3葉期建立2 cm水層持續7 d和水稻1~2葉期建立3 cm水層持續10 d地上部干重分別為0.028 g和0.012 g,顯著低于其他處理(P<0.05),與對照相比干重抑制率達95%以上,其他水層處理馬唐地上部干重在0.117~0.294 g,與對照處理相比干重抑制率在52%~81%。綜合分析可知,水稻2~3葉期建立2 cm水層持續7 d和水稻1~2葉期建立3 cm水層持續10 d可以在不影響水稻正常生長的前提下較好地控制馬唐的發生危害。
3 討論
農田生態系統是受人類活動影響較大的一種生態系統,雜草在農田生態系統中不僅受光照、溫度、降水等自然環境因子影響,還受到農作物栽培方式、耕作方式、施肥、灌溉等農事操作的影響[20]。種子萌發和幼苗建成是雜草在田間存活并造成危害的關鍵環節,也是進行雜草防控的最佳窗口期,雜草對不同環境因子的響應情況反映了其對不同環境的適應性和競爭性,據此可針對性地采取控制措施,減輕雜草在田間的發生危害[1112, 21]。
溫度是影響種子萌發的重要環境因子,本研究表明,馬唐在20℃/10℃~40℃/30℃的變溫條件下均可萌發,說明馬唐具有較寬的溫度適應范圍。溫廣月等[22]研究表明,馬唐在15~40℃時均可萌發,在10℃和45℃時無法萌發。與馬唐生育期較一致的其他禾本科雜草也表現出相似的結果,如在25℃/15℃、30℃/20℃及35℃/25℃變溫條件下,升馬唐Digitaria ciliaris萌發率為39%~93%,長花馬唐Digitaria longiflora萌發率為78%~89%[4],牛筋草Eleusine indica萌發率為10%~72%[19],與本研究的結果相一致,而千金子Leptochloa chinensis等雜草則只適合在較高溫度下萌發[23]。在春季至秋季,我國南北方大部分地區均有適合馬唐萌發的溫度條件,因此馬唐的分布范圍廣布全國各地,華南地區因溫度適宜,馬唐在一年內可繁殖多代,甚至周年發生[24],極大地提高了其在田間生存的幾率。
不同雜草種子萌發對光照的響應差異較大。有些雜草種子萌發對光照不敏感,光照或黑暗條件下萌發率無明顯差異,如看麥娘Alopecurus aequalis[25]、雀麥Bromus japonicus[26]、菵草Beckmannia syzigachne[27]等。有些雜草則屬于嚴格需光雜草,黑暗條件下幾乎不萌發,如長花馬唐[4]、勝紅薊Ageratum conyzoides[12]、鱧腸Eclipta prostrata[28]等。本研究發現馬唐在持續光照和光暗交替條件下萌發率均在90%以上,在黑暗條件下萌發率為66.7%,顯著低于光照條件下的萌發率。這說明光照不是馬唐種子萌發的必要因素,但對其萌發具有一定的刺激作用,升馬唐[4]、光頭稗[14]、牛筋草[19]、藜Chenopodium album[29]等雜草種子的萌發也具有相似的結果。不同深度的土層中光照強度差異較大,隨著土層深度增加,土層中的光照強度迅速下降,據報道,雜草種子在2 mm深的土層中接收到的光照強度僅為土壤表面的1%[12]。因此,對于嚴格需光種子,在較深的土層中無法萌發出苗,如東方大蒜芥Sisymbrium orientale種子在2.5 mm土層中萌發出苗率低于2%[30],勝紅薊在1 cm土層中無法萌發[12]。本研究表明,馬唐在土表及0.5 cm的淺土層中出苗率較高,隨著土層深度增加出苗率持續下降,在6 cm深的土層中無法出苗。這與光照處理的結果相一致:即馬唐不屬于嚴格需光的種子,但光照對其具有一定的刺激作用。除光照強度的影響外,隨著土層深度的增加,雜草種子本身的養分含量也可能不足以支持其萌發出苗,除少數雜草外,馬唐、升馬唐[4]、長花馬唐[4]、光頭稗[14]、棒頭草Polypogon fugax[18]、牛筋草[19]、千金子[23]、看麥娘[25]等常見禾本科雜草在6 cm深的土層中均無法出苗或出苗率極低。因此,在免耕或少耕條件下,大多數雜草種子聚集在土壤表層并暴露在陽光下,雜草更容易萌發出苗,而土壤翻耕處理有助于減少種子與光照接觸,增加其出苗的難度,從而減輕雜草發生危害[19,31]。
土壤酸堿度、滲透勢、鹽濃度通常會成為影響植物萌發、生長的環境因子。本研究表明,馬唐在pH 4~10的溶液中萌發率在63.3%~96.7%,尤其在pH 4.0的條件下萌發率仍高達63.3%,光頭稗[14]、棒頭草[18]、牛筋草[19]等雜草也具有相似的結果,而有些雜草對極端pH環境適應能力較差,如雀麥在pH 4的環境中無法萌發[26],看麥娘在pH 4的環境中萌發率僅為27%[25]。中國不同地區農田土壤pH多處于4.5~9.0[32],因此,馬唐對我國不同類型的土壤均具有較強的適應能力,有利于其在不同地區、不同生境侵入、定殖。一些雜草僅能適應較潮濕的土壤環境,在略干燥的土壤條件下萌發率明顯降低,如:勝紅薊在滲透勢-0.4 MPa時萌發率僅為30%,滲透勢-0.6 MPa時無法萌發[12];看麥娘、牛筋草等在滲透勢<-0.2 MPa時萌發率快速下降,滲透勢-0.6 MPa時無法萌發[19,25];棒頭草在滲透勢-0.4 MPa時萌發率低于15%,滲透勢-0.5 MPa時即可完全抑制萌發[18]。本研究發現,馬唐在滲透勢-0.4~0 MPa時,萌發率均在90%以上,滲透勢-0.6 MPa時萌發率仍達41.7%,說明馬唐對潮濕和干燥的土壤環境均具有較強的適應能力。因此,在直播稻田早期干濕交替的土壤環境中,馬唐相對于其他雜草可能具有一定的生長優勢,這可能是導致馬唐在直播稻田發生危害的重要因素。本研究表明,馬唐在240 mmol/L的NaCl溶液中萌發率達20%,在280 mmol/L的NaCl溶液中仍可萌發,而光頭稗[14]、牛筋草[19]、長花馬唐[4]等多種雜草在200 mmol/L的NaCl溶液中均無法萌發,錦葵屬植物Malva parviflora在80 mmol/L的NaCl溶液中萌發率僅為10%,在160 mmol/L濃度下無法萌發[33]。因此,相對于其他一些雜草,馬唐對高鹽環境可能具有更強的適應能力。
水層是控制稻田雜草的重要手段,在直播稻田中,水稻與雜草出苗時期一致,水層建立時間、水層深度和持續周期對水稻和雜草均有明顯影響。本研究表明,所有水層處理的馬唐成苗率及地上部干重均顯著低于對照處理,而多數水層處理的水稻成苗率和地上部干重與對照無顯著性差異,說明馬唐對水層的耐受能力低于水稻,這是利用水層控制馬唐在稻田危害的基礎。在播后0 d建立水層,與對照相比,水稻的成苗率和地上部干重均有所下降;水稻1~2葉期及以后建立1~3 cm水層對水稻生長無明顯影響。劉慶虎[34]研究表明播后0 d建立2 cm水層可導致水稻在20 d的出苗率下降30%以上,而播后6 d及以后建立水層對水稻出苗影響較小,與本研究結果相一致。此外,播后0 d建立水層對馬唐的防效并不理想,在取消水層處理后仍有43.3%~70.0%的馬唐出苗,說明馬唐種子在經水層處理后仍保持萌發活力。因此,為減輕水層對水稻的影響并提高對馬唐的防效,應避免在水稻播種后立即建立水層。隨著建立水層時間延遲,水稻和馬唐均已出苗,對水層耐受能力逐漸增強,水層深度成為影響雜草防效的重要因素:對比水稻2~3葉時建立1 cm水層持續10 d和建立2 cm水層持續7 d可發現,前者的馬唐成苗率(63.3%)顯著高于前者(16.7%),這可能與此時期多數馬唐心葉高度大于1 cm有關,只有水層淹沒雜草心葉時才具有良好的控草效果[34]。水層持續周期是影響控草效果的另一重要因素,對比水稻1~2葉期建立1 cm水層持續7 d和建立2 cm水層持續3 d可發現,后者的馬唐成苗率(43.3%)略高于前者(40.0%);進一步比較發現,水稻2~3葉期建立2 cm水層持續7 d的馬唐成苗率(16.7%)顯著低于1~2葉期建立2 cm水層持續3 d(43.3%)。本研究采用正交設計法明確了水層建立時間、水層深度和持續周期對水稻和馬唐成苗及生長的影響,在獲得代表性數據的同時,大幅減少了處理次數,若采用全面組合試驗,每因素3個水平的情況下需至少設置27組處理。本研究還發現,在相同條件下馬唐出苗比水稻早1~2 d,但水稻出苗后生長速度更快,出苗后2 d心葉高度可超過馬唐,可利用其心葉高度差建立合適深度的水層。
參考文獻
[1] LI Jian, LI Mei, GAO Xingxiang, et al. A novel amino acid substitution Trp574Arg in acetolactate synthase (ALS) confers broad resistance to ALS-inhibiting herbicides in crabgrass (Digitaria sanguinalis) [J]. Pest Management Science, 2017, 73(12): 25382543.
[2] ZHU Jinwen, WANG Jian, DITOMMASO A, et al. Weed research status, challenges, and opportunities in China [J/OL]. Crop Protection, 2020, 134: 104449. DOI: 10.1016/j.cropro.2018.02.001.
[3] 蔣易凡, 陳國奇, 董立堯. 稻田馬唐對稻田常用莖葉處理除草劑的抗性水平研究[J]. 雜草學報, 2017, 35(2): 6772.
[4] CHAUHAN B S, JOHNSON D E. Germination ecology of southern crabgrass (Digitaria ciliaris) and India crabgrass (Digitaria longiflora): two important weeds of rice in tropics [J]. Weed Science, 2008, 56: 722728.
[5] 程來品, 曹方元, 仇學平, 等. 不同除草劑對直播稻田馬唐等雜草的防效[J]. 雜草科學, 2013, 31(1): 6465.
[6] 劉慶虎, 陳國奇, 張玉華, 等. 不同葉齡千金子、稗和馬唐對氰氟草酯和五氟磺草胺的敏感性[J]. 南京農業大學學報, 2016, 39(5): 771776.
[7] 王紅春, 徐蓬, 孫鈺晨, 等. 江蘇省稻田雜草的發生現狀與防控建議[J]. 雜草學報, 2019, 37(4): 15.
[8] 郭文磊, 吳丹丹, 張純, 等. 5種莖葉處理除草劑對直播稻田禾本科雜草的除草活性及田間防效[J]. 南方農業學報, 2019, 50(6): 12401246.
[9] HEAP I. International survey of herbicide resistant weeds [EB/OL]. (20191030) [20210105]. http: ∥www.weedscience.com.
[10]KAUNDUN S S. Resistance to acetyl-CoA carboxylase-inhibiting herbicides [J]. Pest Management Science, 2014, 70: 14051417.
[11]RAJJOU L, DUVAL M, GALLARDO K, et al. Seed germination and vigor [J]. Annual Review of Plant Biology, 2012, 63(1): 507533.
[12]杜麗思, 李銣, 董玉梅, 等. 勝紅薊種子萌發/出苗對環境因子的響應[J]. 生態學報, 2019, 39(15): 56625669.
[13]BEWLEY J D, BRADFORD K, HILHORST H, et al. Seeds: physiology of development, germination and dormancy [M]. New York: Springer-Verlag, 2012.
[14]CHAUHAN B S, JOHNSON D E. Seed germination ecology of junglerice (Echinochloa colona): a major weed of rice [J]. Weed Science, 2009, 57(3): 235240.
[15]VILA-AIUB M M, NEVE P, STEADMAN K J, et al. Ecological fitness of a multiple herbicide-resistant Lolium rigidum population: dynamics of seed germination and seedling emergence of resistant and susceptible phenotypes [J]. Journal of Applied Ecology, 2005, 42(2): 288298.
[16]CHACHALIS D, REDDY K N. Factors affecting Campsis radicans seed germination and seedling emergence [J]. Weed Science, 2000, 48(2): 212216.
[17]MICHEL B E, RADCLIFFE D. A computer program relating solute potential to solution composition for five solutes [J]. Agronomy Journal, 1995, 87(1): 126130.
[18]WU Xian, LI Jun, XU Hongle, et al. Factors affecting seed germination and seedling emergence of Asia minor bluegrass (Polypogon fugax) [J]. Weed Science, 2015, 63(2): 440447.
[19]CHAUHAN B S, JOHNSON D E. Germination ecology of goosegrass (Eluesine indica): an important grass weed of rainfed rice [J]. Weed Science, 2008, 56(5): 699706.
[20]趙玉信, 楊惠敏. 作物格局、土壤耕作和水肥管理對農田雜草發生的影響及其調控機制[J]. 草業學報, 2015, 24(8): 199210.
[21]李香菊. 近年我國農田雜草防控中的突出問題與治理對策[J]. 植物保護, 2018, 44(5): 7784.
[22]溫廣月, 錢振官, 李濤, 等. 馬唐生物學特性初步研究[J]. 雜草科學, 2014, 32(2): 14.
[23]BENVENUTI, S, DINELLI G, BONETTI A. Germination ecology of Leptochloa chinensis: a new weed in the Italian agro-environment [J]. Weed Research, 2004, 44(2): 8796.
[24]張純, 郭文磊, 張泰劼, 等. 廣東農田雜草防控的問題與對策[J]. 廣東農業科學, 2020, 47(12): 98108.
[25]ZHAO Ning, LI Qi, GUO Wenlei, et al. Effect of environmental factors on germination and emergence of shortawn foxtail (Alopecurus aequalis) [J]. Weed Science, 2018, 66(1): 4756.
[26]LI Qi, TAN Jinni, LI Wei, et al. Effects of environmental factors on seed germination and emergence of Japanese brome (Bromus japonicus) [J]. Weed Science, 2015, 63(1): 110.
[27]RAO Na, DONG Liyao, LI Jun, et al. Influence of environmental factors on seed germination and seedling emergence of American sloughgrass (Beckmannia syzigachne) [J]. Weed Science, 2008, 56(4): 529533.
[28]CHAUHAN B S, JOHNSON D E. Influence of environmental factors on seed germination and seedling emergence of eclipta (Eclipta prostrata) in a tropical environment [J]. Weed Science, 2008, 56(3): 383388.
[29]JURSIK M, SOUKUP J, HOLEC J. Seed dormancy and germination of shaggy soldier (Galinsoga ciliata Blake.) and common lambsquarter (Chenopodium album L.) [J]. Plant Soil and Environment, 2003, 49(11): 511518.
[30]CHAUHAN, B S, GILL G, PRESTON C. Influence of environmental factors on seed germination and seedling emergence of oriental mustard (Sisymbrium orientale) [J]. Weed Science, 2006, 54(5): 10251031.
[31]CHAUHAN, B S, GILL G, PRESTON C. Influence of tillage systems on vertical distribution, seedling recruitment and persistence of rigid ryegrass (Lolium rigidum) seed bank [J]. Weed Science, 2006, 54(4): 669676.
[32]戴萬宏, 黃耀, 武麗, 等. 中國地帶性土壤有機質含量與酸堿度的關系[J]. 土壤學報, 2009, 46(5): 851860.
[33]CHAUHAN B S, GILL G, PRESTON C. Factors affecting seed germination of little mallow (Malva parviflora) in southern Australia [J]. Weed Science, 2006, 54(6): 10451050.
[34]劉慶虎. 長江中下游地區直播稻田雜草種子庫及千金子(Leptochloa chinensis)防控技術研究[D]. 南京: 南京農業大學, 2016: 4350.
(責任編輯:楊明麗)
